 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Nusrat Nabila Chowdhury | -- | 6155 | 2023-12-22 21:40:31 | | | |
| 2 | Wendy Huang | -132 word(s) | 6023 | 2023-12-26 02:32:03 | | |
Video Upload Options
The COVID-19 pandemic has had a profound impact on societies, public health, healthcare systems, and the world economy. Vaccination emerged as the most effective strategy to combat this infectious disease. For vaccination strategies, any conventional vaccine approach using attenuated live or inactivated/engineered virus, as well as other approaches, typically requires years of research and assessment. However, the urgency of the situation promoted a faster and more effective approach to vaccine development against COVID-19. The role of nanotechnology in designing, manufacturing, boosting, and delivering vaccines to the host to counter this virus was unquestionably valued and assessed.
1. Introduction
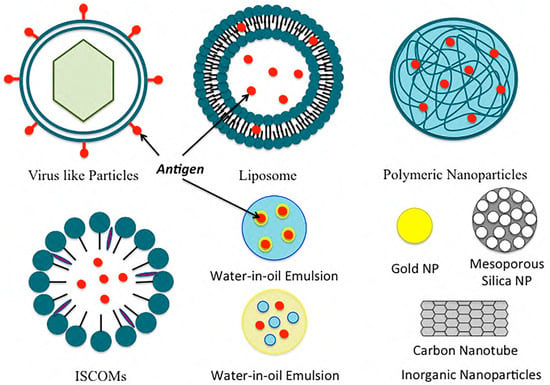
Figure 2. Examples of common nanoformulations used for vaccine delivery.
2. Nanoparticle Vaccine Adjuvants and Delivery Systems
2.1. Polymeric Nanoparticles
2.2. Inorganic Nanoparticles
2.3. Liposomes
2.4. Immunostimulatory Complexes (ISCOMs)
2.5. Emulsions
2.6. Virus-like Particles and Virosomes
3. The Vaccine Development Approach for Coronavirus
4. Conclusions
| Delivery System | Composition | Antigen | References |
|---|---|---|---|
| Polymeric-Based System | PLGA | OVA | Demento et al. [80] |
| PLGA, polylactic acid | Hepatitis B surface antigen | Thomas et al. [81] | |
| Lipid-coated PLGA | OVA | Bershteyn et al. [82] | |
| Lipid-coated PLGA | Malaria antigen | Moon et al. [83] | |
| Deacylated cationic polyethyleneimine | HIV CN54gp140 antigen | Mann et al. [84] | |
| Polylactic acid | Hepatitis B surface antigen | Saini et al. [85] | |
| Chitosan-coated polycaprolactone | H1N1 hemagglutinin | Gupta et al. [86] | |
| Polyanhydrides | Yersinia pestis antigen | Ulery et al. [87] | |
| Chitosan nanoparticles | HBsAg | Lugade et al. [88] | |
| Mannosylated chitosan nanoparticles | Recombinant hepatitis B virus surface antigen | Mehrabi et al. [5] | |
| Cholesteryl-conjugated pullulan | Clostridium botulinum type-A neurotoxin subunit antigen | Nochi et al. [89] | |
| N-trimethyl chitosan | OVA | Slutter et al. [90] | |
| Alginate nanoparticles | Diphtheria toxoid | Sarei et al. [8] | |
| Hyaluronic acid (HA), monophosphoryl lipid A (MPLA), aluminum salt (Alum) | Hepatitis B antigen | Moon et al. [91] | |
| Inorganic Nanoparticles | Gold nanoparticles | Escherichia coli-specific immunogenic antigens | Sanchez-Villamil et al. [92] |
| Gold nanoparticles | West Nile virus envelops protein | Niikura et al. [12] | |
| Carbon nanoparticles | Bovine serum albumin | Wang et al. [13] | |
| Carbon magnetic nanoparticles | Hen egg lysozyme | Schreiber et al. [93] | |
| Mesoporous silica nanoparticles | Schitosoma mansoni | Montalvo-Quiros et al. [94] | |
| Silica nanoparticle-based drug delivery system | H1N1 influenza hemagglutinin antigen | Neuhaus et al. [95] | |
| Alum | Combination of an influenza antigen | Knudsen et al. [96] | |
| Calcium phosphate nanoparticle | H1N1 hemagglutinin antigen | Morcol et al. [97] | |
| Liposomes | DOPC, DOPG, MPB | OVA | Moon et al. [27] |
| EPC, DOGS-NTA-Ni | His-tagged heat shock protein | Mašek et al. [98] | |
| Pegylated DDA, TDB | Ag85B-ESAT-6 | Kaur et al. [25] | |
| DDA, TDB | OVA | Milicic et al. [99] | |
| DDA, DSPC, cholesterol, TDB | Ag85B-ESAT-6 | McNeil et al. [100] | |
| DDA, TDB | Trivalent influenza vaccine | Rosenkrands et al. [101] | |
| DDA, TDB | Ag85B-ESAT-6 | Henriksen-Lacey et al. [102] | |
| DDA, DODA, TDB | Ag85B-ESAT-6 | Christensen et al. [103] | |
| Lecithin, cholesterol | Diphtheria toxoid | de Veer et al. [104] | |
| Immunostimulatory Complexes (ISCOMS) | Cholesterol, phospholipids, saponins | hemagglutinin antigen | Cox et al. [105] |
| ISCOMATRIX | HPV16 E6 and E7 recombinant bacterial fusion protein | Frazer et al. [37] | |
| Emulsion | MF59 | Recombinant meningococcal B protein | Brito et al. [106] |
| MF59 | Hemagglutinin | Calabro et al. [107] | |
| MF59 | Recombinant meningococcal B protein | Singh et al. [108] | |
| W805EC | OVA | Myc et al. [109] | |
| W805EC | OVA | Makidon et al. [110] | |
| GLA | Falciparum subunit | Lousada-Dietrich et al. [111] | |
| GLA-SE | Recombinant hemagglutinin | Treanor et al. [112] | |
| GLA-SE | Plasmodium vivax subunit | Lumsden et al. [113] | |
| Virus-Like Particles And Virosomes | Epaxal® (Crucell, Leiden, The Netherlands) A (H1N1) virosomes + inactivated hepatitis A virus | Bovier et al. [114] | |
| Inflexal® V (Crucell) Virosomes from three influenza strains: A (H1N1), A (H3N2), and B | Herzog et al. [115] | ||
| Nasalflu® (Berna Biotech, Bern, Switzerland) Virosomes from three influenza strains: A (H1N1), A (H3N2), and B + heat labile toxin adjuvant | Gluck et al. [116]; Mutsch et al. [117] | ||
| Invivac® (Solvay, Brussels, Belgium) Virosomes from three influenza strains: A (H1N1), A (H3N2), and B |
de Bruijn et al. [118]; de Bruijn et al. [119] | ||
| Epaxal® Junior (Crucell) A (H1N1) virosomes + inactivated hepatitis A virus | Bovier et al. [114]; Van der Wielen et al. [120] |
| Study Title | Clinical Trails Gov ID | Clinical Trial | Interventions |
|---|---|---|---|
| Training the Innate Immune System Against SARS-CoV-2 (COVID-19) Using the Shingrix Vaccine in Nursing Home Residents (NH-Shingrix) | NCT04523246 | Early Phase 1 | Biological: SHINGRIX (zoster vaccine Recombinant, adjuvanted) Drug: normal saline |
| A Study Assessing the Safety, Tolerability, Immunogenicity of COVID-19 Vaccine Candidate PRIME-2-CoV_Beta, Orf Virus Expressing SARS-CoV_2 Spike and Nucleocapsid Proteins | NCT05367843 | Phase 1 | Drug: PRIME-2-CoV_Beta |
| Phase 1 Study of Intranasal PIV5 COVID-19 Vaccine Expressing SARS-CoV-2 Spike Protein in Healthy Adults and Adolescents (CVXGA1-001) | NCT04954287 | Phase 1 | Biological: CVXGA1 low dose Biological: CVXGA1 high dose |
| Safety And Immunogenicity Of HDT-301 Targeting A SARS-CoV-2 Variant Spike Protein | NCT05132907 | Phase 1 | Biological: HDT-301 |
| Delayed Heterologous SARS-CoV-2 Vaccine Dosing (Boost) After Receipt of EUA Vaccines | NCT04889209 | Phase 1 Phase 2 |
Biological: Ad26.COV2.S Biological: BNT162b2 Biological: mRNA-1273 Biological: mRNA-1273.211 Biological: mRNA-1273.222 Biological: SARS-CoV-2 rS/M1 |
| GLS-5310 Vaccine in Healthy Volunteers as a Booster for SARS-CoV-2 (COVID-19) | NCT05182567 | Phase 1 | Drug: GLS-5310 (Group 1) Drug: GLS-5310 (Group 2) Drug: GLS-5310 (Group 3) Drug: GLS-5310 (Group 4) |
| COVID-19 Variant Immunologic Landscape Trial (COVAIL Trial) | NCT05289037 | Phase 1 Phase 2 |
Drug: AS03 Biological: BNT162b2 Biological: BNT162b2 (B.1.1.529) Biological: BNT162b2 (B.1.351) Biological: BNT162b2 bivalent (wild type and Omicron BA.1) Biological: BNT162b2 bivalent (wild type and Omicron BA.4/BA.5) Biological: CoV2 preS dTM [B.1.351] Biological: CoV2 preS dTM/D614 Biological: CoV2 preS dTM/D614 + B.1.351 Biological: mRNA-1273 Biological: mRNA-1273.351 Biological: mRNA-1273.529 Biological: mRNA-1273.617.2 Other: sodium chloride, 0.9% |
| A Safety, Reactogenicity, and Immunogenicity Study of mRNA-1045 (Influenza and Respiratory Syncytial Virus [RSV]) or mRNA-1230 (Influenza, RSV, and Severe Acute Respiratory Syndrome Coronavirus 2 [SARS-CoV-2]) Vaccine in Adults 50 to 75 Years Old | NCT05585632 | Phase 1 | Biological: mRNA-1010 Biological: mRNA-1345 Biological: mRNA-1273.214 Biological: mRNA-1045 Biological: mRNA-1230 |
| Chimpanzee Adenovirus and Self-Amplifying mRNA Prime-Boost Prophylactic Vaccines Against SARS-CoV-2 in Healthy Adults | NCT04776317 | Phase 1 | Biological: ChAdV68-S Biological: ChAdV68-S-TCE Biological: SAM-LNP-S Biological: SAM-LNP-S-TCE Other: sodium chloride, 0.9% |
| A Live Recombinant Newcastle Disease Virus-vectored COVID-19 Vaccine Phase 1 Study | NCT05181709 | Phase 1 | Drug: sodium chloride Biological: NDV-HXP-S IN low dose Biological: NDV-HXP-S IM low dose Biological: NDV-HXP-S IN high dose Biological: NDV-HXP-S IM high dose |
| Safety and Immunogenicity Study of a Booster Dose of the Investigational CV0501 mRNA COVID-19 Vaccine in Adults at Least 18 Years Old | NCT05477186 | Phase 1 | Biological: CV0501 (3 μg) Biological: CV0501 (6 μg) Biological: CV0501 (12 μg) Biological: CV0501 (25 μg) Biological: CV0501 (50 μg) Biological: CV0501 (75 μg) Biological: CV0501 (100 μg) Biological: CV0501 (150 μg) Biological: CV0501 (200 μg) |
| SARS-CoV-2-Spike-Ferritin-Nanoparticle (SpFN) Vaccine With ALFQ Adjuvant for Prevention of COVID-19 in Healthy Adults | NCT04784767 | Phase 1 | Biological: 25 µg SpFN_1B-06-PL + ALFQ (QS21 adjuvant) Drug: sodium chloride, USP, for injection (0.9% NaCl) Biological: 50 µg SpFN_1B-06-PL + ALFQ (QS21 adjuvant) |
| A Study of Modified mRNA Vaccines in Healthy Adults | NCT05397223 | Phase 1 | Biological: mRNA-1273 Biological: mRNA-1010 Biological: mRNA-1345 Biological: FLUAD® Biological: mRNA-1647 |
| Study of Recombinant Protein Vaccines With Adjuvant as a Primary Series and as a Booster Dose Against COVID-19 in Adults 18 Years of Age and Older (VAT00002) | NCT04762680 | Phase 2 Phase 3 |
Biological: SARS-CoV-2 recombinant protein vaccine Phase 2 Formulation 1 Biological: SARS-CoV-2 recombinant protein vaccine Phase 2 Formulation 2 Biological: SARS-CoV-2 recombinant protein vaccine Phase 2 Formulation 3 Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine, monovalent (D614)-AS03, Dosage A Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine, monovalent (B.1.351)-AS03 Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine, monovalent (D614)-AS03, Dosage B Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine, monovalent (B.1.351)-AS03 Alternative Exploratory Formulation 1 Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine, monovalent (B.1.351)-AS03 Alternative Exploratory Formulation 2 Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine, monovalent (B.1.351)-AS03 Alternative Exploratory Formulation 3 Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine, monovalent (B.1.351)-AS03 Alternative Exploratory Formulation 4 Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine, bivalent (D614 + B.1.351)-AS03 |
| Study of a Recombinant Coronavirus-Like Particle COVID-19 Vaccine in Adults | NCT04636697 | Phase 2 Phase 3 |
Drug: intramuscular injection Biological: intramuscular vaccine |
| A Ph 2 Trial With an Oral Tableted COVID-19 Vaccine | NCT05067933 | Phase 2 | Drug: VXA-CoV2-1.1-S Other: placebo tablets |
| Safety and Immunogenicity of RNA-based Vaccines Against SARS-CoV-2 Variants in Healthy Participants | NCT05004181 | Phase 2 | Biological: BNT162b2 Biological: BNT162b2 (B.1.1.7 + B.1.617.2) Biological: BNT162b2 (B.1.1.7) Biological: BNT162b2 (B.1.617.2) Biological: BNT162b2 (B.1.1.529) Other: observational |
| COVID-19 VAX Booster Dosing in Patients With Hematologic Malignancies | NCT05028374 | Phase 2 | Drug: A single “booster” dose of the Moderna mRNA COVID-19 vaccine |
| A Study to Evaluate Safety and Effectiveness of mRNA-1273 COVID-19 Vaccine in Healthy Children Between 6 Months of Age and Less Than 12 Years of Age | NCT04796896 | Phase 2 Phase 3 |
Biological: mRNA-1273 Biological: placebo Biological: mRNA-1273.214 |
| A Phase 1/2/3 Study to Evaluate the Safety, Tolerability, and Immunogenicity of an RNA Vaccine Candidate Against COVID-19 in Healthy Children | NCT04816643 | Phase 2 Phase 3 |
Biological: biological/vaccine: BNT162b2 10mcg Biological: BNT162b2 20mcg Biological: BNT162b2 30mcg Other: placebo Biological: biological/vaccine: BNT162b2 3mcg |
| A Study to Evaluate the Immunogenicity and Safety of mRNA Vaccine Boosters for SARS-CoV-2 (COVID-19) Variants | NCT04927065 | Phase 2 Phase 3 |
Biological: mRNA-1273.211 Biological: mRNA-1273 Biological: mRNA-1273.617.2 Biological: mRNA-1273.213 Biological: mRNA-1273.529 Biological: mRNA-1273.214 Biological: mRNA-1273.222 Biological: mRNA-1273.815 Biological: mRNA-1273.231 |
| Study to Evaluate Safety, Tolerability & Immunogenicity of BNT162b2 in Immunocompromised Participants ≥2 Years | NCT04895982 | Phase 2 | Biological: BNT162b2 |
| COVID-19 Booster Vaccine in Autoimmune Disease Non-Responders | NCT05000216 | Phase 2 | Biological: Moderna mRNA-1273 Biological: BNT162b2 Biological: Ad26.COV2.S Drug: continue IS (MMF or MPA) Drug: continue IS (MTX) Biological: continue IS (B cell depletion therapy) Biological: monovalent (B.1.351) CoV2 preS dTM-AS03 Drug: withhold IS (MMF or MPA) Drug: withhold IS (MTX) Drug: withhold IS (B cell depletion therapy) Biological: Moderna mRNA-1273, bivalent Biological: BNT162b2, bivalent |
| A Study to Learn About Two or More Vaccines That Are Put Together as One Shot Against Infectious Lung Illnesses, Including COVID-19 and Respiratory Syncytial Virus (RSV) | NCT05886777 | Phase 2 | Biological: combination (RSVpreF + BNTb162b2) Biological: bivalent BNT162b2 (original/Omi BA.4/BA.5) Biological: RSVpreF Biological: QIV Biological: normal saline placebo |
| Study of Monovalent and Bivalent Recombinant Protein Vaccines Against COVID-19 in Adults 18 Years of Age and Older (VAT00008) | NCT04904549 | Phase 3 | Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine (monovalent D614) (primary series) Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine (bivalent D614 + B.1.351) (primary series) Biological: placebo Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine (monovalent B.1.351) (booster dose) ≥4 months after last vaccination Biological: SARS-CoV-2 adjuvanted recombinant protein vaccine (monovalent D614) (primary series) and SARS-CoV-2 adjuvanted recombinant protein vaccine (monovalent B.1.351) (booster dose) ≥4 months after last vaccination |
| Phase 3 Study of Novavax Vaccine(s) as Booster Dose After mRNA Vaccines | NCT05875701 | Phase 3 | Biological: NVX-CoV2373 Biological: SARS-CoV-2 rS antigen/Matrix-M adjuvant |
| A Study to Evaluate Safety and Immunogenicity of mRNA-1273 Vaccine to Prevent COVID-19 in Adult Organ Transplant Recipients and in Healthy Adult Participants | NCT04860297 | Phase 3 | Biological: mRNA-1273 |
| A Study to Evaluate the Safety and Immunogenicity of the mRNA-1273.214 COVID-19 Vaccine in Healthy Children Between 6 Months to Less Than 6 Years of Age | NCT05436834 | Phase 3 | Biological: mRNA-1273.214 |
| ABNCoV2 Vaccine in Adult Subjects Previously Vaccinated for SARS-CoV-2 | NCT05329220 | Phase 3 | Biological: ABNCoV2 Biological: Comirnaty |
| A Study to Evaluate the Efficacy, Immune Response, and Safety of a COVID-19 Vaccine in Adults ≥18 Years With a Pediatric Expansion in Adolescents (12 to <18 Years) at Risk for SARS-CoV-2 | NCT04611802 | Phase 3 | Biological: SARS-CoV-2 rS/Matrix-M1 adjuvant (initial vaccination period) Other: placebo (initial vaccination period) Biological: SARS-CoV-2 rS/Matrix-M1 adjuvant (crossover vaccination period) Other: placebo (crossover vaccination period) Biological: SARS-CoV-2 rS/Matrix-M1 adjuvant (booster vaccination) Biological: SARS-CoV-2 rS/Matrix-M1 adjuvant (second booster vaccination) |
| Safety and Immunogenicity of 9-valent Human Papillomavirus (9vHPV) Vaccine Coadministered With Messenger Ribonucleic Acid (mRNA)-1273 Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) (COVID-19) Vaccine (V503-076) | NCT05119855 | Phase 3 | Biological: 9vHPV vaccine Biological: mRNA-1273 vaccine |
| Platform Trial to Compare Homologous Boost of Authorized COVID-19 Vaccines and Heterologous Boost With UB-612 Vaccine | NCT05293665 | Phase 3 | Biological: UB-612 Biological: BNT162b2 vaccine Biological: ChAdOx1-S vaccine Biological: Sinopharm BIBP |
| BCG Vaccine for Health Care Workers as Defense Against COVID 19 (BADAS) | NCT04348370 | Phase 4 | Biological: BCG vaccine Biological: placebo vaccine |
References
- World Health Organization. WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19. 2020. Available online: https://www.who.int/director-general/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020 (accessed on 5 August 2023).
- Chattopadhyay, S.; Chen, J.Y.; Chen, H.W.; Hu, C.J. Nanoparticle Vaccines Adopting Virus-like Features for Enhanced Immune Potentiation. Nanotheranostics 2017, 1, 244–260.
- Vartak, A.; Sucheck, S.J. Recent Advances in Subunit Vaccine Carriers. Vaccines 2016, 4, 12.
- Boni, M.F. Vaccination and antigenic drift in influenza. Vaccine 2008, 26 (Suppl. S3), C8–C14.
- Mehrabi, M.; Dounighi, N.M.; Rezayat, S.M.; Doroud, D.; Amani, A.; Khoobi, M.; Ajdary, S. Novel approach to improve vaccine immunogenicity: Mannosylated chitosan nanoparticles loaded with recombinant hepatitis B antigen as a targeted vaccine delivery system. J. Drug Deliv. Sci. Technol. 2018, 44, 19–26.
- Draget, K.I.; Taylor, C. Chemical, physical and biological properties of alginates and their biomedical implications. Food Hydrocoll. 2011, 25, 251–256.
- Downs, E.C.; Robertson, N.E.; Riss, T.L.; Plunkett, M.L. Calcium alginate beads as a slow-release system for delivering angiogenic molecules In Vivo and In Vitro. J. Cell. Physiol. 1992, 152, 422–429.
- Sarei, F.; Dounighi, N.M.; Zolfagharian, H.; Khaki, P.; Bidhendi, S.M. Alginate nanoparticles as a promising adjuvant and vaccine delivery system. Indian J. Pharm. Sci. 2013, 75, 442–449.
- Jiang, D.; Liang, J.; Noble, P.W. Hyaluronan as an immune regulator in human diseases. Physiol. Rev. 2011, 91, 221–264.
- Taylor, K.R.; Trowbridge, J.M.; Rudisill, J.A.; Termeer, C.C.; Simon, J.C.; Gallo, R.L. Hyaluronan fragments stimulate endothelial recognition of injury through TLR4. J. Biol. Chem. 2004, 279, 17079–17084.
- Gu, P.; Liu, Z.; Sun, Y.; Ou, N.; Hu, Y.; Liu, J.; Wu, Y.; Wang, D. Angelica sinensis polysaccharide encapsulated into PLGA nanoparticles as a vaccine delivery and adjuvant system for ovalbumin to promote immune responses. Int. J. Pharm. 2019, 554, 72–80.
- Niikura, K.; Matsunaga, T.; Suzuki, T.; Kobayashi, S.; Yamaguchi, H.; Orba, Y.; Kawaguchi, A.; Hasegawa, H.; Kajino, K.; Ninomiya, T.; et al. Gold nanoparticles as a vaccine platform: Influence of size and shape on immunological responses in vitro and in vivo. ACS Nano 2013, 7, 3926–3938.
- Wang, T.; Zou, M.; Jiang, H.; Ji, Z.; Gao, P.; Cheng, G. Synthesis of a novel kind of carbon nanoparticle with large mesopores and macropores and its application as an oral vaccine adjuvant. Eur. J. Pharm. Sci. 2011, 44, 653–659.
- An, M.; Li, M.; Xi, J.; Liu, H. Silica Nanoparticle as a Lymph Node Targeting Platform for Vaccine Delivery. ACS Appl. Mater. Interfaces 2017, 9, 23466–23475.
- Popat, A.; Hartono, S.; Stahr, F.; Liu, J.; Qiao, S.; Lu, M. Mesoporous Silica Nanoparticles for Bioadsorption, Enzyme Immobilisation, and Delivery Carriers. Nanoscale 2011, 3, 2801–2818.
- Huang, X.; Cavalcante, D.P.; Townley, H.E. Macrophage-like THP-1 cells show effective uptake of silica nanoparticles carrying inactivated diphtheria toxoid for vaccination. J. Nanoparticle Res. 2020, 22, 23.
- Thurmond, K.B.; Remsen, E.E.; Kowalewski, T.; Wooley, K.L. Packaging of DNA by shell crosslinked nanoparticles. Nucleic Acids Res. 1999, 27, 2966–2971.
- Wang, T.; Jiang, H.; Zhao, Q.; Wang, S.; Zou, M.; Cheng, G. Enhanced mucosal and systemic immune responses obtained by porous silica nanoparticles used as an oral vaccine adjuvant: Effect of silica architecture on immunological properties. Int. J. Pharm. 2012, 436, 351–358.
- Jiménez-Periáñez, A.; Abos Gracia, B.; López Relaño, J.; Diez-Rivero, C.M.; Reche, P.A.; Martínez-Naves, E.; Matveyeva, E.; Gómez del Moral, M. Mesoporous silicon microparticles enhance MHC class I cross-antigen presentation by human dendritic cells. Clin. Dev. Immunol. 2013, 2013, 362163.
- Sun, B.; Ji, Z.; Liao, Y.P.; Wang, M.; Wang, X.; Dong, J.; Chang, C.H.; Li, R.; Zhang, H.; Nel, A.E.; et al. Engineering an effective immune adjuvant by designed control of shape and crystallinity of aluminum oxyhydroxide nanoparticles. ACS Nano 2013, 7, 10834–10849.
- Allison, A.C.; Gregoriadis, G. Liposomes as immunological adjuvants. Nature 1974, 252, 252.
- Oussoren, C.; Zuidema, J.; Crommelin, D.J.A.; Storm, G. Lymphatic uptake and biodistribution of liposomes after subcutaneous injection.: II. Influence of liposomal size, lipid composition and lipid dose. Biochim. Et Biophys. Acta (BBA)—Biomembr. 1997, 1328, 261–272.
- Henriksen-Lacey, M.; Bramwell, V.W.; Christensen, D.; Agger, E.M.; Andersen, P.; Perrie, Y. Liposomes based on dimethyldioctadecylammonium promote a depot effect and enhance immunogenicity of soluble antigen. J. Control. Release 2010, 142, 180–186.
- Henriksen-Lacey, M.; Christensen, D.; Bramwell, V.W.; Lindenstrøm, T.; Agger, E.M.; Andersen, P.; Perrie, Y. Comparison of the depot effect and immunogenicity of liposomes based on dimethyldioctadecylammonium (DDA), 3β- cholesterol (DC-Chol), and 1,2-Dioleoyl-3-trimethylammonium propane (DOTAP): Prolonged liposome retention mediates stronger Th1 responses. Mol. Pharm. 2011, 8, 153–161.
- Kaur, R.; Bramwell, V.W.; Kirby, D.J.; Perrie, Y. Pegylation of DDA:TDB liposomal adjuvants reduces the vaccine depot effect and alters the Th1/Th2 immune responses. J. Control. Release 2012, 158, 72–77.
- Badiee, A.; Khamesipour, A.; Samiei, A.; Soroush, D.; Shargh, V.H.; Kheiri, M.T.; Barkhordari, F.; Robert Mc Master, W.; Mahboudi, F.; Jaafari, M.R. The role of liposome size on the type of immune response induced in BALB/c mice against leishmaniasis: Rgp63 as a model antigen. Exp. Parasitol. 2012, 132, 403–409.
- Moon, J.J.; Suh, H.; Bershteyn, A.; Stephan, M.T.; Liu, H.; Huang, B.; Sohail, M.; Luo, S.; Um, S.H.; Khant, H.; et al. Interbilayer-crosslinked multilamellar vesicles as synthetic vaccines for potent humoral and cellular immune responses. Nat. Mater. 2011, 10, 243–251.
- Spang, A.; Martijn, J.; Saw, J.H.; Lind, A.E.; Guy, L.; Ettema, T.J. Close encounters of the third domain: The emerging genomic view of archaeal diversity and evolution. Archaea 2013, 2013, 202358.
- Patel, G.B.; Zhou, H.; KuoLee, R.; Chen, W. Archaeosomes as adjuvants for combination vaccines. J. Liposome Res. 2004, 14, 191–202.
- Butts, C.; Murray, N.; Maksymiuk, A.; Goss, G.; Marshall, E.; Soulières, D.; Cormier, Y.; Ellis, P.; Price, A.; Sawhney, R.; et al. Randomized phase IIB trial of BLP25 liposome vaccine in stage IIIB and IV non-small-cell lung cancer. J. Clin. Oncol. 2005, 23, 6674–6681.
- Morein, B.; Sundquist, B.; Höglund, S.; Dalsgaard, K.; Osterhaus, A. Iscom, a novel structure for antigenic presentation of membrane proteins from enveloped viruses. Nature 1984, 308, 457–460.
- Morein, B.; Lövgren, K.; Rönnberg, B.; Sjölander, A.; Villacrés-Eriksson, M. Immunostimulating Complexes. Clin. Immunother. 1995, 3, 461–475.
- Trudel, M.; Nadon, F.; Séguin, C.; Simard, C.; Lussier, G. Experimental polyvalent ISCOMs subunit vaccine induces antibodies that neutralize human and bovine respiratory syncytial virus. Vaccine 1989, 7, 12–16.
- Pearse, M.J.; Drane, D. ISCOMATRIX adjuvant for antigen delivery. Adv. Drug Deliv. Rev. 2005, 57, 465–474.
- Sanders, M.T.; Brown, L.E.; Deliyannis, G.; Pearse, M.J. ISCOM-based vaccines: The second decade. Immunol. Cell. Biol. 2005, 83, 119–128.
- Ennis, F.A.; Cruz, J.; Jameson, J.; Klein, M.; Burt, D.; Thipphawong, J. Augmentation of human influenza A virus-specific cytotoxic T lymphocyte memory by influenza vaccine and adjuvanted carriers (ISCOMS). Virology 1999, 259, 256–261.
- Frazer, I.H.; Quinn, M.; Nicklin, J.L.; Tan, J.; Perrin, L.C.; Ng, P.; O’Connor, V.M.; White, O.; Wendt, N.; Martin, J.; et al. Phase 1 study of HPV16-specific immunotherapy with E6E7 fusion protein and ISCOMATRIX adjuvant in women with cervical intraepithelial neoplasia. Vaccine 2004, 23, 172–181.
- Cox, J.C.; Sjölander, A.; Barr, I.G. ISCOMs and other saponin based adjuvants. Adv. Drug Deliv. Rev. 1998, 32, 247–271.
- Sjölander, A.; Drane, D.; Maraskovsky, E.; Scheerlinck, J.P.; Suhrbier, A.; Tennent, J.; Pearse, M. Immune responses to ISCOM formulations in animal and primate models. Vaccine 2001, 19, 2661–2665.
- Drane, D.; Gittleson, C.; Boyle, J.; Maraskovsky, E. ISCOMATRIX adjuvant for prophylactic and therapeutic vaccines. Expert Rev. Vaccines 2007, 6, 761–772.
- Skene, C.D.; Sutton, P. Saponin-adjuvanted particulate vaccines for clinical use. Methods 2006, 40, 53–59.
- Morein, B.; Hu, K.F.; Abusugra, I. Current status and potential application of ISCOMs in veterinary medicine. Adv. Drug Deliv. Rev. 2004, 56, 1367–1382.
- Vesikari, T.; Knuf, M.; Wutzler, P.; Karvonen, A.; Kieninger-Baum, D.; Schmitt, H.J.; Baehner, F.; Borkowski, A.; Tsai, T.F.; Clemens, R. Oil-in-water emulsion adjuvant with influenza vaccine in young children. N. Engl. J. Med. 2011, 365, 1406–1416.
- Grgacic, E.V.; Anderson, D.A. Virus-like particles: Passport to immune recognition. Methods 2006, 40, 60–65.
- Huckriede, A.; Bungener, L.; Stegmann, T.; Daemen, T.; Medema, J.; Palache, A.M.; Wilschut, J. The virosome concept for influenza vaccines. Vaccine 2005, 23 (Suppl. S1), S26–S38.
- Glück, R.; Moser, C.; Metcalfe, I.C. Influenza virosomes as an efficient system for adjuvanted vaccine delivery. Expert Opin. Biol. Ther. 2004, 4, 1139–1145.
- de Bruijn, I.A.; Nauta, J.; Cramer, W.C.; Gerez, L.; Palache, A.M. Clinical experience with inactivated, virosomal influenza vaccine. Vaccine 2005, 23 (Suppl. S1), S39–S49.
- Choi, Y.; Chang, J. Viral vectors for vaccine applications. Clin. Exp. Vaccine Res. 2013, 2, 97–105.
- Schuldt, N.J.; Amalfitano, A. Malaria vaccines: Focus on adenovirus based vectors. Vaccine 2012, 30, 5191–5198.
- Simon, J.; Edelman, R. Clinical Evaluation of Adjuvants; Academic Press: Cambridge, MA, USA, 2006; pp. 319–342.
- Schijns, V.E.J.C.; O’Hagan, D.T. (Eds.) Immunopotentiators in Modern Vaccines; Academic Press: Cambridge, MA, USA, 2006.
- Villa, L.L.; Costa, R.L.; Petta, C.A.; Andrade, R.P.; Ault, K.A.; Giuliano, A.R.; Wheeler, C.M.; Koutsky, L.A.; Malm, C.; Lehtinen, M.; et al. Prophylactic quadrivalent human papillomavirus (types 6, 11, 16, and 18) L1 virus-like particle vaccine in young women: A randomised double-blind placebo-controlled multicentre phase II efficacy trial. Lancet Oncol. 2005, 6, 271–278.
- Guerrini, G.; Magrì, D.; Gioria, S.; Medaglini, D.; Calzolai, L. Characterization of nanoparticles-based vaccines for COVID-19. Nat. Nanotechnol. 2022, 17, 570–576.
- Huynh, A.; Kelton, J.G.; Arnold, D.M.; Daka, M.; Nazy, I. Antibody epitopes in vaccine-induced immune thrombotic thrombocytopaenia. Nature 2021, 596, 565–569.
- Nanomedicine and the COVID-19 vaccines. Nat. Nanotechnol. 2020, 15, 963.
- Gopalakrishnan, A.M.; Kundu, A.K.; Mandal, T.K.; Kumar, N. Novel nanosomes for gene delivery to Plasmodium falciparum-infected red blood cells. Sci. Rep. 2013, 3, 1534.
- Yamada, Y. Nucleic Acid Drugs-Current Status, Issues, and Expectations for Exosomes. Cancers 2021, 13, 5002.
- Kulkarni, J.A.; Witzigmann, D.; Thomson, S.B.; Chen, S.; Leavitt, B.R.; Cullis, P.R.; van der Meel, R. The current landscape of nucleic acid therapeutics. Nat. Nanotechnol. 2021, 16, 630–643.
- Chandra, P.K.; Kundu, A.K.; Hazari, S.; Chandra, S.; Bao, L.; Ooms, T.; Morris, G.F.; Wu, T.; Mandal, T.K.; Dash, S. Inhibition of hepatitis C virus replication by intracellular delivery of multiple siRNAs by nanosomes. Mol. Ther. 2012, 20, 1724–1736.
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the clinic: An update. Bioeng. Transl. Med. 2019, 4, e10143.
- Akinc, A.; Maier, M.A.; Manoharan, M.; Fitzgerald, K.; Jayaraman, M.; Barros, S.; Ansell, S.; Du, X.; Hope, M.J.; Madden, T.D.; et al. The Onpattro story and the clinical translation of nanomedicines containing nucleic acid-based drugs. Nat. Nanotechnol. 2019, 14, 1084–1087.
- Mendes, B.B.; Conniot, J.; Avital, A.; Yao, D.; Jiang, X.; Zhou, X.; Sharf-Pauker, N.; Xiao, Y.; Adir, O.; Liang, H.; et al. Nanodelivery of nucleic acids. Nat. Rev. Methods Primers 2022, 2, 24.
- Yang, K.; Wang, C.; White, K.I.; Pfuetzner, R.A.; Esquivies, L.; Brunger, A.T. Structural conservation among variants of the SARS-CoV-2 spike postfusion bundle. Proc. Natl. Acad. Sci. USA 2022, 119, e2119467119.
- Kandwal, S.; Fayne, D. Genetic conservation across SARS-CoV-2 non-structural proteins—Insights into possible targets for treatment of future viral outbreaks. Virology 2023, 581, 97–115.
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. mRNA vaccines for infectious diseases: Principles, delivery and clinical translation. Nat. Rev. Drug Discov. 2021, 20, 817–838.
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA Modifications in Gene Expression Regulation. Cell 2017, 169, 1187–1200.
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279.
- Chen, S.; Evert, B.; Adeniyi, A.; Salla-Martret, M.; Lua, L.H.; Ozberk, V.; Pandey, M.; Good, M.F.; Suhrbier, A.; Halfmann, P.; et al. Ambient Temperature Stable, Scalable COVID-19 Polymer Particle Vaccines Induce Protective Immunity. Adv. Healthc. Mater. 2022, 11, e2102089.
- Chavda, V.P.; Bezbaruah, R.; Valu, D.; Patel, B.; Kumar, A.; Prasad, S.; Kakoti, B.B.; Kaushik, A.; Jesawadawala, M. Adenoviral Vector-Based Vaccine Platform for COVID-19: Current Status. Vaccines 2023, 11, 432.
- Fausther-Bovendo, H.; Kobinger, G.P. Pre-existing immunity against Ad vectors: Humoral, cellular, and innate response, what’s important? Hum. Vaccines Immunother. 2014, 10, 2875–2884.
- Yin, Q.; Luo, W.; Mallajosyula, V.; Bo, Y.; Guo, J.; Xie, J.; Sun, M.; Verma, R.; Li, C.; Constantz, C.M.; et al. A TLR7-nanoparticle adjuvant promotes a broad immune response against heterologous strains of influenza and SARS-CoV-2. Nat. Mater. 2023, 22, 380–390.
- Yang, M.C.; Wang, C.C.; Tang, W.C.; Chen, K.M.; Chen, C.Y.; Lin, H.H.; Hsieh, Y.C.; Wang, N.H.; Kuo, Y.C.; Chu, P.T.; et al. Immunogenicity of a spike protein subunit-based COVID-19 vaccine with broad protection against various SARS-CoV-2 variants in animal studies. PLoS ONE 2023, 18, e0283473.
- Peek, L.J.; Middaugh, C.R.; Berkland, C. Nanotechnology in vaccine delivery. Adv. Drug Deliv. Rev. 2008, 60, 915–928.
- Shin, M.D.; Shukla, S.; Chung, Y.H.; Beiss, V.; Chan, S.K.; Ortega-Rivera, O.A.; Wirth, D.M.; Chen, A.; Sack, M.; Pokorski, J.K.; et al. COVID-19 vaccine development and a potential nanomaterial path forward. Nat. Nanotechnol. 2020, 15, 646–655.
- Liang, W.; Dong, Y.; Shao, R.; Zhang, S.; Wu, X.; Huang, X.; Sun, B.; Zeng, B.; Zhao, J. Application of nanoparticles in drug delivery for the treatment of osteosarcoma: Focussing on the liposomes. J. Drug Target 2022, 30, 463–475.
- Shimon, M.B.; Shapira, S.; Seni, J.; Arber, N. The Big Potential of Small Particles: Lipid-Based Nanoparticles and Exosomes in Vaccination. Vaccines 2022, 10, 1119.
- García-Pinel, B.; Porras-Alcalá, C.; Ortega-Rodríguez, A.; Sarabia, F.; Prados, J.; Melguizo, C.; López-Romero, J.M. Lipid-Based Nanoparticles: Application and Recent Advances in Cancer Treatment. Nanomaterials 2019, 9, 638.
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid nanoparticles for mRNA delivery. Nat. Rev. Mater. 2021, 6, 1078–1094.
- Panahi, Y.; Einollahi, B.; Beiraghdar, F.; Darvishi, M.; Fathi, S.; Javanbakht, M.; Shafiee, S.; Akhavan-Sigari, R. Fully understanding the efficacy profile of the COVID-19 vaccination and its associated factors in multiple real-world settings. Front. Immunol. 2022, 13, 947602.
- Demento, S.L.; Cui, W.; Criscione, J.M.; Stern, E.; Tulipan, J.; Kaech, S.M.; Fahmy, T.M. Role of sustained antigen release from nanoparticle vaccines in shaping the T cell memory phenotype. Biomaterials 2012, 33, 4957–4964.
- Thomas, C.; Rawat, A.; Hope-Weeks, L.; Ahsan, F. Aerosolized PLA and PLGA nanoparticles enhance humoral, mucosal and cytokine responses to hepatitis B vaccine. Mol. Pharm. 2011, 8, 405–415.
- Bershteyn, A.; Hanson, M.C.; Crespo, M.P.; Moon, J.J.; Li, A.V.; Suh, H.; Irvine, D.J. Robust IgG responses to nanograms of antigen using a biomimetic lipid-coated particle vaccine. J. Control. Release 2012, 157, 354–365.
- Moon, J.J.; Suh, H.; Polhemus, M.E.; Ockenhouse, C.F.; Yadava, A.; Irvine, D.J. Antigen-displaying lipid-enveloped PLGA nanoparticles as delivery agents for a Plasmodium vivax malaria vaccine. PLoS ONE 2012, 7, e31472.
- Mann, J.F.; McKay, P.F.; Arokiasamy, S.; Patel, R.K.; Klein, K.; Shattock, R.J. Pulmonary delivery of DNA vaccine constructs using deacylated PEI elicits immune responses and protects against viral challenge infection. J. Control. Release 2013, 170, 452–459.
- Saini, V.; Jain, V.; Sudheesh, M.S.; Dixit, S.; Gaur, R.L.; Sahoo, M.K.; Joseph, S.K.; Verma, S.K.; Jaganathan, K.S.; Murthy, P.K.; et al. Humoral and cell-mediated immune-responses after administration of a single-shot recombinant hepatitis B surface antigen vaccine formulated with cationic poly(l-lactide) microspheres. J. Drug Target. 2010, 18, 212–222.
- Gupta, P.N.; Vyas, S.P. Investigation of lectinized liposomes as M-cell targeted carrier-adjuvant for mucosal immunization. Colloids Surf. B Biointerfaces 2011, 82, 118–125.
- Ulery, B.D.; Kumar, D.; Ramer-Tait, A.E.; Metzger, D.W.; Wannemuehler, M.J.; Narasimhan, B. Design of a protective single-dose intranasal nanoparticle-based vaccine platform for respiratory infectious diseases. PLoS ONE 2011, 6, e17642.
- Lugade, A.A.; Bharali, D.J.; Pradhan, V.; Elkin, G.; Mousa, S.A.; Thanavala, Y. Single low-dose un-adjuvanted HBsAg nanoparticle vaccine elicits robust, durable immunity. Nanomedicine 2013, 9, 923–934.
- Nochi, T.; Yuki, Y.; Takahashi, H.; Sawada, S.; Mejima, M.; Kohda, T.; Harada, N.; Kong, I.G.; Sato, A.; Kataoka, N.; et al. Nanogel antigenic protein-delivery system for adjuvant-free intranasal vaccines. Nat. Mater. 2010, 9, 572–578.
- Slütter, B.; Bal, S.; Keijzer, C.; Mallants, R.; Hagenaars, N.; Que, I.; Kaijzel, E.; van Eden, W.; Augustijns, P.; Löwik, C.; et al. Nasal vaccination with N-trimethyl chitosan and PLGA based nanoparticles: Nanoparticle characteristics determine quality and strength of the antibody response in mice against the encapsulated antigen. Vaccine 2010, 28, 6282–6291.
- Moon, S.H.; Shin, E.C.; Noh, Y.W.; Lim, Y.T. Evaluation of hyaluronic acid-based combination adjuvant containing monophosphoryl lipid A and aluminum salt for hepatitis B vaccine. Vaccine 2015, 33, 4762–4769.
- Sanchez-Villamil, J.I.; Tapia, D.; Torres, A.G. Development of a Gold Nanoparticle Vaccine against Enterohemorrhagic Escherichia coli O157:H7. mBio 2019, 10, e01869-19.
- Schreiber, H.A.; Prechl, J.; Jiang, H.; Zozulya, A.; Fabry, Z.; Denes, F.; Sandor, M. Using carbon magnetic nanoparticles to target, track, and manipulate dendritic cells. J. Immunol. Methods 2010, 356, 47–59.
- Montalvo-Quirós, S.; Vallet-Regí, M.; Palacios, A.; Anguita, J.; Prados-Rosales, R.C.; González, B.; Luque-Garcia, J.L. Mesoporous Silica Nanoparticles as a Potential Platform for Vaccine Development against Tuberculosis. Pharmaceutics 2020, 12, 1218.
- Neuhaus, V.; Chichester, J.A.; Ebensen, T.; Schwarz, K.; Hartman, C.E.; Shoji, Y.; Guzmán, C.A.; Yusibov, V.; Sewald, K.; Braun, A. A new adjuvanted nanoparticle-based H1N1 influenza vaccine induced antigen-specific local mucosal and systemic immune responses after administration into the lung. Vaccine 2014, 32, 3216–3222.
- Knudsen, N.P.; Olsen, A.; Buonsanti, C.; Follmann, F.; Zhang, Y.; Coler, R.N.; Fox, C.B.; Meinke, A.; D’Oro, U.; Casini, D.; et al. Different human vaccine adjuvants promote distinct antigen-independent immunological signatures tailored to different pathogens. Sci. Rep. 2016, 6, 19570.
- Morçöl, T.; Hurst, B.L.; Tarbet, E.B. Calcium phosphate nanoparticle (CaPNP) for dose-sparing of inactivated whole virus pandemic influenza A (H1N1) 2009 vaccine in mice. Vaccine 2017, 35, 4569–4577.
- Mašek, J.; Bartheldyová, E.; Turánek-Knotigová, P.; Skrabalová, M.; Korvasová, Z.; Plocková, J.; Koudelka, S.; Skodová, P.; Kulich, P.; Křupka, M.; et al. Metallochelating liposomes with associated lipophilised norAbuMDP as biocompatible platform for construction of vaccines with recombinant His-tagged antigens: Preparation, structural study and immune response towards rHsp90. J. Control. Release 2011, 151, 193–201.
- Milicic, A.; Kaur, R.; Reyes-Sandoval, A.; Tang, C.K.; Honeycutt, J.; Perrie, Y.; Hill, A.V. Small cationic DDA:TDB liposomes as protein vaccine adjuvants obviate the need for TLR agonists in inducing cellular and humoral responses. PLoS ONE 2012, 7, e34255.
- McNeil, S.E.; Rosenkrands, I.; Agger, E.M.; Andersen, P.; Perrie, Y. Subunit vaccines: Distearoylphosphatidylcholine-based liposomes entrapping antigen offer a neutral alternative to dimethyldioctadecylammonium-based cationic liposomes as an adjuvant delivery system. J. Pharm. Sci. 2011, 100, 1856–1865.
- Rosenkrands, I.; Vingsbo-Lundberg, C.; Bundgaard, T.J.; Lindenstrøm, T.; Enouf, V.; van der Werf, S.; Andersen, P.; Agger, E.M. Enhanced humoral and cell-mediated immune responses after immunization with trivalent influenza vaccine adjuvanted with cationic liposomes. Vaccine 2011, 29, 6283–6291.
- Henriksen-Lacey, M.; Devitt, A.; Perrie, Y. The vesicle size of DDA:TDB liposomal adjuvants plays a role in the cell-mediated immune response but has no significant effect on antibody production. J. Control. Release 2011, 154, 131–137.
- Christensen, D.; Henriksen-Lacey, M.; Kamath, A.T.; Lindenstrøm, T.; Korsholm, K.S.; Christensen, J.P.; Rochat, A.F.; Lambert, P.H.; Andersen, P.; Siegrist, C.A.; et al. A cationic vaccine adjuvant based on a saturated quaternary ammonium lipid have different in vivo distribution kinetics and display a distinct CD4 T cell-inducing capacity compared to its unsaturated analog. J. Control. Release 2012, 160, 468–476.
- de Veer, M.; Neeland, M.; Burke, M.; Pleasance, J.; Nathanielsz, J.; Elhay, M.; Meeusen, E. Cell recruitment and antigen trafficking in afferent lymph after injection of antigen and poly(I:C) containing liposomes, in aqueous or oil-based formulations. Vaccine 2013, 31, 1012–1018.
- Cox, J.; Coulter, A.; Macfarlan, R.; Beezum, L.; Bates, J.; Wong, T.-Y.; Drane, D. Development of an Influenza-Iscom™ Vaccine. In Vaccine Design: The Role of Cytokine Networks; Gregoriadis, G., McCormack, B., Allison, A.C., Eds.; Springer: Boston, MA, USA, 1997; pp. 33–49.
- Brito, L.A.; Chan, M.; Baudner, B.; Gallorini, S.; Santos, G.; O’Hagan, D.T.; Singh, M. An alternative renewable source of squalene for use in emulsion adjuvants. Vaccine 2011, 29, 6262–6268.
- Calabro, S.; Tritto, E.; Pezzotti, A.; Taccone, M.; Muzzi, A.; Bertholet, S.; De Gregorio, E.; O’Hagan, D.T.; Baudner, B.; Seubert, A. The adjuvant effect of MF59 is due to the oil-in-water emulsion formulation, none of the individual components induce a comparable adjuvant effect. Vaccine 2013, 31, 3363–3369.
- Singh, M.; Kazzaz, J.; Ugozzoli, M.; Baudner, B.; Pizza, M.; Giuliani, M.; Hawkins, L.D.; Otten, G.; O’Hagan, D.T. MF59 oil-in-water emulsion in combination with a synthetic TLR4 agonist (E6020) is a potent adjuvant for a combination Meningococcus vaccine. Hum. Vaccines Immunother. 2012, 8, 486–490.
- Myc, A.; Kukowska-Latallo, J.F.; Smith, D.M.; Passmore, C.; Pham, T.; Wong, P.; Bielinska, A.U.; Baker, J.R., Jr. Nanoemulsion nasal adjuvant W₈₀5EC induces dendritic cell engulfment of antigen-primed epithelial cells. Vaccine 2013, 31, 1072–1079.
- Makidon, P.E.; Nigavekar, S.S.; Bielinska, A.U.; Mank, N.; Shetty, A.M.; Suman, J.; Knowlton, J.; Myc, A.; Rook, T.; Baker, J.R., Jr. Characterization of stability and nasal delivery systems for immunization with nanoemulsion-based vaccines. J. Aerosol Med. Pulm. Drug Deliv. 2010, 23, 77–89.
- Lousada-Dietrich, S.; Jogdand, P.S.; Jepsen, S.; Pinto, V.V.; Ditlev, S.B.; Christiansen, M.; Larsen, S.O.; Fox, C.B.; Raman, V.S.; Howard, R.F.; et al. A synthetic TLR4 agonist formulated in an emulsion enhances humoral and Type 1 cellular immune responses against GMZ2—A GLURP-MSP3 fusion protein malaria vaccine candidate. Vaccine 2011, 29, 3284–3292.
- Treanor, J.J.; Essink, B.; Hull, S.; Reed, S.; Izikson, R.; Patriarca, P.; Goldenthal, K.L.; Kohberger, R.; Dunkle, L.M. Evaluation of safety and immunogenicity of recombinant influenza hemagglutinin (H5/Indonesia/05/2005) formulated with and without a stable oil-in-water emulsion containing glucopyranosyl-lipid A (SE+GLA) adjuvant. Vaccine 2013, 31, 5760–5765.
- Lumsden, J.M.; Nurmukhambetova, S.; Klein, J.H.; Sattabongkot, J.; Bennett, J.W.; Bertholet, S.; Fox, C.B.; Reed, S.G.; Ockenhouse, C.F.; Howard, R.F.; et al. Evaluation of immune responses to a Plasmodium vivax CSP-based recombinant protein vaccine candidate in combination with second-generation adjuvants in mice. Vaccine 2012, 30, 3311–3319.
- Bovier, P.A. Epaxal: A virosomal vaccine to prevent hepatitis A infection. Expert Rev. Vaccines 2008, 7, 1141–1150.
- Herzog, C.; Hartmann, K.; Künzi, V.; Kürsteiner, O.; Mischler, R.; Lazar, H.; Glück, R. Eleven years of Inflexal V-a virosomal adjuvanted influenza vaccine. Vaccine 2009, 27, 4381–4387.
- Glück, R. Intranasal immunization against influenza. J. Aerosol Med. 2002, 15, 221–228.
- Mutsch, M.; Zhou, W.; Rhodes, P.; Bopp, M.; Chen, R.T.; Linder, T.; Spyr, C.; Steffen, R. Use of the inactivated intranasal influenza vaccine and the risk of Bell’s palsy in Switzerland. N. Engl. J. Med. 2004, 350, 896–903.
- de Bruijn, I.A.; Nauta, J.; Gerez, L.; Palache, A.M. The virosomal influenza vaccine Invivac®: Immunogenicity and tolerability compared to an adjuvanted influenza vaccine (Fluad®) in elderly subjects. Vaccine 2006, 24, 6629–6631.
- de Bruijn, I.; Meyer, I.; Gerez, L.; Nauta, J.; Giezeman, K.; Palache, B. Antibody induction by virosomal, MF59-adjuvanted, or conventional influenza vaccines in the elderly. Vaccine 2007, 26, 119–127.
- Jain, H.; Kumavat, V.; Singh, T.; Versteilen, A.; Sarnecki, M. Immunogenicity and safety of a pediatric dose of a virosomal hepatitis A vaccine in healthy children in India. Hum. Vaccines Immunother. 2014, 10, 2089–2097.

