Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Dimitrios Tsiptsios | -- | 1720 | 2023-02-24 07:34:34 | | | |
| 2 | Dean Liu | -3 word(s) | 1717 | 2023-02-27 02:40:34 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Papadopoulos, C.; Anagnostopoulos, K.; Tsiptsios, D.; Karatzetzou, S.; Liaptsi, E.; Lazaridou, I.Z.; Kokkotis, C.; Makri, E.; Ioannidou, M.; Aggelousis, N.; et al. Red Blood Cells in Stroke. Encyclopedia. Available online: https://encyclopedia.pub/entry/41612 (accessed on 13 July 2026).
Papadopoulos C, Anagnostopoulos K, Tsiptsios D, Karatzetzou S, Liaptsi E, Lazaridou IZ, et al. Red Blood Cells in Stroke. Encyclopedia. Available at: https://encyclopedia.pub/entry/41612. Accessed July 13, 2026.
Papadopoulos, Charalampos, Konstantinos Anagnostopoulos, Dimitrios Tsiptsios, Stella Karatzetzou, Eirini Liaptsi, Irene Zacharo Lazaridou, Christos Kokkotis, Evangelia Makri, Maria Ioannidou, Nikolaos Aggelousis, et al. "Red Blood Cells in Stroke" Encyclopedia, https://encyclopedia.pub/entry/41612 (accessed July 13, 2026).
Papadopoulos, C., Anagnostopoulos, K., Tsiptsios, D., Karatzetzou, S., Liaptsi, E., Lazaridou, I.Z., Kokkotis, C., Makri, E., Ioannidou, M., Aggelousis, N., & Vadikolias, K. (2023, February 24). Red Blood Cells in Stroke. In Encyclopedia. https://encyclopedia.pub/entry/41612
Papadopoulos, Charalampos, et al. "Red Blood Cells in Stroke." Encyclopedia. Web. 24 February, 2023.
Copy Citation
Stroke constitutes the second highest cause of morbidity and mortality worldwide while also impacting the world economy, triggering substantial financial burden in national health systems. High levels of blood glucose, homocysteine, and cholesterol are causative factors for atherothrombosis. These molecules induce erythrocyte dysfunction, which can culminate in atherosclerosis, thrombosis, thrombus stabilization, and post-stroke hypoxia. Glucose, toxic lipids, and homocysteine result in erythrocyte oxidative stress. This leads to phosphatidylserine exposure, promoting phagocytosis. Phagocytosis by endothelial cells, intraplaque macrophages, and vascular smooth muscle cells contribute to the expansion of the atherosclerotic plaque.
erythrocyte
ischemic stroke
atherothrombosis
lipotoxicity
1. Red Blood Cells Participate in Hypoxia after Atherothrombotic Stroke
Erythrocytes regulate oxygen release. This function is mainly regulated by the formation of the glycolytic intermediate 2,3 biphosphoglycerate (2,3-BPG). This metabolite is formed by biphosphoglycerate mutase and results in the lower oxygen affinity of hemoglobin. The activity of mutase in erythrocytes increases after exposure to hypoxia. Its levels of activity are regulated by transglutaminase, which stabilizes mutase [1]. In addition, the formation of 2,3-BPG is favored when glycolysis is upregulated. In red blood cells under hypoxia, this is mediated by adenosine receptor 2b-mediated sphingosine kinase 1 activity, and the subsequent sphingosine-1-phosphate (S1P)-induced glycolytic enzyme is release from the plasma membrane to the cytoplasm [2]. In addition, erythrocyte adenosine receptor 2b activation during hypoxia also results in adenosine monophosphate-dependent kinase (AMPK) activation and AMPK-dependent BPG mutase [3]. Furthermore, S1P increase also enhances AMPK activation and subsequent BPG mutase elevated activity by downregulating the activity of the pp2a phosphatase, which negatively controls AMPK [4]. Increased levels of erythrocyte 2,3-BPG have been shown to protect from ischemia [5], while the beneficial effects of remote ischemic conditioning are attributed to increased erythrocyte 2,3-BPG [5].
However, obesity and aging have been associated with reduced 2,3-BPG levels [6] and BPG mutase activity [2], respectively (Figure 1). Animals lacking erythrocyte adenosine receptor a2b, and consequently, BPG mutase activity, present markers of brain inflammation.
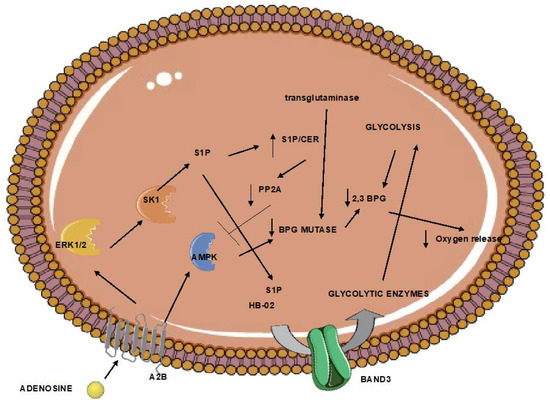
Figure 1. Erythrocytes trapped in the ischemic tissue can respond to limited oxygen through the regulation of the 2,3-bisphosphoglycerate. However, aging and obesity halter this pathway. The end result can be increased brain inflammation. (A normal arrow signifies activation. An arrow with a vertical line signifies inhibition). 2,3 BPG: 2,3 biphosphoglycerate; A2B: adenosine receptor 2B; AMPK; adenosine monophosphate-activated kinase, BPG mutase: biphosphoglycerate mutase; CER: ceramide; ERK1/2: extracellular signal-regulated kinase; HB-O2: oxygenated hemoglobin; S1P: sphingosine-1 phosphate; SK1: sphingosine kinase 1.
2. Red blood Cells Respond to Damage-Associated Molecular Patterns Released after Ischemic Stroke
Ischemia and sustained oxygen deprivation of the brain can lead to the release of damage-associated molecular patterns (DAMPs), such as ATP, extracellular histones, amyloids, and cell-free mitochondrial DNA. DAMPs can act on microglial cells and infiltrate immune cells. This event regulates both brain inflammation and tissue repair [7]. Apart from DAMPs, pathogen-associated molecular patterns (PAMPs) are also implicated in the progression of stroke; lipopolysaccharide (LPS), a component of Gram-negative bacteria, is associated with poor prognosis in stroke [8]. Apart from immune cells, DAMPs can also act on red blood cells (Table 1). Erythrocytes express various protein receptors that are capable not only of scavenging but also sensing DAMPs and PAMPs. In addition, as analyzed below, several DAMPs and PAMPs trigger physicochemical alterations on red blood cells. The net effect of DAMP-induced erythrocyte dysfunction is mainly determined by the effect of erythrophagocytosis on macrophage polarization. While in the liver erythrolysis prior to erythrophagocytosis results in an anti-inflammatory phenotype of macrophages [9], in the brain, this mechanism induces inflammation [10]. Researchers speculate that the relative quantities of CD47, oxidized CD47, exposed phosphatidylserine, and the level of ROS determine the ratio of erythrolysis: intact erythrophagocytosis.
Table 1. Summary of the mechanisms underlying the response of red blood cells to pathogen- and damage-associated molecular patterns.
| Damp/Pamp | Receptor | Effect |
|---|---|---|
| mtDNA | TLR9 |
|
| CpG DNA | TLR9 |
|
| ATP | P2X7 |
|
| Extracellular histones | TLR2? |
|
| LPS | - |
|
| Amyloids | - |
|
ATP: adenosine triphosphate; CpG DNA: cytosine-guanine deoxyribonucleic acid; LPS: lipopolysaccharide; mtDNA: mitochondrial deoxyribonucleic acid; P2X7: purinergic receptor 2x7; TLR2: toll-like receptor 2; TLR9: toll-like receptor 9.
2.1. Cell-Free Mitochondrial DNA and CpG DNA
Hotz et al. [11] were the first to report that human erythrocytes expressed the toll-like receptor 9 (TLR9) on their plasma membranes. In their study, they showed that TLR9 on erythrocyte membranes bound cell-free mitochondrial DNA (mtDNA). In fact, they reported that when the circulating levels of mtDNA were low, most TLR9s of erythrocytes had bound mtDNA. However, an increase in the circulating levels of mtDNA resulted in the saturation of the scavenging capacity of red blood cells. In addition, the loss of TLR9 on red blood cells led to increased lung injury, while the administration of red blood cells from healthy donors attenuated the CpG DNA-induced inflammation.
Subsequent reports from the same research group provided important clues for the effects of cell-free mitochondrial DNA and CpG DNA, in general, on erythrocytes. First, red blood cells from septic patients contained increased TLR9 and bound CpG DNA levels in comparison to healthy controls [12]. Furthermore, CpG DNA was found to induce morphological changes on erythrocytes in a TLR9-dependent manner. Specifically, a redistribution of BAND3 was observed, as well as a conformational change and the loss of the “do not eat me” signaling protein CD47. It was also remarkable that CpG DNA binding to TLR9 brought about a marked increase in the levels of TLR9 on the surface of erythrocytes. Finally, it was shown that CpG DNA binding to TLR9 augmented erythrophagocytosis and inflammation in vivo.
These results provide clear evidence that red blood cells respond to CpG DNA, both as a DAMP and a PAMP.
2.2. ATP
Parker and Snow [13] first showed that canine red blood cells expressed the P2X7 receptor, and its activation by ATP led to an increased cation flux. This effect was not seen when erythrocytes were incubated with other adenine nucleotides. It is notable that the P2X7 receptor is also expressed on the membranes of human erythrocytes [14], and its activation by ATP increases cation flux. More importantly, the activation of the P2X7 receptor of human red blood cells leads to phosphatidylserine exposure [15]. However, this effect was much more prominent in canine erythrocytes, perhaps due to the higher levels of P2X7 receptors [15] expressed therein. The ATP-induced phosphatidylserine exposure of canine erythrocytes was also confirmed by Faulks et al. [16], excluding other nucleotides as triggers for this effect. Another important finding came from the study of Sophocleous et al. [17], who reported that the P2X7 activation-induced phosphatidylserine exposure of canine red blood cells was not altered during cellular aging. This may indicate that P2X7 is not expelled from the membrane through vesiculation.
2.3. Extracellular Histones
In 2014, Semerano et al. [18] reported that extracellular histones induced phosphatidylserine exposure on human erythrocytes. This effect was mainly mediated by histone 4 (H4) and led to the increased activation of thrombin. Later, another research group found that extracellular histones triggered anemia and increased erythrocyte fragility and tendency to aggregate [19]. The results of Semerano et al. were confirmed by Yeung et al. [20], who reported histone-induced phosphatidylserine exposure on red blood cells. They found that extracellular histones induced calcium influx, generation of reactive oxygen species, and activation of caspase-3. Remarkably, these effects were attenuated with the pre-treatment of red blood cells with a neutralizing antibody for TLR2. Other investigators also found that all histones could bind to erythrocytes, albeit to different extents [21]. All the extracellular histones induced hemoglobin release and generation of microvesicles with externalized phosphatidylserine. These results could be mediated by the opening or formation of ion pores by histones [21].
2.4. Lipopolysaccharides (LPS)
Lipopolysaccharides can induce dramatic changes in the conformation of the erythrocyte membrane proteins. This effect is mainly attributed to the hydrophobic interactions of LPS with lipids and proteins, which can subsequently influence the protein–lipid interaction, thus disrupting the red blood cell cytoskeleton and membrane stability [22]. These results were recapitulated in later studies, where LPS was found to increase erythrocyte osmotic fragility and decrease lipid fluidity [23]. More recently, Brauckmann et al. [24] reported that LPS could provoke hemolysis with direct interaction with washed red blood cells. Hence, these studies support the idea of LPS-induced physicochemical changes on erythrocytes, which could result in hemolysis.
2.5. Amyloids
First, Nicolay et al. [25] showed that the incubation of human red blood cells with amyloid beta (1–42) induced sphingomyelin hydrolysis and phosphatidylserine exposure. This effect was amplified by the depletion of intracellular Cl−. Other researchers reported that amyloid Aβ (25–35) downregulated the activity of several glycolytic enzymes and upregulated the Na+/K+ ATPase activity of rat erythrocytes in a cellular age-dependent manner [26]. The latter results possibly hold for human red blood cells too. Aβ amyloid can uncouple the erythrocyte metabolism from oxygen saturation [27], perhaps through the trimeric G protein activation-induced activation of adenylic cyclase and caspase 3 [28]. This pathway also results in the inhibition of ATP release from red blood cells [28]. Further studies identified protein kinase C and nitric oxide as important constituents of this pathway [29][30]. Apart from the regulation of metabolism, this pathway also influences erythrocyte morphology [31].
3. Red Blood Cells Could Connect Non-Alcoholic Fatty Liver Disease with the Risk of Atherothrombotic Strokes
NAFLD severity is associated with an increased risk of stroke [31]. Interestingly, the red blood cells of NAFLD patients present characteristics that are involved in the mechanisms of atherothrombosis. The group has shown that erythrocytes obtained from NAFLD patients exhibited increased membrane cholesterol, sphingosine, bound chemokine MCP1 [32], reduced levels of sphingomyelin [33], and CD47 [32], while also increasing the release of MCP1 [34] and the sustained release of sphingosine-1 phosphate and lysophosphatidic acid [34]. Previously, other researchers showed that erythrocytes of NAFLD patients and animal models also exhibited increased phosphatidylserine exposure and reactive oxygen species [35]. Thus, researchers speculate that erythrocytes of NAFLD patients could partially expose patients to a greater risk for the development of atherothrombotic stroke.
References
- Xu, P.; Chen, C.; Zhang, Y.; Dzieciatkowska, M.; Brown, B.C.; Zhang, W.; Xie, T.; Abdulmalik, O.; Song, A.; Tong, C.; et al. Erythrocyte Transglutaminase-2 Combats Hypoxia and Chronic Kidney Disease by Promoting Oxygen Delivery and Carnitine Homeostasis. Cell Metab. 2022, 34, 299–316.e6.
- Qiang, Q.; Manalo, J.M.; Sun, H.; Zhang, Y.; Song, A.; Wen, A.Q.; Edward Wen, Y.; Chen, C.; Liu, H.; Cui, Y.; et al. Erythrocyte Adenosine A2B Receptor Prevents Cognitive and Auditory Dysfunction by Promoting Hypoxic and Metabolic Reprogramming. PLoS Biol. 2021, 19, e3001239.
- Sun, K.; Zhang, Y.; Bogdanov, M.v.; Wu, H.; Song, A.; Li, J.; Dowhan, W.; Idowu, M.; Juneja, H.S.; Molina, J.G.; et al. Elevated Adenosine Signaling via Adenosine A2B Receptor Induces Normal and Sickle Erythrocyte Sphingosine Kinase 1 Activity. Blood 2015, 125, 1643–1652.
- Sun, K.; Zhang, Y.; D’Alessandro, A.; Nemkov, T.; Song, A.; Wu, H.; Liu, H.; Adebiyi, M.; Huang, A.; Wen, Y.E.; et al. Sphingosine-1-Phosphate Promotes Erythrocyte Glycolysis and Oxygen Release for Adaptation to High-Altitude Hypoxia. Nat. Commun. 2016, 7, 12086.
- Kimura, H.; Hamasaki, N.; Yamamoto, M.; Tomonaga, M. Circulation of Red Blood Cells Having High Levels of 2,3-Bisphosphoglycerate Protects Rat Brain from Ischemic Metabolic Changes during Hemodilution. Stroke 1995, 26, 1431–1437.
- Domingo-Ortí, I.; Lamas-Domingo, R.; Ciudin, A.; Hernández, C.; Herance, J.R.; Palomino-Schätzlein, M.; Pineda-Lucena, A. Metabolic Footprint of Aging and Obesity in Red Blood Cells. Aging 2021, 13, 4850.
- Stanzione, R.; Forte, M.; Cotugno, M.; Bianchi, F.; Marchitti, S.; Rubattu, S. Role of DAMPs and of Leukocytes Infiltration in Ischemic Stroke: Insights from Animal Models and Translation to the Human Disease. Cell. Mol. Neurobiol. 2022, 42, 545–556.
- Hakoupian, M.; Ferino, E.; Jickling, G.C.; Amini, H.; Stamova, B.; Ander, B.P.; Alomar, N.; Sharp, F.R.; Zhan, X. Bacterial lipopolysaccharide is associated with stroke. Sci. Rep. 2021, 11, 6570.
- Pfefferlé, M.; Ingoglia, G.; Schaer, C.A.; Yalamanoglu, A.; Buzzi, R.; Dubach, I.L.; Tan, G.; López-Cano, E.Y.; Schulthess, N.; Hansen, K.; et al. Hemolysis transforms liver macrophages into antiinflammatory erythrophagocytes. J. Clin. Investig. 2020, 130, 5576–5590.
- Ni, W.; Mao, S.; Xi, G.; Keep, R.F.; Hua, Y. Role of Erythrocyte CD47 in Intracerebral Hematoma Clearance. Stroke 2016, 47, 505–511.
- Hotz, M.J.; Qing, D.; Shashaty, M.G.S.; Zhang, P.; Faust, H.; Sondheimer, N.; Rivella, S.; Worthen, G.S.; Mangalmurti, N.S. Red Blood Cells Homeostatically Bind Mitochondrial DNA through TLR9 to Maintain Quiescence and to Prevent Lung Injury. Am. J. Respir. Crit. Care Med. 2018, 197, 470–480.
- Lam, L.K.M.; Murphy, S.; Kokkinaki, D.; Venosa, A.; Sherrill-Mix, S.; Casu, C.; Rivella, S.; Weiner, A.; Park, J.; Shin, S.; et al. DNA binding to TLR9 expressed by red blood cells promotes innate immune activation and anemia. Sci. Transl. Med. 2021, 13, eabj1008.
- Parker, J.C.; Snow, R.L. Influence of external ATP on permeability and metabolism of dog red blood cells. Am. J. Physiol. 1972, 223, 888–893.
- Sluyter, R.; Shemon, A.N.; Barden, J.; Wiley, J.S. Extracellular ATP increases cation fluxes in human erythrocytes by activation of the P2X7 receptor. J. Biol. Chem. 2004, 279, 44749–44755.
- Sluyter, R.; Shemon, A.N.; Hughes, W.E.; Stevenson, R.O.; Georgiou, J.G.; Eslick, G.D.; Taylor, R.M.; Wiley, J.S. Canine erythrocytes express the P2X7 receptor: Greatly increased function compared with human erythrocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, 2090–2098.
- Faulks, M.; Kuit, T.A.; Sophocleous, R.A.; Curtis, B.L.; Curtis, S.J.; Jurak, L.M.; Sluyter, R. P2X7 receptor activation causes phosphatidylserine exposure in canine erythrocytes. World J. Hematol. 2016, 5, 88–93.
- Sophocleous, R.A.; Mullany, P.R.F.; Winter, K.M.; Marks, D.C.; Sluyter, R. Propensity of red blood cells to undergo P2X7 receptor-mediated phosphatidylserine exposure does not alter during in vivo or ex vivo aging. Transfusion 2015, 55, 1946–1954.
- Semeraro, F.; Ammollo, C.T.; Esmon, N.L.; Esmon, C.T. Histones induce phosphatidylserine exposure and a procoagulant phenotype in human red blood cells. J. Thromb. Haemost. 2014, 12, 1697–1702.
- Kordbacheh, F.; O’Meara, C.H.; Coupland, L.A.; Lelliott, P.M.; Parish, C.R. Extracellular histones induce erythrocyte fragility and anemia. Blood 2017, 130, 2884–2888.
- Yeung, K.W.; Lau, P.M.; Tsang, H.L.; Ho, H.P.; Kwan, Y.W.; Kong, S.K. Extracellular histones induced eryptotic death in human erythrocytes. Cell. Physiol. Biochem. 2019, 53, 229–241.
- Noubouossie, D.; Sokol, J.; Piegore, M.G.; Ilich, A.; Henderson, M.W.; Mooberry, M.; Monroe, D.; Key, S.N. Histones Induce the Release of Extracellular Hemoglobin and Red Blood Cell-Derived Microvesicles with Procoagulant Activity. Blood 2018, 132 (Suppl. S1), 2514.
- Gwoździński, K.; Pienia̧zek, A.; Kaca, W. Lipopolysaccharide from Proteus mirabilis O29 induces changes in red blood cell membrane lipids and proteins. Int. J. Biochem. Cell Biol. 2003, 35, 333–338.
- Gwozdzinski, K.; Pieniazek, A.; Sudak, B.; Kaca, W. Alterations in human red blood cell membrane properties induced by the lipopolysaccharide from Proteus mirabilis S1959. Chem. Biol. Interact. 2003, 146, 73–80.
- Brauckmann, S.; Effenberger-Neidnicht, K.; De Groot, H.; Nagel, M.; Mayer, C.; Peters, J.; Hartmann, M. Lipopolysaccharide-induced hemolysis: Evidence for direct membrane interactions. Sci. Rep. 2016, 6, 35508.
- Nicolay, J.P.; Gatz, S.; Liebig, G.; Gulbins, E.; Lang, F. Amyloid induced suicidal erythrocyte death. Cell. Physiol. Biochem. 2007, 19, 175–184.
- Tikhonova, L.A.; Kaminskii, Y.G.; Kosenko, E.A. Effects of amyloid-β peptide Aβ25-35 on glycolytic and antioxidant enzymes in erythrocytes of different ages. Biol. Bull. 2014, 41, 312–317.
- Clementi, M.E.; Giardina, B.; Colucci, D.; Galtieri, A.; Misiti, F. Amyloid-beta peptide affects the oxygen dependence of erythrocyte metabolism: A role for caspase 3. Int. J. Biochem. Cell Biol. 2007, 39, 727–735.
- Misiti, F.; Orsini, F.; Clementi, M.E.; Masala, D.; Tellone, E.; Galtieri, A.; Giardina, B. Amyloid peptide inhibits ATP release from human erythrocytes. Biochem. Cell Biol. 2008, 86, 501–508.
- Carelli-Alinovi, C.; Giardina, B.; Misiti, F. Amyloid beta peptide (1-42)-mediated antioxidant imbalance is associated with activation of protein kinase C in red blood cells. Cell Biochem. Funct. 2015, 33, 196–201.
- Carelli-Alinovi, C.; Dinarelli, S.; Sampaolese, B.; Misiti, F.; Girasole, M. Morphological changes induced in erythrocyte by amyloid beta peptide and glucose depletion: A combined atomic force microscopy and biochemical study. Biochim. Biophys. Acta-Biomembr. 2019, 1861, 236–244.
- Xu, J.; Dai, L.; Zhang, Y.; Wang, A.; Li, H.; Wang, Y.; Meng, X.; Wu, S.; Wang, Y. Severity of Nonalcoholic Fatty Liver Disease and Risk of Future Ischemic Stroke Events. Stroke 2021, 52, 103–110.
- Papadopoulos, C.; Spourita, E.; Mimidis, K.; Kolios, G.; Tentes, L.; Anagnostopoulos, K. Nonalcoholic Fatty Liver Disease Patients Exhibit Reduced CD47 and Increased Sphingosine, Cholesterol, and Monocyte Chemoattractant Protein-1 Levels in the Erythrocyte Membranes. Metab. Syndr. Relat. Disord. 2022, 20, 377–383.
- Papadopoulos, C.; Mimidis, K.; Tentes, I.; Tente, T.; Anagnostopoulos, K. Validation and application of a protocol for the extraction and quantitative analysis of sphingomyelin in erythrocyte membranes of patients with non-alcoholic fatty liver disease. JPC-J. Planar Chromatogr. 2021, 34, 411–418.
- Papadopoulos, C.; Mimidis, K.; Papazoglou, D.; Kolios, G.; Tentes, I.; Anagnostopoulos, K. Red Blood Cell-Conditioned Media from Non-Alcoholic Fatty Liver Disease Patients Contain Increased MCP1 and Induce TNF-α Release. Rep. Biochem. Mol. Biol. 2022, 11, 54–62.
- Otogawa, K.; Kinoshita, K.; Fujii, H.; Sakabe, M.; Shiga, R.; Nakatani, K.; Ikeda, K.; Nakajima, Y.; Ikura, Y.; Ueda, M.; et al. Erythrophagocytosis by liver macrophages (Kupffer cells) promotes oxidative stress, inflammation, and fibrosis in a rabbit model of steatohepatitis: Implications for the pathogenesis of human nonalcoholic steatohepatitis. Am. J. Pathol. 2007, 170, 967–980.
More
Information
Subjects:
Clinical Neurology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Revisions:
2 times
(View History)
Update Date:
27 Feb 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No