 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Emilio Cervantes | + 2104 word(s) | 2104 | 2021-08-24 08:29:37 | | | |
| 2 | Camila Xu | -71 word(s) | 2033 | 2021-09-13 04:06:11 | | | | |
| 3 | Camila Xu | -71 word(s) | 2033 | 2021-09-13 04:07:11 | | |
Video Upload Options
The Vitaceae Juss., in the basal lineages of Rosids, contains sixteen genera and 950 species, mainly of tropical lianas. The family has been divided in five tribes: Ampelopsideae, Cisseae, Cayratieae, Parthenocisseae and Viteae. Seed shape is variable in this family and is described based on the comparison of bi-dimensional seed images with geometric models. Ten morphological types are described in the Vitaceae. Seven of them are general and three specific. Among the general types, three are shared with the Arecaceae and correspond to geometric figures well described (lenses, superellipses and elongated waterdrops). Four additional groups include waterdrops, normal or rounded, heart curves, normal or rounded, elongated heart curves and other elongated curves, respectively. Finally, the three specific types correspond to heart curves of the Cayratia and Pseudocayratia types, heart curves of the Squared Heart Curve (SqHC) type of Ampelocissus and Ampelopsis, and Elongated Superellipse-Heart Curves (ESHCs), frequent in Tetrastigma species and observed also in Cissus species and R. rhomboidea. All these groups are defined by geometric models obtained by the representation of algebraic equations. Modifications in the equations result in models adjusting to the shape of seeds for each species.
1. Introduction
| Tribe | Genera |
| I. Ampelopsideae (47) | Ampelopsis Michx. (18)Nekemias Raf. (9) |
| Rhoicissus Planch. (14) | |
| Clematicissus Planch. (6) | |
| II. Cisseae (300) | Cissus L. (300) |
| III. Cayratieae (368) | Cayratia Juss. (25) |
| Causonis Raf. (30) | |
| Acareosperma Gagnep. (1) | |
| Afrocayratia (7) | |
| Cyphostemma (Planch.) Alston (200) | |
| Pseudocayratia J. Wen,L.M.Lu and Z.D. Chen (5) | |
| Tetrastigma (Miq.) Planch. (100) | |
| IV. Parthenocisseae (16) | Parthenocissus Planch. (14) |
| Yua C.L.Li (2) | |
| V. Viteae (190) | Ampelocissus Planch. (115) |
| Vitis L. (75) |
Cissus is the largest genus in the family with 300 species of complex classification [8]. Cyphostemma is second, with 200 species of an interesting diversity in their range of distribution as well as in growth habits (vines and lianas, herbs, stem succulents and a tree) [9]. Vitis has seventy-five inter-fertile wild species distributed in three continents under subtropical, Mediterranean and continental-temperate climatic conditions. Vitis vinifera L. is the species with highest economic importance in the family with some taxonomic uncertainty about the differentiation between V. vinifera L. subsp. vinifera and V. vinifera L. subsp. sylvestris (Willd.) Hegi [10][11]. Thousands of cultivars of V. vinifera are used worldwide in Viticulture. Species of other genera are widely cultivated, such as Parthenocissus quinquefolia (L.) Planch., the Virginia creeper, in temperate areas, and Cissus incisa Des Moul., the grape ivy, in tropical areas. Species of the genus Tetrastigma are the only host plants for the parasitic plant Rafflesia arnoldii R.Br., Rafflesiaceae, which is native only to a few areas within the Malay Archipelago [12].
2. Seed Morphology in the Vitaceae
2.1. Quantification of Seed Shape by Geometric Models
2.2. Seed Morphology in the Vitaceae
| Tribe (Species Observed/Total) |
Genera (Species Observed/Total) |
Species (References for the Images) |
|---|---|---|
| I. Ampelopsideae (15/47) |
Ampelopsis Michx. (13/18) |
Ampelopsis aconitifolia [33], A. arborea [34], A. bodinieri [33], A. cantoniensis [35][33], A. cordata [36], A. chaffanjoni [33], A. delavayana [35], A. denudata [29], A. glandulosa [37], A. grossedentata [35], A. humulifolia [33], A. japonica [33], A. megalophylla [35][33] |
| Rhoicissus Planch. (2/14) |
Rhoicissus revoilii [35], R. rhomboidea [35] | |
| II. Cisseae (33/300) |
Cissus L. (33/300) |
Cissus antarctica [35], C. aralioides [32][38], C. barbeyana [38], C. bosseri [38], C. cactiformis [38], C. campestris [35][39][40], C. cornifolia [38], C. descoingsii, [35][39], C. diffusiflora [38], C. elongata [38], C. erosa [41], C. floribunda [38], C. fuliginea [35], C. granulosa [35], C. hastata [38], C. hypoglauca [35], C. integrifolia [38][40], C. leucophlea [38], C. penninervis [35], C. petiolata [38], C. pileata [38], C. populnea [38], C. quadrangularis [42], C. reniformis [35][39], C. repens [38], C. sciaphila [38], C. smithiana [38], C. sterculiifolia [35], C. subtetragona [38], C. trianae [35], C. tuberosa [40], C. verticillata [35][39][40][43], C. willardii [40], |
| III. Cayratieae (40/365) |
Causonis Raf. (1/9) | Causonis sp. [44] |
| Cayratia Juss. (7/60) |
Cayratia cheniana [44], C. geniculata [35], C. imerinensis [45], C. japonica [35][46], C. oligocarpa [35], C. saponaria [35], C. sp. [African, [44]] | |
| Cyphostemma (Planch.) Alston (3/200) |
Cyphostemma elephantopus [47], C. laza [35], C. junceum [35] | |
| Pseudocayratia J. Wen, L.M.Lu and Z.D.Chen (3/5) |
Pseudocayratia dichromocarpa [48], P. pengiana [48], P. speciosa [48][49] | |
| Tetrastigma (Miq.) Planch. (26/100) |
Tetrastigma campylocarpum [50], T. cauliflorum [50], T. caudatum [50], T. delavayi [50], T. dichotomum [49], T. formosanum [50], T. harmandi [35], T. hemsleyanum [35][50], T. henryi [50], T. hypoglaucum [50], T. jinghongense [50], T. kwangsiense [29][35], T. lanceolarium [29], T. laoticum [50], T. obovatum [49][50], T. obtectum [49][50], T. pachyllylum [50], T. pedunculare [35][49][50], T. petraeum [50], T. retinervum [50], T. rumicispermum [35][49][50], T. serrulatum [50], T. sichouense [50], T. thorsborneorum [50], T. triphyllum [35][50], T. xishuangbannaense [35][50] | |
| IV. Parthenocisseae (11/16) |
Parthenocissus Planch. (9/14) |
Parthenocissus dalzielii [33], P. heptaphylla [35], P. henryana [33], P. heterophylla [33], P. himalayana [51], P. laetevirens [33], P. quinquefolia [52], P. tricuspidata [33][34][43], P. vitacea [35] |
| Yua C.L.Li (2/2) | Yua austro-orientalis [35], Y. chinensis [35] | |
| V. Viteae (32/190) |
Ampelocissus Planch. (13/115) |
Ampelocissus acapulcensis [29], A. bombycina [29], A. bravoi [40], A. cavicaulis [29], A. erdvendbergiana [29], A. grantii [29], A. javalensis [29][40], A. latifolia [29], A. macrocirrha [29], A. martinii [40], A. obtusata [29], A. ochracea [29], A. robinsonii [29] |
| Vitis L. (19/75) |
Vitis aestivalis [53], V. amurensis [43][54], V. brandoniana [52], V. cinerea [55], V. eolabrusca [52], V. flexuosa [52], V. grayensis [56], V. labrusca [43][52][57], V. lanatoides [56], V. latisulcata [56], V. palmata [58], V. pseudorotundifolia [52], V. rostrata [52], V. rotundifolia [35][39][52], V. rupestris [43], V. tiliifolia [59], V. tsoi [35][39], V. vulpina [57][60], V. wilsoniae [35][39] |
Table 3. A summary of groups based on morphological seed types for the analysed species in the Vitaceae. The number of cases found in each group is given between dashes.
| Group (Geometric Model) | Examples |
|---|---|
| Group I (Lenses)-3- | Cissus quadrangularis [42], C. sterculiifolia [35], Tetrastigma petraeum [50] |
| Group II (Superellipses)-7- | Ampelocissus bravoi [40], C. reniformis [35][39], Cyphostemma elephantopus [47], C. laza [35], Tetrastigma campylocarpum [50], T. caudatum [50], T. henryi [50] |
| Group III (Elongated water drops)-15- | Ampelopsis arborea [34], Cayratia imerinensis [45], Cissus aralioides [32][38], C. cornifolia [38], C. erosa [41], C. integrifolia [38][40], C. petiolata [38], C. pileata [38], C. populnea [38], C. verticillata [35][39][40][43], C. sciaphila [38], C. smithiana [38], C. willardii [40], Cyphostemma junceum [35], V. vulpina [57][60] |
| Group IV (Water drops, normal or rounded)-14- | Ampelopsis bodinieri [33], A. glandulosa [33][37], A. humulifolia [33], Cayratia cheniana [44], Cissus campestris [35][39][40], C. fuliginea [35], C. tuberosa [40], C. granulosa [35], Parthenocissus dalzielii [33], Tetrastigma triphyllum [35][50], Vitis amurensis [43][54], V. labrusca [43][52][57], V. palmata [58], V. rupestris [43] |
| Group V (Heart curves normal or rounded)-19- | Ampelopsis aconitifolia [33], A. chaffanjoni [33], A. cordata [36], A. japonica [33], Parthenocissus heptaphylla [35], P. heterophylla [33], P. henryana [33], P. himalayana [50][51], P. quinquefolia [52], P. vitacea [35], P. tricuspidata [33][34][43], Rhoicissus revoilii [35], T. lanceolarium [29], Vitis cinerea [55], V. flexuosa [52], V. lanatoides [56], V. latisulcata [56], V. tsoi [35][39], V. wilsoniae [35][39] |
| Group VI (Elongated Heart curves)-6- | Ampelocissus acapulcensis [29], Cissus oligocarpa [35]. V. eolabrusca [52], V. grayensis [56], V. pseudorotundifolia [52], V. tiliifolia [59] |
| Group VII (Other elongated types)-11- | Ampelopsis megalophylla [35][33], Causonis sp. [44], Cayratia saponaria [35], Cissus trianae [35], C. hypoglauca [35], Parthenocissus laetevirens [33], T. hypoglaucum [50] Vitis aestivalis [53], V. rotundifolia [35][39][52], Yua austro-orientalis [35], Y. chinensis [35] |
| Group VIII (Heart curves of the Cayratia and Pseudocayratia types)-7- | Cayratia japonica [35][46], Cayratia sp. [African, [44]], Pseudocayratia dichromocarpa [48], P. pengiana [48], P. speciosa [48][50], Tetrastigma formosanum [49], T. pedunculare [35][49][50] |
| Group IX (Heart curves of the SqHC type of Ampelocissus and Ampelopsis)-15- | Ampelocissus bombycina [29], A. cavicaulis [29], A. erdvendbergiana [29], A. grantii [29], A. javalensis [29][40], A. latifolia [29], A. macrocirrha [29], A. martinii [40], A. obtusata [29], A. ochracea [29], A. robinsonii [29], Ampelopsis cantoniensis [35][33], A. delavayana [35], A. denudata [29], A. grossedentata [35] |
| Group X Elongated Superellipse-heart curves-16- | Cissus elongata [38], C. penninervis [35], Rhoicissus rhomboidea [35], Tetrastigma hemsleyanum [35][50], T. jinghongense [50], T. laoticum [50], T. cauliflorum [50], T. dichotomum [49], T. harmandi [35], T. pachyllylum [50], T. kwangsiense [29][35], T. obovatum [49][50], T. obtectum [49][50], T. retinervum [49], T. serrulatum [50], T. sichouense [50] |
| Undefined-18- | Cayratia geniculata [35], Cissus antarctica [35], C. barbeyana [38], C. bosseri [38], C. cactiformis [38], C. descoingsii, [35][39], C. diffusiflora [38], C. floribunda [38], C. hastata [38], C. leucophlea [38], C. repens [38], C. subtetragona [38], T. delavayi [50], T. rumicispermum [35][49][50], T. thorsborneorum [50], T. xishuangbannaense [35][50], V. brandoniana [52], V. rostrata [52] |
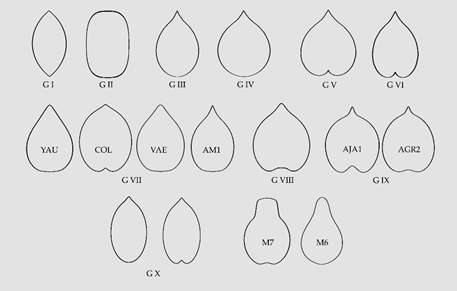
Figure 1. The images of seeds of Cissus sterculiifolia [35], Tetrastigma petraeum [49] and Cissus quadrangularis [42] resemble lenses of different proportions.
In general, the distribution of morphological types is not in close agreement with the current taxonomic classification; nevertheless, some results may be summarized in this aspect. First, the seeds of the Elongated Superellipse-Heart Curves (ESHCs) type (Group X) are more frequent in Tetrastigma and have been observed in Rhoicissus and Cissus, but not in species of other genera. While many seeds in species of Ampelopsis, Parthenocissus and Vitis share the typical shapes of water drop and heart curves, the squared heart curve (SqHC) type (Group IX) has been predominantly observed in Ampelocissus and Ampelopsis. A number of species remain undefined due to one of these two reasons: First, their irregular seed shape making difficult the identification of an adequate model (Cayratia geniculata, Cissus antarctica) and, second, the seed images have geometric shapes but the identification of the model with the corresponding equation is pending (Tetrastigma delavayi, T. rumicispermum). In addition, further work will be done on the seeds of Vitis species.
References
- The Angiosperm Phylogeny Group; Chase, M.W.; Christenhusz, M.J.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20.
- Ridsdale, C.E. A revision of the family Leeaceae. Blumea 1974, 22, 57–100.
- Wen, J.; Lu, L.-M.; Nie, Z.-L.; Liu, X.-Q.; Zhang, N.; Ickert-Bond, S.; Gerrath, J.; Manchester, S.R.; Boggan, J.; Chen, Z.-D. A new phylogenetic tribal classification of the grape family (Vitaceae). J. Syst. Evol. 2018, 56, 262–272.
- Zhang, N.; Wen, J.; Zimmer, E.A. Congruent deep relationships in the grape family (Vitaceae) based on sequences of chloroplast genomes and mitochondrial genes via genome skimming. PLoS ONE 2015, 10, e0144701.
- Zeng, L.P.; Zhang, N.; Zhang, Q.; Endress, P.K.; Huang, J.; Ma, H. Resolution of deep eudicot phylogeny and their temporal diversification using nuclear genes from transcriptomic and genomic datasets. New Phytol. 2017, 214, 1338–1354.
- Wen, J. Vitaceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin, Germany, 2007; pp. 467–479.
- Planchon, J.E. Monographie des Ampélidées vrais. In Monographiae Phanaerogamarum 5; de Candolle, A.P., de Candolle, A.C.P., Eds.; Masson: Paris, France, 1887; pp. 305–654.
- Rossetto, M.; Jackes, B.R.; Scott, K.D.; Henry, R.J. Is the genus Cissus (Vitaceae) monophyletic? Evidence from Plastid and Nuclear Ribosomal DNA. Syst. Bot. 2002, 27, 522–533.
- Hearn, D.J.; Evans, M.; Wolf, B.; McGinty, M.; Wen, J. Dispersal is associated with morphological innovation, but not increased diversification, in Cyphostemma (Vitaceae). J. Syst. Evol. 2018, 56, 340–359.
- Terral, J.F.; Tabard, E.; Bouby, L.; Ivorra, S.; Pastor, T.; Figueiral, I.; Picq, S.; Chevance, J.-B.; Jung, C.; Fabre, L.; et al. Evolution and history of grapevine (Vitis vinifera) under domestication: New morphometric perspectives to understand seed domestication syndrome and reveal origins of ancient European cultivars. Ann. Bot. 2010, 105, 443–455.
- This, P.; Lacombe, T.; Thomas, M.R. Historical origins and genetic diversity of wine grapes. Trends Genet. 2006, 22, 511–519.
- Encyclopedia Britannica. Vitales. Available online: https://www.britannica.com/plant/Vitales (accessed on 25 July 2021).
- Gutiérrez del Pozo, D.; Martín-Gómez, J.J.; Tocino, Á.; Cervantes, E. Seed geometry in the Arecaceae. Horticulturae 2020, 6, 64.
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. Available online: http://imagej.nih.gov/ij/docs/guide (accessed on 20 January 2021).
- Cervantes, E.; Martín-Gómez, J.J.; Saadaoui, E. Updated methods for seed shape analysis. Scientifica 2016, 5691825.
- Cervantes, E.; Martín-Gómez, J.J. Seed shape description and quantification by comparison with geometric models. Horticulturae 2019, 5, 60.
- Cervantes, E.; Martín-Gómez, J.J.; del Pozo, D.G.; Silva Días, L. An angiosperm species dataset reveals relationships between seed size and two-dimensional shape. Horticulturae 2019, 5, 71.
- Cervantes, E.; Martín-Gómez, J.J.; Ardanuy, R.; de Diego, J.G.; Tocino, Á. Modeling the Arabidopsis seed shape by a cardioid: Efficacy of the adjustment with a scale change with factor equal to the Golden Ratio and analysis of seed shape in ethylene mutants. J. Plant. Physiol. 2010, 167, 408–410.
- Martín Gómez, J.J.; Tocino, Á.; Ardanuy, R.; de Diego, J.G.; Cervantes, E. Dynamic analysis of Arabidopsis seed shape reveals differences in cellulose mutants. Acta Physiol. Plant. 2014, 36, 1585–1592.
- Cervantes, E.; Martín-Gómez, J.J.; Chan, P.K.; Gresshoff, P.M.; Tocino, Á. Seed shape in model legumes: Approximation by a cardioid reveals differences in ethylene insensitive mutants of Lotus japonicus and Medicago truncatula. J. Plant Physiol. 2012, 169, 1359–1365.
- Saadaoui, E.; Martín-Gómez, J.J.; Cervantes, E. Intraspecific variability of seed morphology in Capparis spinosa L. Acta Biol. Cracov. Bot. 2013, 55, 99–106.
- Saadaoui, E.; Martín-Gómez, J.J.; Tlili, N.; Khaldi, A.; Cervantes, E. Effect of climate in seed diversity of wild Tunisian Rhus tripartita (Ucria) Grande. J. Adv. Biol. Biotechnol. 2017, 13, 1–10.
- Martín-Gómez, J.J.; Saadaoui, E.; Cervantes, E. Seed shape of castor bean (Ricinus communis L.) grown in different regions of Tunisia. J. Agric. Ecol. Res. Int. 2016, 8, 1–11.
- Saadaoui, E.; Martín, J.J.; Bouazizi, R.; Chokri, B.R.; Grira, M.; Abdelkabir, S.; Khouja, M.L.; Cervantes, E. Phenotypic variability and seed yield of Jatropha curcas L. introduced to Tunisia. Acta Botánica Mex. 2015, 110, 119–134.
- Martín-Gómez, J.J.; Rewicz, A.; Goriewa-Duba, K.; Wiwart, M.; Tocino, Á.; Cervantes, E. Morphological description and classification of wheat kernels Based on geometric models. Agronomy 2019, 9, 399.
- Cervantes, E.; Martín-Gómez, J.J. Seed shape quantification in the order Cucurbitales. Mod. Phytomorphol. 2018, 12, 1–13.
- Martín-Gómez, J.J.; Rewicz, A.; Cervantes, E. Seed shape diversity in families of the order Ranunculales. Phytotaxa 2019, 425, 193–207.
- Martín-Gómez, J.J.; del Pozo, D.G.; Cervantes, E. Seed shape quantification in the Malvaceae reveals cardioid-shaped seeds predominantly in herbs. Botanica 2019, 25, 21–31.
- Chen, I.; Manchester, S.R. Seed morphology of modern and fossil Ampelocissus (Vitaceae) and implications for phytogeography. Am. J. Bot. 2007, 94, 1534–1553.
- Trias-Blasi, A.; Parnell, J.A.N.; Chayamarit, K. Cissus amplexicaulis (Vitaceae), a new endemic species from Thailand. Kew Bull. 2010, 65, 487–490.
- Kew Science. Plants of the World Online. Cissus aphylla Chiov. Available online: http://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:869770-1 (accessed on 9 July 2021).
- Kew Science. Plants of the World Online. Cissus aralioides (Welw. ex Baker) Planch. Available online: http://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:67514-1 (accessed on 9 July 2021).
- Latiff, A. Studies in Malesian Vitaceae VII. The genus Tetrastigma in the Malay Peninsula. Gard. Bull. Singap. 1984, 36, 213–228.
- Manchester, S.R.; Kapgate, D.K.; Wen, J. Oldest fruits of the grape family (Vitaceae) from the Late Cretaceous Deccan Cherts of India. Am. J. Bot. 2013, 100, 1849–1859.
- Chen, I.; Manchester, S.R. Seed morphology of Vitaceae. Int. J. Plant. Sci. 2011, 172, 1–35.
- Native Plant Trust: Go Botany. Ampelopsis cordata Michx. Available online: https://gobotany.nativeplanttrust.org/species/ampelopsis/cordata/ (accessed on 9 July 2021).
- Norfolk Cottage Garden Seeds & Plants. Ampelopsis glandulosa var. Brevipedunculata. Available online: https://www.norfolkcottagegarden.co.uk/images/seeds/macro/ampelopsis-glandulosa-var-brevipedunculata_seed-macro_img148_960px.jpg (accessed on 9 July 2021).
- Adams, N.F.; Collinson, M.E.; Smith, S.Y.; Bamford, M.K.; Forest, F.; Malakasi, P.; Marone, F.; Sykes, D. X-rays and virtual taphonomy resolve the first Cissus (Vitaceae) macrofossils from Africa as early-diverging members of the genus. Am. J. Bot. 2016, 103, 1657–1677.
- Chen, I. History of Vitaceae Inferred from Morphology-Based Phylogeny and the Fossil Record of Seeds. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2009.
- Manchester, S.R.; Chen, I.; Lot, T.A. Seeds of Ampelocissus, Cissus and Leea (Vitales) from the Paleogene of Western Peru and their Biogeographic significance. Int. J. Plant. Sci. 2012, 173, 933–943.
- Smithsonian Tropical Research Institute. Cissus Erosa Subsp Erosa. Available online: https://biogeodb.stri.si.edu/bioinformatics/dfm/metas/view/7894 (accessed on 9 July 2021).
- DELTA—Description Language for Taxonomy. Cissus quadrangularis. Available online: https://www.delta-intkey.com/famfs/images/vitace03.jpg (accessed on 9 July 2021).
- Martín-Gómez, J.J.; Gutiérrez del Pozo, D.; Ucchesu, M.; Bacchetta, G.; Cabello Sáenz de Santamaría, F.; Tocino, Á.; Cervantes, E. Seed morphology in the Vitaceae based on geometric models. Agronomy 2020, 10, 739.
- Lu, L.; Wen, J.; Chen, Z. Cayratia cheniana (Vitaceae): An endangered new species endemic to the limestone mountains of Ninh Thuan Province, Vietnam. Syst. Bot. 2016, 41, 49–55.
- Descoings, B. Flore de Madagascar et des Comores (Plantes Vasculaires) 124 Famille—Vitacées; Museum National d’Histoire Naturelle: Paris, France, 1967; Available online: https://www.delta-intkey.com/angio/images/cayratia.gif (accessed on 9 July 2021).
- Jiang, H.; Yang, J.; Liang, T.; Zhang, Z.; Wang, S.; Qi, X.; Sheng, P. Palaeoethnobotanical analysis of plant remains discovered in the graveyard of the Haihun Marquis, Nanchang, China. Veg. Hist. Archaeobot. 2021, 30, 119–135.
- The World of Plants: Cyphostemma elephantopus. Available online: https://public.fotki.com/PanosS/vitaceae-pages/cyphostemma/cyphostema_elephant.html (accessed on 9 July 2021).
- Wen, J.; Lu, L.-M.; Hsu, T.-W.; Dang, V.-C.; Habib, S.; Boggan, J.K.; Okada, H.; Chen, I.-J.; Chen, Z.-D. Pseudocayratia, a new genus of Vitaceae from China and Japan with two new species and three new combinations. J. Syst. Evol. 2018, 56, 374–393.
- Habib, S.; Dang, V.-C.; Ickert-Bond, S.M.; Wen, J.; Chen, Z.-D.; Lu, L.-M. Evolutionary trends in Tetrastigma (Vitaceae): Morphological diversity and taxonomic implications. J. Syst. Evol. 2018, 56, 360–373.
- Habib, S.; Dang, V.-C.; Ickert-Bond, S.M.; Zhang, J.-L.; Lu, L.-M.; Wen, J.; Chen, Z.-D. Robust phylogeny of Tetrastigma (Vitaceae) based on ten plastid DNA regions: Implications for infrageneric classification and seed character evolution. Front. Plant. Sci. 2017, 8, 590.
- Norfolk Cottage Garden Seeds and Plants. Parthenocissus himalayana. Available online: https://www.norfolkcottagegarden.co.uk/images/seeds/macro/parthenocissus-himalayana-syn-semicordata_seed-macro_518_960px.jpg (accessed on 9 July 2021).
- Tiffney, B.H.; Barghoorn, E.S. Fruits and seeds of the Brandon lignite. I. Vitaceae. Rev. Palaeobot. Palynol. 1976, 22, 169–191.
- Plants USDA. Vitis aestivalis. Available online: https://plants.sc.egov.usda.gov/home/plantProfile?symbol=VIAE (accessed on 9 July 2021).
- Kirkbride, J.H., Jr.; Gunn, C.R.; Dallwitz, M.J. 2000 Onwards. Family Guide for Fruits and Seeds: Descriptions, Illustrations, Identification, and Information Retrieval; Version: 12th April 2021. Available online: https://www.delta-intkey.com/famfs/images/vitace04.jpg. (accessed on 9 July 2021).
- Plants USDA. Vitis cinerea. Available online: https://plants.sc.egov.usda.gov/home/plantProfile?symbol=VICI2 (accessed on 9 July 2021).
- Gong, F.; Karsai, I.; Liu, Y.-S. Vitis seeds (Vitaceae) from the late Neogene Gray Fossil Site, northeastern Tennessee, USA. Rev. Palaeobot. Palynol. 2010, 162, 71–83.
- Ardenghi, N.M.G.; Galasso, G.; Banfi, E.; Cauzzi, P. Vitis ×novae-angliae (Vitaceae): Systematics, distribution and history of an “illegal” alien grape in Europe. Willdenowia 2015, 45, 197–207.
- Plants USDA. Vitis palmata. Available online: https://plants.sc.egov.usda.gov/home/plantProfile?symbol=VIPA7 (accessed on 9 July 2021).
- Smithsonian Tropical Research Institute. Vitis tiliifolia. Available online: https://biogeodb.stri.si.edu/bioinformatics/dfm/metas/view/10813?&lang=es (accessed on 9 July 2021).
- Plants USDA. Vitis vulgaris. Available online: https://plants.sc.egov.usda.gov/home/plantProfile?symbol=VIVU (accessed on 9 July 2021).

