2. Mechanisms of Phosphorus Uptake
Phosphorus is taken up by plant roots from the soil solution in the form of H
2PO
4−1 ions (P
i), the concentration of which ranges from 1 to 100 µM. In the plant root tissues, its concentration is many times higher, ranging from 5 to 20 mM
[20]. A P
i concentration below 10 µM in the growth medium is considered as critical for the growth of crop plants
[21]. The uptake of P
i by a plant from the soil solution is a complex, multi-phase process. The two basic stages are:
- (1)
-
Passive—flow of Pi anions in the soil solution towards the plant root surface;
- (2)
-
Active—transfer of the Pi ions across the plasmalemma of the cortical root cell.
The potential of the soil to feed the crop plant with P results from:
- (1)
-
Pi concentration in the soil solution;
- (2)
-
Root absorption surface;
- (3)
-
Transport rate of Pi ions from the soil solution to the root surface;
- (4)
-
The rate of Pi ions’ incorporation into the plant’s metabolic processes.
The accumulation of P
i ions on the root surface depends on the rate of their diffusion (I
d, µg cm
−2 s
−1) in the soil solution
[22]. This process is subject to Fick’s law, according to which the rate of ions’ movement in the solution depends on both their concentration gradient (dc × dx
−1, µg cm
−3 cm
−1) and the value of the diffusion coefficient (D
eff, cm
2 s
−1):
A necessary condition for this process to occur is a plant, or more precisely, a plant root in the soil. The sufficient condition is the content of P
i ions in the appropriate concentration in the soil solution. The plant root is the factor that changes the initial concentration of ions in the soil solution, creating a concentration gradient toward the root and thus inducing their flow. According to the mechanistic model of P uptake by a plant root, the amount of the absorbed nutrient depends on the root surface area
[23]. This plant’s characteristics are determined by the rate of root growth into the soil and its diameter. Therefore, one of the most important factors determining the P nutrition of a crop plant is the size of the root system, defined by the root density and the plant’s effective rooting depth. This trait of the crop plant is genetically determined, but at the same time, it shows a strong response to environmental (soil, water content, temperature) and physiological factors (nutritional state of the plant)
[24][25]. Among the soil factors, the most important are the concentration of P
i in the soil solution and the value of its diffusion coefficient. The value of the P
i coefficient in the soil solution at field capacity is in the range of 10
−9 cm
2 s
−1. For comparison, the N–NO
3 coefficient is at least 1000 times greater and amounts to 1 × 10
−6 cm
2 s
−1 [26].
Two aspects of P
i uptake by crop plants require special attention from the farmer. The first is the impact of soil moisture on the value of P
i coefficient. Variable water content during the growing season is a natural phenomenon. It affects most severely the topsoil, which is rich in P (at least compared with the subsoil). As a consequence, this soil layer becomes heavily overgrown with roots
[27]. The decrease in water content in the soil, in turn, increases the contribution of capillary pores filled with air. These conditions lead to a decrease in the values of diffusion coefficients, the strongest for P
i. As a consequence, the amount of P reaching the plant root surface decreases (
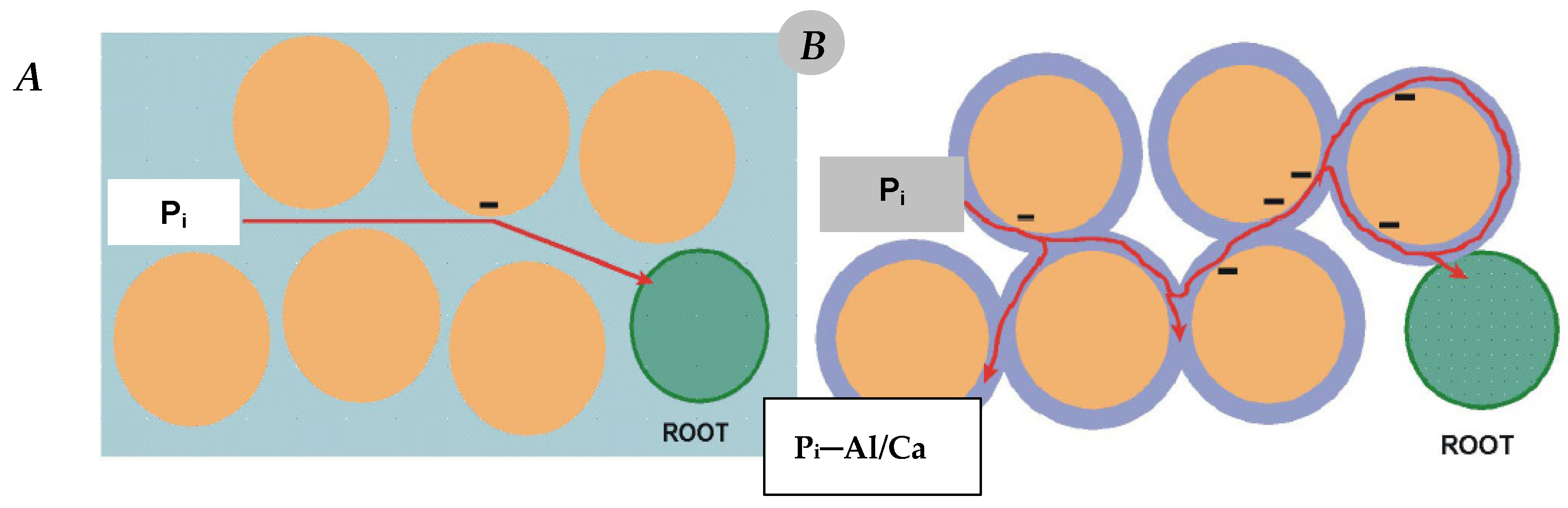
Figure 1).
Figure 1. Mobility and availability of inorganic phosphorus in the soil solution under optimal ((A) field capacity) and suboptimal water conditions ((B) water deficiency-induced precipitation by Al3+ or Ca2+); the red line indicates the direction of orthophosphate ion flow in the soil solution.
The seemingly small reduction in the water content from 27% to 22% resulted in a drastic decrease in the length of maize roots, consequently leading to a significant reduction in P uptake by plants
[28]. The negative impact of water shortage on P supply to the plant can be at least partially mitigated by maintaining a higher content of available P in the soil solution. Most crop plants take up and accumulate large amounts of P during the initial growth phase, usually in optimal soil water conditions
[29][30]. The high P uptake by the plant during this period, which is only apparently luxurious, increases its tolerance to water stress in the later stages of growth, which are crucial for yield formation. A classic example is maize. Phosphorus deficiency in the fourth (fifth) leaf stage, in which the structure of generative organs is formed, is reflected in the flowering stage
[31]. This state is manifested by poor development or even the complete disappearance of the cob (
Figure 2).
Figure 2. Drastic phosphorus deficiency in the early stage of maize development (A) leads to no or poor cob formation (B). (Source: Witold Grzebisz).
The second aspect of supplying a plant with P is related to the supply of nitrate nitrogen (N–NO
3). This molecule behaves like a plant morphogen
[32]. The decreasing content of N–NO
3 in the growth medium stimulates the growth of main roots (primary roots, seminal roots) into new areas of soil and an accompanying reduction in the growth of lateral roots. Opposite effects are observed when the content of N-NO
3 increases in the growth medium
[33]. N-NO
3 ions are very labile in the soil solution, so these resources are quickly depleted, forcing the plant to penetrate new areas of the soil, rich in both water and nitrates. As a result of these processes, the plant abandons previously exploited soil. As a consequence of these processes, the use of freshly applied phosphorus does not exceed 5% (maximum 20%). However, the actual use of P
i from the soil solution can reach even 60%
[34][35].
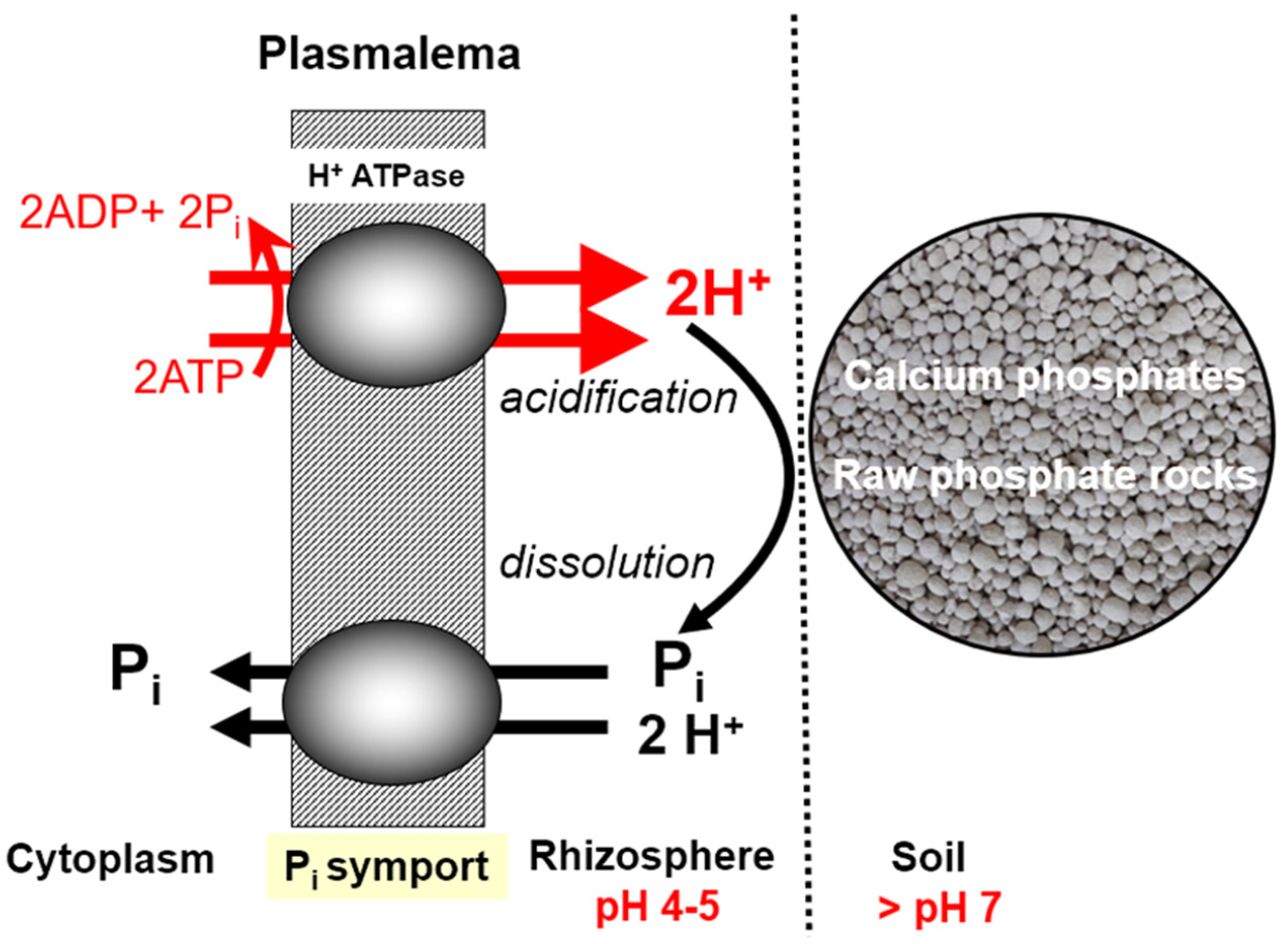
The process of P uptake from the soil solution requires the plant to use metabolic energy, the main source of which is the ATP (adenosine triphosphate) molecule. Its basic components are P and N, so there is a close interaction of both nutrients at the primary level for the functioning of a living organism
[36]. The paradox of this process is that the plant efficiently takes up P
i from the soil solution, provided it is in a good nutritional P state (
Figure 3). The ion H
2PO
4−1 that is to be transported across the cytoplasmatic membrane must first be protonated {[2H
+ × H
2PO
4−1]
+}. This process requires energy provided by ATP
[37]. A necessary condition for the stabilization of these processes is a constant, stable content of P in the cell cytoplasm. The P content in other compartments of the cell and plant organs is variable. The greatest variability occurs in the vacuole
[12].
Figure 3. Hypothetical model of Pi ions uptake by a legume plant from soil, in a neutral soil reaction.
3. Response Strategies of Crop Plants to Pi Deficiency
Plants usually show a high level of adaptation to changes in P supply, from the molecular, through physiological, and finally morphological levels. Root epidermal cells, induced by a signal from the shoot, synthesize and release acid phosphatases into the rhizosphere. Enzymes from the PAPS (3′-Phosphoadenosine-5′-phosphosulfate) family operate in the soil at pH of 4 to 7.7 and in a wide range of temperature of 25–60 °C
[38]. The secretion of acid phosphatases by roots of P-deficient barley plants into the rhizosphere is in principle greater than the number of intracellular phosphatases produced in shoots
[39]. These enzymes hydrolyze P
i from organic soil P compounds
[40]. Plants deficient in P also excrete into the rhizosphere other organic compounds, which may constitute 10–25% of total fixed carbon. The synthesis of PAPs by soybean and common bean roots is considered to be a tolerance mechanism to the low P
i concentration in the soil P
i [41]. Some plants, mainly cereals, including maize, oil rapeseed, alfalfa, and chickpea secrete organic acids in response to two factors, i.e., P deficiency and Al toxicity
[41]. It is well recognized that Al cations, depending on the oxidation state, are active in P
i absorption or fixation, lowering their concentration in the soil solution. The root’s secretions are dominated by organic acids, mainly citric, malate, and oxalate (
Table 1). The main geochemical role of these compounds is to solubilize P
i from Ca, Al, and Fe phosphates. Citric acid has the highest efficiency in releasing P
i from organic-P
i-esters
[40].
Table 1. Diversity of organic acid produced by PSMs (phosphate solubilizing microbes).
| PSM Isolates |
Organic Acids |
References |
| Pseudomonas |
Citric acid, succinic acid, fumaric acid, gluconic acid, 2-ketogluconic acids |
[42][43] |
| Bacillus sp. |
Citric acid, malic acid, succinic acid, fumaric acid, tartaric acid, gluconic acid |
[42] |
| Proteus sp. |
Citric acid, succinic acid, fumaric acid, gluconic acid |
[42] |
| Azospirillum sp. |
Citric acid, succinic acid, fumaric acid, gluconic acid |
[42] |
| Aspergillus |
Citric acid, gluconic acid, oxalic acid, succinic acid, malic acid, glycolic acid |
[44] |
| Penicillium sp. |
Gluconic acid, glycolic acid, succinic acid, malic acid, oxalic acid, citric acid |
[44] |
| Erwinia herbicola |
Gluconic acid, 2-ketogluconic acid |
[43] |
Another mechanism of plant response to moderate P deficiency is progressive infection and symbiosis with mycorrhizal fungi
[45]. The latter phenomenon does not include plants from the
Brassicaceae and
Chenopodiacae families
[46][47]. Therefore, for optimal growth, these plants require soil with a pH in the neutral range, at least. In soil solution with a pH on the neutral/alkaline border, the P mineral called brushite dominates
[47]. The hypothetical effect of a rhizosphere acidifying plant (families Brassicaceae, Fabaceae) is as follows:
Mycorrhiza is a common phenomenon occurring between two kingdoms, plants and fungi, involving mutual symbiosis. Both ectotrophic and endotrophic (arbuscular) mycorrhizal fungi (AMF) are important for plants’ growth and development
[48]. This group of fungi belongs to the phylum
Glomeromycota [49] and is present in over 90% of land plants
[50]. AMF fungal hyphae germinating in the root cortex come into physical contact with the host plant. They elongate and grow intercellularly through appressoria, and penetrate the cortex root cells, where they form arbuscules
[51]. At the same time, the fungus forms extracellular hyphae that grow into the soil, thereby increasing the root surface area. Their main function is the absorption of water and nutrients, mainly P and N
[52]. The fungal hyphae are 100 times smaller in diameter than the smallest root and 10–20 times smaller than the diameter of the root hair. In drought conditions, when the hydraulic potential of the soil is very low, fungal hyphae participate in water uptake
[53]. AMF have a key role in phosphate uptake particularly in P-deficient soils by mobilizing P from rock phosphate
[54]. Hyphae exudates can decompose larger soil organic molecules, containing both N and P compounds
[55]. The transfer of N from organic matter to plant tissues by AMF hyphae has been documented to increase plant biomass
[56].
Severe P deficiency in a crop plant, i.e., P starvation, leads to a reduction in the total root system size
[57]. However, it is assumed that in conditions of P deficiency below 1 µM in the growth medium, some plants form cluster roots (proteid roots), which are a specific superabundance of root hairs or a tertiary form of lateral roots. Moreover, most plants of these species do not form mycorrhizal associations. This type of adaptation mainly involves natural plants, and the classic examples are plants from the Proteaceae family. Among crop plants, cluster roots form on pumpkin, white lupine, yellow lupine, and macadamia. These roots excrete huge amounts of protons, carboxylates, and also acid phosphatases into the narrow zone around the root (around 1 mm). As a result of this dual root activity, P
i is released both from inorganic and organic P compounds
[58][59].
Farmers should consider three practical solutions based on the P uptake characteristics of crops grown on the farm. The first concerns the type of plant’s response to P deficiency. Based on the response of crop plants to low P deficiency, it was found that in the group of plants with fibrous root systems, such as cereals, the morphological type of adaptation dominates. In contrast, legumes show a dominant physiological type of adaptation, involving the secretion of protons and organic acids into the rhizosphere
[60]. In the case of cruciferous plants, three interacting types of adaptation should probably be considered. These plants can be classified as morphologically adapted as indicated by their large, extensive root systems [
Table 1;
[61]]. Moreover, the roots of these plants release organic compounds into the soil, in turn inducing the activity of soil microorganisms
[62]. On the other hand, cruciferous plants are usually grown on neutral or alkaline soil. In such geochemical conditions, phosphorus is in forms unavailable to plants. To absorb P, a plant must acidify the rhizosphere
[63]. As a consequence, legumes or cruciferous plants create a very favorable growth environment for the successor crop, which usually is cereal. Crop rotation, taking into account both groups of plants, is one of the most beneficial systemic solutions in P management on the farm.
4. Environmental Factors Controlling Phosphorus Uptake by Crop Plants
4.1. Temperature
The uptake of P
i from the soil solution and, thus, the growth and development of a crop plant are affected by many environmental factors
[64]. Of these, the key ones are temperature and water. The soil temperature is usually lower than the air temperature, but its fluctuation has a significant impact on the content of water, which determines the supply of nutrients to the root surface
[65]. Two temperature extrema are important to recognize the response of the plant’s root system, i.e., minimum and maximum. The root growth proceeds up to the optimal range of temperature for a given species
[66]. As reported by Fonseca de Lima et al.
[67], the optimum temperature for root growth of temperate cereals is in the range of 15–20 °C, for rice 25–28 °C, and tropical maize 25–35 °C. Extreme temperatures during the reproductive phase, i.e., during the final stages of yield formation, lead to a significant reduction in the grain weight
[68].
Plants are most sensitive to temperature extremes during juvenile stages of growth because the root system is not well developed. Exposure of crop plants to low temperature stress (LTS: chilling temperature range of 0–15 °C) significantly reduces the main traits of the root system architecture (RSA). For example, wheat seedlings exposed for 14 days to a temperature of 4 ± 1 °C decrease their absorption potential for uptake of nutrients
[69]. The key reason is the reduction in the root surface area. Tropical crops such as sugarcane are strongly sensitive to chilling stress. The drop in temperature below 4 °C results in root activity decrease in the range of 64–80% due to reductions in root length, root diameter, and root hair density, and also disturbance in root gravitropism
[70]. The slowdown in the rate of root growth of plants exposed to low temperatures is due to the reduced accumulation of auxin in the root apex, in turn repressing the rate of meristematic cell division. At the same time, the increasing content of cytokinin suppresses the growth of roots. The root system for an optimum elongation rate requires high auxin transport to the root apex and at the same time, a low content of cytokinin
[70][71].
The disturbance in both morphological and physiological traits of crop plants due to extreme soil temperatures results in significant reduction in water and nutrient uptake. The end effect is a drop in yield
[72]. The impact of extremely high temperatures on the RSA results in (i) a reduction in the length of the primary root, (ii) reductions in the growth rate and the number of lateral roots, (iii) an increase in the number of the second-and third-order roots, which have also a larger diameter, (iv) a lower angle of new, emerging lateral roots, (v) inhibition of the number and length of shoot-borne roots (adventitious and nodal roots—maize for example), (vi) an increase in the number and length of root hairs
[73]. The total reduction in the growth rate of the root system and its size results in a narrower root/shoot ratio, mainly due to a much stronger reduction in the shoot biomass compared with the root biomass. Moreover, every 10 °C increase in soil temperature results in the respiration rate doubling, leading to a strong loss of dry matter
[74]. The increased temperature activates calcium channels in the cytoplasmatic membrane, in turn increasing the flow of Ca
2+ ions into the cell and inducing heat shock transcription factors
[75]. This is important information for farmers working with sandy soils, which are naturally poor in calcium. These negative effects of extreme temperatures can be counterbalanced by increasing the supply of water, including through irrigation and phosphorus fertilizer
[76].
4.2. Water—Drought
In the classic hierarchy of yield-forming factors, water is considered the yield-limiting factor
[77]. Water controls the plant’s life processes at various levels of its organization
[78]:
- (1)
-
Cells: (i) water photolysis—water acts as a nutrient; (ii) a component of spatial structures of organic compounds, including proteins, carbohydrates, and fats; (iii) a component of the osmotic sap of cells;
- (2)
-
Tissues and organs: (i) connects cells to form tissues and then organs; (ii) determines the turgor of conducting cells (xylem); (iii) a critical component of assimilates transported in the phloem;
- (3)
-
Plant: controls (i) the optimum level of temperature through transpiration of water and thus the rate of metabolic processes, (ii) the movement of stomata during the circadian cycle, and (iii) CO2 uptake from the atmosphere.
Water flows in the soil–plant–atmosphere continuum according to the principle of decreasing water potential: from the soil (−0.03 to −1.5 MPa megapascal), through the plant root (−0.3 to −2.0 MPa), then arriving at leaves (−1.5 to −3.0 MPa), and finally to the atmosphere (−50 MPa)
[79]. The movement of water from the soil to the plant root is induced by hydrostatic root pressure, forcing water to move from the root pericycle to the xylem. Root pressure requires the expenditure of metabolic energy and therefore occurs only in the youngest, not finally formed tissues
[80]. For this reason, the plant’s water uptake depends on the potential and the physical properties of the growing roots to penetrate the soil. On the other hand, the physical properties of the soil (texture, structure, penetrance resistance) determine both the amount of water available to the plant and the conditions of root growth
[81]. During the growing season, under natural rainfall conditions, the amount of water available is variable, increasing its non-homogenous distribution in the soil profile. In conditions of increasing water shortage, intensified by drought, the growth of lateral roots takes place mainly in the water-rich soil paths
[82][83]. The mechanism of active search by lateral roots for water, called hydrotropism or hydro-patterning, is controlled by auxins in response to the difference in water potential in the roots. As a consequence, in dry parts of the soil body, the growth of lateral roots is drastically reduced. This phenomenon is called xerobranching
[84]. The response of the plant root morphological traits to drought is deeply associated with the type of root system. The rooting depth is the most conservative RSA trait because it does not undergo changes in response to drought
[81]. The most sensitive traits, responding negatively to drought regardless of the plant species, are the root length and root dry weight, whereas root hair length and root hair density show the opposite trend. Differences between key types of roots refer mainly to the root diameter. Plants with a tap root system show a clear response to water stress. As a result of water stress, the root/shoot ratio decreases, because the shoot biomass shows a much stronger reduction than the root biomass
[84][85][86].
In plant production, the expected yield is influenced not only by the plant’s total water demand and its efficiency (transpiration coefficient), but mainly due to its supply during the critical phases of yield formation
[87]. Too little water supply to the plant during this period leads to disruption of the yield-forming processes. In cereals, the maximum water depletion in the rooting zone should not exceed 40 (60)% in the period extending from the beginning of shooting to the beginning of flowering, while during the grain-filling period, it can even reach 70%
[88][89]. Maize, despite a relatively low value of transpiration coefficient compared with cereals (270–400 versus 500–600 dm
3 H
2O kg
−1 DW), is most sensitive to reduction in water content in the period from silking to the watery stage of kernel development (maximum water drop is 50%)
[90][91].
Reduction in the root system size of crop plants under conditions of water stress, which is frequently concomitant with the thermal stress, must therefore lead to disruption in P
i uptake by crop plants. The meta-analysis by He and Dijkstra
[92] showed a significantly higher reduction in P (−9.18%) than N (−3.73%) uptake by plants. The authors emphasized the significant impact of the length of the drought period and the role of drying–rewetting periods on N and P uptake by crop plants. In the authors’ opinion, short-term drought (<90 d) reduces N and P uptake more strongly than C assimilation by plants. The reduction in P uptake is due to the disruption of P flow towards the root, as well as the decrease in the content of P
i in the soil solution due to its precipitation and the decreased activity of microbes in the root rhizosphere
[93]. This situation is especially dangerous for short-growing crops, especially when drought occurs during the period of intensive P accumulation
[94].
Effective management of nutrients in world regions under seasonally changing weather conditions, including in-seasonal and irregular drought sequences, is difficult. At least two aspects need to be considered. The first, strategic one assumes maintaining a high content of available P in the rooting zone of cultivated plants, especially those sensitive to P supply during the critical period of yield formation. The second, tactical one concerns fertilizer treatments, assuming lower P uptake by plants in a dry season. In light of the strategy of sustainable P management, the question arises here whether an increase in P doses or the level of the soil P
i content can alleviate, at least partly, the effect of water shortage on crop yield. Increased doses of P under water stress in the critical period of wheat yield formation did not reduce but actually increased the yield gap, which doubled for the optimal P dose relative to the P control (0.81 → 1.65 g plant
−1). However, the plant was sufficiently supplied with P, as evidenced by a linear increase in the P content in both shoots and grain. Wheat plants under water stress, regardless of the P dose, contained more P in both straw and grain. The key difference between well-watered and water-stressed plants was the number of grains per plant, which was lower in plants subjected to water stress
[95]. This simple experiment allows us to formulate two general conclusions. The first is consistent with Körner’s hypothesis
[96] that nutrient resources, in this case referring to P, accumulated in the pre-flowering period by the crop plant will not be used when the sink capacity is too low. The second conclusion indicates the need to create growth conditions for at least full exploitation of P resources in the plant. This is possible, provided the sink is large
[35].
The long-term analysis of yields in temperate regions of the world indicates a specific year-to-year variability. After yield declines in a dry year, it is at least partially compensated in the subsequent year, provided that the weather conditions are better. The suggested yield compensation mechanism is quite evident in Poland
[97]. For example, the potential yield of winter wheat in 2019–2021 was 9.71 t ha
−1. In dry 2019, it decreased to 9.17 t ha
−1, and in 2020, it increased to 10.19 t ha
−1. In 2021, with standard weather conditions, it amounted to 9.77 t ha
−1. Study on maize and soybean response to the severe drought in the Upper Mississippi River Basin (USA) during the period 1960–2006 showed yield reductions of 27% and 20%, respectively
[98]. In the years with drought, the respective amounts of residual P were 11 ((ranging from 3 to 19) kg P ha
−1) and 3 ((ranging from 0 to 9) kg P ha
−1), for maize and soybean. The effect of land types, soil textural classes, and hydrologic groups was small. In normal years, the P carryover was much lower, amounting to 7 ± 1 kg P ha
─1 for maize and 2 ± 1 kg P ha
−1 for soybean.
 +1 credit
+1 credit