 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Sujeenthar Tharmalingam | -- | 4202 | 2023-12-16 07:17:03 | | | |
| 2 | Lindsay Dong | Meta information modification | 4202 | 2023-12-18 06:58:25 | | |
Video Upload Options
Candidiasis is a highly pervasive infection posing major health risks, especially for immunocompromised populations. Pathogenic Candida species have evolved intrinsic and acquired resistance to a variety of antifungal medications.
1. Introduction
1.1. Candidiasis
1.1.1. Candida Species of Interest
1.1.2. Candida auris
1.2. Primary/Intrinsic Resistance vs. Secondary/Acquired Resistance
1.3. Standardized Measures of Susceptibility Testing
2. Antifungal Classes and Frequency of Resistance
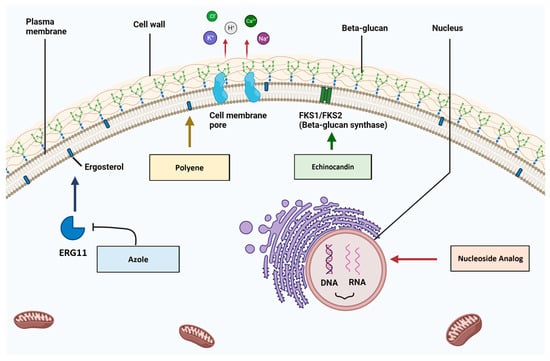
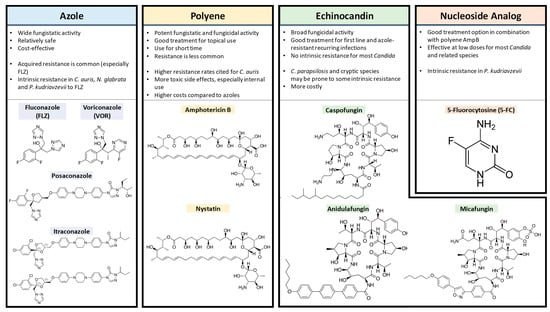
2.1. Azoles
2.2. Polyenes
2.3. Echinocandins
2.4. 5FC
3. The Ergosterol Biosynthesis Pathway and Antifungal Resistance
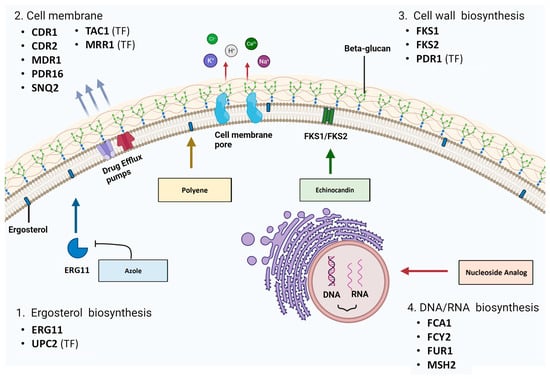
3.1. ERG11
3.2. Mutations in Transcriptional Regulators
3.3. Other ERG Genes and Toxic Diol Formation
4. Cell Membrane Proteins and Antifungal Resistance
4.1. Drug Efflux Pump/Transporter Genes and Resistant Mutations
4.2. Transcriptional Regulators of Transporter Genes
4.3. Post-Translational Regulation of Transporter Genes
5. The Cell Wall Biosynthesis Pathway and Antifungal Resistance
5.1. FKS1 and FKS2 Sequence Mutations
5.2. Transcriptional Regulators of fks Genes
5.3. Protein Analysis Associated with Echinocandin Resistance
6. The Nucleic Acid Biosynthesis Pathway and Antifungal Resistance
The biosynthesis of nucleic acids (DNA and RNA) and subsequent protein synthesis in pathogenic fungi can be targeted with nucleoside analogue 5-fluorocytosine (5-FC). As a prodrug, it requires activation within the fungal cell via metabolism by the pyrimidine salvage pathway [65]. Then, it is incorporated as a toxic substrate, and the affected nucleotides have damaging effects on cell viability [65]. Membrane permeases encoded by FCY2 (cytosine permease) and other homologs (FCY21 and FCY22) are responsible for the active transport of 5-FC into the cell (Figure 3) [65]. 5-FC is then converted to toxic 5-fluoro-uridylate by enzymes encoded by fcy1 (cytosine deaminase) and FUR1 (uracil phosphoribosyltransferase (UPRT)) [65]. The FCY1 homologue in C. albicans and other Candida species is the FCA1 gene [136][137]. The lack of cytosine deaminase in mammalian cells prevents 5-FC conversion and subsequent toxic effects [138].
Resistance to 5-FC could arise with mutation or loss of any of the three key enzymes (FCY1, FCY2 or FUR1), as discovered in model organism yeast Saccharomyces cerevisiae [139][140]. Increased pyrimidine production in the fungal cell can also serve to circumvent toxic antifungal activity [64][65]. Kern et al. (1991) were among the first to identify the correlation between a point mutation (Arg134Ser) in the FUR1 gene and 5-FC resistance in S. cerevisiae yeast cells [141].
7. Biofilm Formation and Antifungal Resistance
7.1. Biofilm Formation during Antifungal Treatment
7.2. The Roles of β-1,3 Glucan and Biofilm-Associated Antifungal Resistance
7.3. Relevant Antifungal Resistance Genes in Biofilm-Associated Candida Infections
8. Summary
With antifungal resistance being a continued problem, there is a need for the development of quick and reliable molecular diagnostic tests that detect organisms with intrinsic and/or secondary resistance due to the genetic mechanisms [29]. Additionally, faster methods of species identification would be useful, given the differences in frequency and impact of antifungal resistance among Candida species. Currently, PCR-based methods using fungal cultures are still the first option for species identification and detection of antifungal resistance in individual strains [124][164]. The usefulness of real-time testing for resistance in Candida species to modify the treatment course has been well documented.
Resistance can be acquired through a dynamic combination of numerous point mutations and other genetic or transcriptional alterations. A large-scale comparison between matched fluconazole-resistant and -susceptible C. albicans clinical isolates using microarray analysis identified almost two hundred genes (n = 198) that were differentially expressed [165]. In resistant isolates, multidrug resistance and oxidative stress response genes, among others, were found to be upregulated compared to susceptible samples [165]. This highlights the fact that a dynamic response that leads to antifungal resistance and identification of reliable biomarkers or gene expression profiles that different strains have in common would be beneficial to improve future treatment decisions.
References
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and Mechanisms of Antifungal Resistance. Antibiotics 2020, 9, 312.
- Vázquez-González, D.; Perusquía-Ortiz, A.M.; Hundeiker, M.; Bonifaz, A. Opportunistic Yeast Infections: Candidiasis, Cryptococcosis, Trichosporonosis and Geotrichosis. JDDG J. Ger. Soc. Dermatol. 2013, 11, 381–394.
- Fisher, M.C.; Gow, N.A.R.; Gurr, S.J. Tackling Emerging Fungal Threats to Animal Health, Food Security and Ecosystem Resilience. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20160332.
- Kourkoumpetis, T.K.; Velmahos, G.C.; Ziakas, P.D.; Tampakakis, E.; Manolakaki, D.; Coleman, J.J.; Mylonakis, E. The Effect of Cumulative Length of Hospital Stay on the Antifungal Resistance of Candida Strains Isolated from Critically Ill Surgical Patients. Mycopathologia 2011, 171, 85–91.
- Dąbrowska, M.; Sienkiewicz, M.; Kwiatkowski, P.; Dąbrowski, M. Diagnosis and Treatment of Mucosa Candida spp. Infections—A Review Article. Ann. Univ. Mariae Curie Sklodowska Sect. C Biol. 2019, 73, 61–68.
- Gonsalves, W.C.; Wrightson, A.S.; Henry, R.G. Common Oral Conditions in Older Persons. Am. Fam. Physician 2008, 78, 845–852.
- Lalla, R.V.; Latortue, M.C.; Hong, C.H.; Ariyawardana, A.; D’Amato-Palumbo, S.; Fischer, D.J.; Martof, A.; Nicolatou-Galitis, O.; Patton, L.L.; Elting, L.S.; et al. A Systematic Review of Oral Fungal Infections in Patients Receiving Cancer Therapy. Support. Care Cancer 2010, 18, 985–992.
- Rohr, Y.; Adams, J.; Young, L. Oral Discomfort in Palliative Care: Results of an Exploratory Study of the Experiences of Terminally Ill Patients. Int. J. Palliat. Nurs. 2010, 16, 439–444.
- Pfaller, M.; Neofytos, D.; Diekema, D.; Azie, N.; Meier-Kriesche, H.U.; Quan, S.P.; Horn, D. Epidemiology and Outcomes of Candidemia in 3648 Patients: Data from the Prospective Antifungal Therapy (PATH Alliance®) Registry, 2004–2008. Diagn. Microbiol. Infect. Dis. 2012, 74, 323–331.
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the Emerging Threat of Antifungal Resistance to Human Health. Nat. Rev. Microbiol. 2022, 20, 557–571.
- Kumar, R.; Srivastava, V. Application of Anti-Fungal Vaccines as a Tool against Emerging Anti-Fungal Resistance. Front. Fungal Biol. 2023, 4, 1241539.
- Benedict, K.; Jackson, B.R.; Chiller, T.; Beer, K.D. Estimation of Direct Healthcare Costs of Fungal Diseases in the United States. Clin. Infect. Dis. 2019, 68, 1791–1797.
- Gabaldón, T.; Naranjo-Ortíz, M.A.; Marcet-Houben, M. Evolutionary Genomics of Yeast Pathogens in the Saccharomycotina. FEMS Yeast Res. 2016, 16, fow064.
- Defosse, T.A.; Le Govic, Y.; Courdavault, V.; Clastre, M.; Vandeputte, P.; Chabasse, D.; Bouchara, J.P.; Giglioli-Guivarc’h, N.; Papon, N. Yeasts from the CTG Clade (Candida Clade): Biology, Impact in Human Health, and Biotechnological Applications. J. Mycol. Med. 2018, 28, 257–268.
- Santos, M.A.S.; Gomes, A.C.; Santos, M.C.; Carreto, L.C.; Moura, G.R. The Genetic Code of the Fungal CTG Clade. C R. Biol. 2011, 334, 607–611.
- Borman, A.M.; Johnson, E.M. Name Changes for Fungi of Medical Importance, 2018 to 2019. J. Clin. Microbiol. 2021, 59.
- Fidel, P.L.; Vazquez, J.A.; Sobel, J.D. Candida Glabrata: Review of Epidemiology, Pathogenesis, and Clinical Disease with Comparison to C. Albicans. Clin. Microbiol. Rev. 1999, 12, 80–96.
- Healey, K.R.; Ortigosa, C.J.; Shor, E.; Perlin, D.S. Genetic Drivers of Multidrug Resistance in Candida Glabrata. Front. Microbiol. 2016, 7, 1995.
- World Health Organization. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; World Health Organization: Geneva, Switzerland, 2022; Volume 1.
- CDC Antibiotic Resistance Threats in the United States; U.S. Department of Health and Human Services: Atlanta, GA, USA, 2019.
- Brandt, M.E.; Lockhart, S.R. Recent Taxonomic Developments with Candida and Other Opportunistic Yeasts. Curr. Fungal Infect. Rep. 2012, 6, 170–177.
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Ellis, D.; Tullio, V.; Rodloff, A.; Fu, W.; Ling, T.A. Results from the Artemis Disk Global Antifungal Surveillance Study, 1997 to 2007: A 10.5-Year Analysis of Susceptibilities of Candida Species to Fluconazole and Voriconazole as Determined by CLSI Standardized Disk Diffusion. J. Clin. Microbiol. 2010, 48, 1366–1377.
- Walsh, T.J.; Groll, A.; Hiemenz, J.; Fleming, R.; Roilides, E.; Anaissie, E. Infections Due to Emerging and Uncommon Medically Important Fungal Pathogens. Clin. Microbiol. Infect. 2004, 10, 48–66.
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida Auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140.
- Infection Prevention and Control. Candida auris. Public Health Ontario. Available online: https://www.publichealthontario.ca/en/Diseases-and-Conditions/Health-Care-Associated-Infections/Candida-auris (accessed on 10 October 2023).
- Centers for Disease Control and Prevention. Antifungal Susceptibility Testing and Interpretation. Available online: https://www.cdc.gov/fungal/candida-auris/c-auris-antifungal.html#print (accessed on 10 October 2023).
- CLSI. CLSI Performance Standards for Antifungal Susceptibility Testing of Yeasts, 3rd ed.; CLSI Supplement M27M44S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022; Volume 40.
- Wiederhold, N.P. Antifungal Susceptibility Testing: A Primer for Clinicians. Open Forum Infect. Dis. 2021, 8, ofab444.
- Garcia-Effron, G. Molecular Markers of Antifungal Resistance: Potential Uses in Routine Practice and Future Perspectives. J. Fungi 2021, 7, 197.
- Castanheira, M.; Deshpande, L.M.; Davis, A.P.; Carvalhaes, C.G.; Pfaller, M.A. Azole Resistance in Candida Glabrata Clinical Isolates from Global Surveillance Is Associated with Efflux Overexpression. J. Glob. Antimicrob. Resist. 2022, 29, 371–377.
- White, T.C.; Holleman, S.; Dy, F.; Mirels, L.F.; Stevens, D.A. Resistance Mechanisms in Clinical Isolates of Candida Albicans. Antimicrob. Agents Chemother. 2002, 46, 1704–1713.
- Shields, R.K.; Nguyen, M.H.; Press, E.G.; Kwa, A.L.; Cheng, S.; Du, C.; Clancy, C.J. The Presence of an FKS Mutation Rather than MIC Is an Independent Risk Factor for Failure of Echinocandin Therapy among Patients with Invasive Candidiasis Due to Candida Glabrata. Antimicrob. Agents Chemother. 2012, 56, 4862–4869.
- Bienvenu, A.L.; Leboucher, G.; Picot, S. Comparison of Fks Gene Mutations and Minimum Inhibitory Concentrations for the Detection of Candida Glabrata Resistance to Micafungin: A Systematic Review and Meta-Analysis. Mycoses 2019, 62, 835–846.
- Pfaller, M.A.; Andes, D.; Diekema, D.J.; Espinel-Ingroff, A.; Sheehan, D. Wild-Type MIC Distributions, Epidemiological Cutoff Values and Species-Specific Clinical Breakpoints for Fluconazole and Candida: Time for Harmonization of CLSI and EUCAST Broth Microdilution Methods. Drug Resist. Updates 2010, 13, 180–195.
- Pfaller, M.A.; Andes, D.; Arendrup, M.C.; Diekema, D.J.; Espinel-Ingroff, A.; Alexander, B.D.; Brown, S.D.; Chaturvedi, V.; Fowler, C.L.; Ghannoum, M.A.; et al. Clinical Breakpoints for Voriconazole and Candida Spp. Revisited: Review of Microbiologic, Molecular, Pharmacodynamic, and Clinical Data as They Pertain to the Development of Species-Specific Interpretive Criteria. Diagn. Microbiol. Infect. Dis. 2011, 70, 330–343.
- Pfaller, M.A.; Diekema, D.J.; Andes, D.; Arendrup, M.C.; Brown, S.D.; Lockhart, S.R.; Motyl, M.; Perlin, D.S. Clinical Breakpoints for the Echinocandins and Candida Revisited: Integration of Molecular, Clinical, and Microbiological Data to Arrive at Species-Specific Interpretive Criteria. Drug Resist. Updates 2011, 14, 164–176.
- EUCAST. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 13.0. 2023. Available online: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_5.0_Breakpoint_Table_01.pdf (accessed on 10 October 2023).
- Maria, S.; Barnwal, G.; Kumar, A.; Mohan, K.; Vinod, V.; Varghese, A.; Biswas, R. Species Distribution and Antifungal Susceptibility among Clinical Isolates of Candida Parapsilosis Complex from India. Rev. Iberoam. Micol. 2018, 35, 147–150.
- Borman, A.M.; Muller, J.; Walsh-Quantick, J.; Szekely, A.; Patterson, Z.; Palmer, M.D.; Fraser, M.; Johnson, E.M. Fluconazole Resistance in Isolates of Uncommon Pathogenic Yeast Species from the United Kingdom. Antimicrob. Agents Chemother. 2019, 63, e00211-19.
- Vigezzi, C.; Icely, P.A.; Dudiuk, C.; Rodríguez, E.; Miró, M.S.; Castillo, G.D.V.; Azcurra, A.I.; Abiega, C.; Caeiro, J.P.; Riera, F.O.; et al. Frequency, Virulence Factors and Antifungal Susceptibility of Candida Parapsilosis Species Complex Isolated from Patients with Candidemia in the Central Region of Argentina. J. Mycol. Med. 2019, 29, 285–291.
- CLSI. Epidemiological Cutoff Values for Antifungal Susceptibility Testing, 2nd ed.; CLSI Supplement M59; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020.
- Pfaller, M.A.; Diekema, D.J.; Turnidge, J.D.; Castanheira, M.; Jones, R.N. Twenty Years of the SENTRY Antifungal Surveillance Program: Results for Candida Species from 1997-2016. Open Forum Infect. Dis. 2019, 6, S79–S94.
- de Tilly, A.N.; Tharmalingam, S. Review of Treatments for Oropharyngeal Fungal Infections in HIV/AIDS Patients. Microbiol. Res. 2022, 13, 219–234.
- Shalini, K.; Kumar, N.; Drabu, S.; Sharma, P.K. Advances in Synthetic Approach to and Antifungal Activity of Triazoles. Beilstein J. Org. Chem. 2011, 7, 668–677.
- Robbins, N.; Caplan, T.; Cowen, L.E. Molecular Evolution of Antifungal Drug Resistance. Annu. Rev. Microbiol. 2017, 71, 753–775.
- Robbins, N.; Wright, G.D.; Cowen, L.E. Antifungal Drugs: The Current Armamentarium and Development of New Agents. Microbiol. Spectr. 2016, 4, 903–922.
- Sanglard, D.; Coste, A.; Ferrari, S. Antifungal Drug Resistance Mechanisms in Fungal Pathogens from the Perspective of Transcriptional Gene Regulation. FEMS Yeast Res. 2009, 9, 1029–1050.
- Azanza, J.R.; García-Quetglas, E.; Sádaba, B. Pharmacology of Azoles. Rev. Iberoam. Micol. 2007, 24, 223–227.
- Efimova, S.S.; Schagina, L.V.; Ostroumova, O.S. Investigation of Channel-Forming Activity of Polyene Macrolide Antibiotics in Planar Lipid Bilayers in the Presence of Dipole Modifiers. Acta Naturae 2014, 6, 67–79.
- Kermani, F.; Taghizadeh-Armaki, M.; Hosseini, S.A.; Amirrajab, N.; Javidnia, J.; Zaghrami, M.F.; Shokohi, T. Antifungal Resistance of Clinical Candida Albicans Isolates in Iran: A Systematic Review and Meta-Analysis. Iran. J. Public Health 2023, 52, 290–305.
- Birch, M.; Sibley, G. Antifungal Chemistry Review. In Comprehensive Medicinal Chemistry III; Elsevier: Amsterdam, The Netherlands, 2017.
- Waller, D.G.; Sampson, A.P. Chemotherapy of Infections. In Medical Pharmacology and Therapeutics; Elsevier: Amsterdam, The Netherlands, 2018.
- Espinel-Ingroff, A.; Arendrup, M.; Canton, E.; Cordob, S.; Dannaoui, E.; Garcia-Rodriguez, J.; Gonzalez, G.M.; Govender, N.P.; Martin-Mazuelos, E.; Lackner, M.; et al. Multicenter Study of Method-Dependent Epidemiological Cutoff Values for Detection of Resistance in Candida Spp. and Aspergillus Spp. to Amphotericin B and Echinocandins for the Etest Agar Diffusion Method. Antimicrob. Agents Chemother. 2017, 61, e01792-16.
- Kanafani, Z.A.; Perfect, J.R. Resistance to Antifungal Agents: Mechanisms and Clinical Impact. Clin. Infect. Dis. 2008, 46, 120–128.
- Popolo, L.; Gualtieri, T.; Ragni, E. The Yeast Cell-Wall Salvage Pathway. Med. Mycol. Suppl. 2001, 39, 111–121.
- Walker, L.A.; Gow, N.A.R.; Munro, C.A. Fungal Echinocandin Resistance. Fungal Genet. Biol. 2010, 47, 117–126.
- Douglas, C.M.; D’Ippolito, J.A.; Shei, G.J.; Meinz, M.; Onishi, J.; Marrinan, J.A.; Li, W.; Abruzzo, G.K.; Flattery, A.; Bartizal, K.; et al. Identification of the FKS1 Gene of Candida Albicans as the Essential Target of 1,3-β-D-Glucan Synthase Inhibitors. Antimicrob. Agents Chemother. 1997, 41, 2471–2479.
- Barchiesi, F.; Spreghini, E.; Tomassetti, S.; Della Vittoria, A.; Arzeni, D.; Manso, E.; Scalise, G. Effects of Caspofungin against Candida Guilliermondii and Candida Parapsilosis. Antimicrob. Agents Chemother. 2006, 50, 2719–2727.
- Cantón, E.; Pemán, J.; Sastre, M.; Romero, M.; Espinel-Ingroff, A. Killing Kinetics of Caspofungin, Micafungin, and Amphotericin B against Candida Guilliermondii. Antimicrob. Agents Chemother. 2006, 50, 2829–2832.
- Arastehfar, A.; Lass-Flörl, C.; Garcia-Rubio, R.; Daneshnia, F.; Ilkit, M.; Boekhout, T.; Gabaldon, T.; Perlin, D.S. The Quiet and Underappreciated Rise of Drug-Resistant Invasive Fungal Pathogens. J. Fungi 2020, 6, 138.
- Bohner, F.; Papp, C.; Gácser, A. The Effect of Antifungal Resistance Development on the Virulence of Candida Species. FEMS Yeast Res. 2022, 22, foac019.
- Vermes, A.; Guchelaar, H.J.; Dankert, J. Flucytosine: A Review of Its Pharmacology, Clinical Indications, Pharmacokinetics, Toxicity and Drug Interactions. J. Antimicrob. Chemother. 2000, 46, 171–179.
- Mourad, A.; Perfect, J.R. Present and Future Therapy of Cryptococcus Infections. J. Fungi 2018, 4, 79.
- Perfect, J.R.; Dismukes, W.E.; Dromer, F.; Goldman, D.L.; Graybill, J.R.; Hamill, R.J.; Harrison, T.S.; Larsen, R.A.; Lortholary, O.; Nguyen, M.H.; et al. Clinical Practice Guidelines for the Management of Cryptococcal Disease: 2010 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2010, 50, 291–322.
- Delma, F.Z.; Al-Hatmi, A.M.S.; Brüggemann, R.J.M.; Melchers, W.J.G.; de Hoog, S.; Verweij, P.E.; Buil, J.B. Molecular Mechanisms of 5-Fluorocytosine Resistance in Yeasts and Filamentous Fungi. J. Fungi 2021, 7, 909.
- Bhattacharya, S.; Esquivel, B.D.; White, T.C. Overexpression or Deletion of Ergosterol Biosynthesis Genes Alters Doubling Time, Response to Stress Agents, and Drug Susceptibility in Saccharomyces Cerevisiae. mBio 2018, 9, e01291-18.
- Xiang, M.J.; Liu, J.Y.; Ni, P.H.; Wang, S.; Shi, C.; Wei, B.; Ni, Y.X.; Ge, H.L. Erg11 Mutations Associated with Azole Resistance in Clinical Isolates of Candida Albicans. FEMS Yeast Res. 2013, 13, 386–393.
- Manastir, L.; Ergon, M.C.; Yücesoy, M. Investigation of Mutations in Erg11 Gene of Fluconazole Resistant Candida Albicans Isolates from Turkish Hospitals. Mycoses 2011, 54, 99–104.
- Morio, F.; Loge, C.; Besse, B.; Hennequin, C.; Le Pape, P. Screening for Amino Acid Substitutions in the Candida Albicans Erg11 Protein of Azole-Susceptible and Azole-Resistant Clinical Isolates: New Substitutions and a Review of the Literature. Diagn. Microbiol. Infect. Dis. 2010, 66, 373–384.
- Feng, L.J.; Wan, Z.; Wang, X.H.; Li, R.Y.; Liu, W. Relationship between Antifungal Resistance of Fluconazole Resistant Candida Albicans and Mutations in ERG11 Gene. Chin. Med. J. 2010, 123, 544–548.
- dos Santos Silva, D.B.; Carbonera Rodrigues, L.M.; De Almeida, A.A.; de Oliveira, K.M.P.; Grisolia, A.B. Novel Point Mutations in the ERG11 Gene in Clinical Isolates of Azole Resistant Candida Species. Mem. Inst. Oswaldo Cruz 2016, 111, 192–199.
- Sanglard, D.; Ischer, F.; Calabrese, D.; Micheli, M.d.; Bille, J. Multiple Resistance Mechanisms to Azole Antifungals in Yeast Clinical Isolates. Drug Resist. Updates 1998, 1, 255–265.
- Sanglard, D.; Ischer, F.; Koymans, L.; Bille, J. Amino Acid Substitutions in the Cytochrome P-450 Lanosterol 14α- Demethylase (CYP51A1) from Azole-Resistant Candida Albicans Clinical Isolates Contribute to Resistance to Azole Antifungal Agents. Antimicrob. Agents Chemother. 1998, 42, 241–253.
- Li, X.; Brown, N.; Chau, A.S.; López-Ribot, J.L.; Ruesga, M.T.; Quindos, G.; Mendrick, C.A.; Hare, R.S.; Loebenberg, D.; DiDomenico, B.; et al. Changes in Susceptibility to Posaconazole in Clinical Isolates of Candida Albicans. J. Antimicrob. Chemother. 2004, 53, 74–80.
- Dunkel, N.; Liu, T.T.; Barker, K.S.; Homayouni, R.; Morschhäuser, J.; Rogers, P.D. A Gain-of-Function Mutation in the Transcription Factor Upc2p Causes Upregulation of Ergosterol Biosynthesis Genes and Increased Fluconazole Resistance in a Clinical Candida Albicans Isolate. Eukaryot. Cell 2008, 7, 1180–1190.
- Hoot, S.J.; Smith, A.R.; Brown, R.P.; White, T.C. An A643V Amino Acid Substitution in Upc2p Contributes to Azole Resistance in Well-Characterized Clinical Isolates of Candida Albicans. Antimicrob. Agents Chemother. 2011, 55, 940–942.
- Znaidi, S.; Weber, S.; Al-Abdin, O.Z.; Bomme, P.; Saidane, S.; Drouin, S.; Lemieux, S.; De Deken, X.; Robert, F.; Raymond, M. Genomewide Location Analysis of Candida Albicans Upc2p, a Regulator of Sterol Metabolism and Azole Drug Resistance. Eukaryot. Cell 2008, 7, 836–847.
- Barker, K.S.; Crisp, S.; Wiederhold, N.; Lewis, R.E.; Bareither, B.; Eckstein, J.; Barbuch, R.; Bard, M.; Rogers, P.D. Genome-Wide Expression Profiling Reveals Genes Associated with Amphotericin B and Fluconazole Resistance in Experimentally Induced Antifungal Resistant Isolates of Candida Albicans. J. Antimicrob. Chemother. 2004, 54, 376–385.
- Kelly, S.L.; Lamb, D.C.; Corran, A.J.; Baldwin, B.C.; Kelly, D.E. Mode of Action and Resistance to Azole Antifungals Associated with the Formation of 14α-Methylergosta-8,24(28)-Dien-3β,6α-Diol. Biochem. Biophys. Res. Commun. 1995, 207, 910–915.
- Watson, P.F.; Rose, M.E.; Ellis, S.W.; England, H.; Kelly, S.L. Defective Sterol C5-6 Desaturation and Azole Resistance: A New Hypothesis for the Mode of Action of Azole Antifungals. Biochem. Biophys. Res. Commun. 1989, 164, 1170–1175.
- Sanglard, D.; Ischer, F.; Parkinson, T.; Falconer, D.; Bille, J. Candida Albicans Mutations in the Ergosterol Biosynthetic Pathway and Resistance to Several Antifungal Agents. Antimicrob. Agents Chemother. 2003, 47, 2404–2412.
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole Antifungal Resistance in Candida Albicans and Emerging Non-Albicans Candida Species. Front. Microbiol. 2017, 7, 2173.
- Yoo, J.I.; Choi, C.W.; Lee, K.M.; Lee, Y.S. Gene Expression and Identification Related to Fluconazole Resistance of Candida Glabrata Strains. Osong Public. Health Res. Perspect. 2010, 1, 36–41.
- Anderson, J.B.; Sirjusingh, C.; Parsons, A.B.; Boone, C.; Wickens, C.; Cowen, L.E.; Kohn, L.M. Mode of Selection and Experimental Evolution of Antifungal Drug Resistance in Saccharomyces Cerevisiae. Genetics 2003, 163, 1287–1298.
- Xu, D.; Jiang, B.; Ketela, T.; Lemieux, S.; Veillette, K.; Martel, N.; Davison, J.; Sillaots, S.; Trosok, S.; Bachewich, C.; et al. Genome-Wide Fitness Test and Mechanism-of-Action Studies of Inhibitory Compounds in Candida Albicans. PLoS Pathog. 2007, 3, e92.
- Jia, Y.; Tang, R.J.; Wang, L.; Zhang, X.; Wang, Y.; Jia, X.M.; Jiang, Y.Y. Calcium-Activated-Calcineurin Reduces the In Vitro and In Vivo Sensitivity of Fluconazole to Candida Albicans via Rta2p. PLoS ONE 2012, 7, e48369.
- Perea, S.; López-Ribot, J.L.; Kirkpatrick, W.R.; McAtee, R.K.; Santillán, R.A.; Martínez, M.; Calabrese, D.; Sanglard, D.; Patterson, T.F. Prevalence of Molecular Mechanisms of Resistance to Azole Antifungal Agents in Candida Albicans Strains Displaying High-Level Fluconazole Resistance Isolated from Human Immunodeficiency Virus-Infected Patients. Antimicrob. Agents Chemother. 2001, 45, 2676–2684.
- Prasad, R.; Nair, R.; Banerjee, A. Multidrug Transporters of Candida Species in Clinical Azole Resistance. Fungal Genet. Biol. 2019, 132, 103252.
- Sanglard, D.; Ischer, F.; Monod, M.; Bille, J. Susceptibilities of Candida Albicans Multidrug Transporter Mutants to Various Antifungal Agents and Other Metabolic Inhibitors. Antimicrob. Agents Chemother. 1996, 40, 2300–2305.
- Khandelwal, N.K.; Wasi, M.; Nair, R.; Gupta, M.; Kumar, M.; Mondal, A.K.; Gaur, N.A.; Prasad, R. Vacuolar Sequestration of Azoles, a Novel Strategy of Azole Antifungal Resistance Conserved across Pathogenic and Nonpathogenic Yeast. Antimicrob. Agents Chemother. 2019, 63, e01347-18.
- Bencova, A.; Goffa, E.; Morvova, M.; Valachovic, M.; Griač, P.; Toth Hervay, N.; Gbelska, Y. The Absence of PDR16 Gene Restricts the Overexpression of CaSNQ2 Gene in the Presence of Fluconazole in Candida Albicans. Mycopathologia 2020, 185, 455–465.
- Znaidi, S.; De Deken, X.; Weber, S.; Rigby, T.; Nantel, A.; Raymond, M. The Zinc Cluster Transcription Factor Tac1p Regulates PDR16 Expression in Candida Albicans. Mol. Microbiol. 2007, 66, 440–452.
- Perepnikhatka, V.; Fischer, F.J.; Niimi, M.; Baker, R.A.; Cannon, R.D.; Wang, Y.K.; Sherman, F.; Rustchenko, E. Specific Chromosome Alterations in Fluconazole-Resistant Mutants of Candida Albicans. J. Bacteriol. 1999, 181, 4041–4049.
- Sanguinetti, M.; Posteraro, B.; Fiori, B.; Ranno, S.; Torelli, R.; Fadda, G. Mechanisms of Azole Resistance in Clinical Isolates of Candida Glabrata Collected during a Hospital Survey of Antifungal Resistance. Antimicrob. Agents Chemother. 2005, 49, 668–679.
- Miyazaki, H.; Miyazaki, Y.; Geber, A.; Parkinson, T.; Hitchcock, C.; Falconer, D.J.; Ward, D.J.; Marsden, K.; Bennett, J.E. Fluconazole Resistance Associated with Drug Efflux and Increased Transcription of a Drug Transporter Gene, PDH1, in Candida Glabrata. Antimicrob. Agents Chemother. 1998, 42, 1695–1701.
- Whaley, S.G.; Zhang, Q.; Caudle, K.E.; Rogers, P.D. Relative Contribution of the ABC Transporters Cdr1, Pdh1, and Snq2 to Azole Resistance in Candida Glabrata. Antimicrob. Agents Chemother. 2018, 62, e01070-18.
- Costa, C.; Dias, P.J.; Sá-Correia, I.; Teixeira, M.C. MFS Multidrug Transporters in Pathogenic Fungi: Do They Have Real Clinical Impact? Front. Physiol. 2014, 5, 197.
- Costa, C.; Ribeiro, J.; Miranda, I.M.; Silva-Dias, A.; Cavalheiro, M.; Costa-de-Oliveira, S.; Rodrigues, A.G.; Teixeira, M.C. Clotrimazole Drug Resistance in Candida Glabrata Clinical Isolates Correlates with Increased Expression of the Drug: H+ Antiporters CgAqr1, CgTpo1_1, CgTpo3, and CgQdr2. Front. Microbiol. 2016, 7, 526.
- Bhattacharya, S.; Friesa, B.C. Enhanced Efflux Pump Activity in Old Candida Glabrata Cells. Antimicrob. Agents Chemother. 2018, 62, e02227-17.
- Culakova, H.; Dzugasova, V.; Valencikova, R.; Gbelska, Y.; Subik, J. Stress Response and Expression of Fluconazole Resistance Associated Genes in the Pathogenic Yeast Candida Glabrata Deleted in the CgPDR16 Gene. Microbiol. Res. 2015, 174, 17–23.
- Coste, A.; Turner, V.; Ischer, F.; Morschhäuser, J.; Forche, A.; Selmecki, A.; Berman, J.; Bille, J.; Sanglard, D. A Mutation in Tac1p, a Transcription Factor Regulating CDR1 and CDR2, Is Coupled with Loss of Heterozygosity at Chromosome 5 to Mediate Antifungal Resistance in Candida Albicans. Genetics 2006, 172, 2139–2156.
- Rybak, J.M.; Muñoz, J.F.; Barker, K.S.; Parker, J.E.; Esquivel, B.D.; Berkow, E.L.; Lockhart, S.R.; Gade, L.; Palmer, G.E.; White, T.C.; et al. Mutations in TAC1B: A Novel Genetic Determinant of Clinical Fluconazole Resistance in Candida Auris. mBio 2020, 11, e00365-20.
- Kalkandelen, K.T.; Doluca Dereli, M. Investigation of Mutations in Transcription Factors of Efflux Pump Genes in Fluconazole-Resistant Candida Albicans Strains Overexpressing the Efflux Pumps. Mikrobiyol. Bul. 2015, 49, 609–618.
- Schubert, S.; Rogers, P.D.; Morschhäuser, J. Gain-of-Function Mutations in the Transcription Factor MRR1 Are Responsible for Overexpression of the MDR1 Efflux Pump in Fluconazole-Resistant Candida Dubliniensis Strains. Antimicrob. Agents Chemother. 2008, 52, 4274–4280.
- Moran, G.P.; Sullivan, D.J.; Henman, M.C.; McCreary, C.E.; Harrington, B.J.; Shanley, D.B.; Coleman, D.C. Antifungal Drug Susceptibilities of Oral Candida Dubliniensis Isolates from Human Immunodeficiency Virus (HIV)-Infected and Non-HIV-Infected Subjects and Generation of Stable Fluconazole-Resistant Derivatives in Vitro. Antimicrob. Agents Chemother. 1997, 41, 617–623.
- Moran, G.P.; Sanglard, D.; Donnelly, S.M.; Shanley, D.B.; Sullivan, D.J.; Coleman, D.C. Identification and Expression of Multidrug Transporters Responsible for Fluconazole Resistance in Candida Dubliniensis. Antimicrob. Agents Chemother. 1998, 42, 1819–1830.
- Morschhäuser, J.; Barker, K.S.; Liu, T.T.; Blaß-Warmuth, J.; Homayouni, R.; Rogers, P.D. The Transcription Factor Mrr1p Controls Expression of the MDR1 Efflux Pump and Mediates Multidrug Resistance in Candida Albicans. PLoS Pathog. 2007, 3, e164.
- Schubert, S.; Barker, K.S.; Znaidi, S.; Schneider, S.; Dierolf, F.; Dunkel, N.; Aïd, M.; Boucher, G.; Rogers, P.D.; Raymond, M.; et al. Regulation of Efflux Pump Expression and Drug Resistance by the Transcription Factors Mrr1, Upc2, and Cap1 in Candida Albicans. Antimicrob. Agents Chemother. 2011, 55, 2212–2223.
- Dunkel, N.; Blaß, J.; Rogers, P.D.; Morschhäuser, J. Mutations in the Multi-Drug Resistance Regulator MRR1, Followed by Loss of Heterozygosity, Are the Main Cause of MDR1 Overexpression in Fluconazole-Resistant Candida Albicans Strains. Mol. Microbiol. 2008, 69, 827–840.
- Shukla, S.; Yadav, V.; Mukhopadhyay, G.; Prasad, R. Ncb2 Is Involved in Activated Transcription of CDR1 in Azole-Resistant Clinical Isolates of Candida Albicans ∇. Eukaryot. Cell 2011, 10, 1357–1366.
- Mogavero, S.; Tavanti, A.; Senesi, S.; Rogers, P.D.; Morschhäuser, J. Differential Requirement of the Transcription Factor Mcm1 for Activation of the Candida Albicans Multidrug Efflux Pump MDR1 by Its Regulators Mrr1 and Cap1. Antimicrob. Agents Chemother. 2011, 55, 2061–2066.
- Alarco, A.M.; Raymond, M. The BZip Transcription Factor Cap1p Is Involved in Multidrug Resistance and Oxidative Stress Response in Candida Albicans. J. Bacteriol. 1999, 181, 700–708.
- Thomas, E.; Roman, E.; Claypool, S.; Manzoor, N.; Pla, J.; Panwar, S.L. Mitochondria Influence CDR1 Efflux Pump Activity, Hog1-Mediated Oxidative Stress Pathway, Iron Homeostasis, and Ergosterol Levels in Candida Albicans. Antimicrob. Agents Chemother. 2013, 57, 5580–5599.
- Alexander, B.D.; Johnson, M.D.; Pfeiffer, C.D.; Jiménez-Ortigosa, C.; Catania, J.; Booker, R.; Castanheira, M.; Messer, S.A.; Perlin, D.S.; Pfaller, M.A. Increasing Echinocandin Resistance in Candida Glabrata: Clinical Failure Correlates with Presence of FKS Mutations and Elevated Minimum Inhibitory Concentrations. Clin. Infect. Dis. 2013, 56, 1724–1732.
- Cleary, J.D.; Garcia-Effron, G.; Chapman, S.W.; Perlin, D.S. Reduced Candida Glabrata Susceptibility Secondary to an FKS1 Mutation Developed during Candidemia Treatment. Antimicrob. Agents Chemother. 2008, 52, 2263–2265.
- Garcia-Effron, G.; Lee, S.; Park, S.; Cleary, J.D.; Perlin, D.S. Effect of Candida Glabrata FKS1 and FKS2 Mutations on Echinocandin Sensitivity and Kinetics of 1,3-β-D-Glucan Synthase: Implication for the Existing Susceptibility Breakpoint. Antimicrob. Agents Chemother. 2009, 53, 3690–3699.
- Garcia-Effron, G.; Chua, D.J.; Tomada, J.R.; DiPersio, J.; Perlin, D.S.; Ghannoum, M.; Bonilla, H. Novel FKS Mutations Associated with Echinocandin Resistance in Candida Species. Antimicrob. Agents Chemother. 2010, 54, 2225–2227.
- Thompson, G.R.; Wiederhold, N.P.; Vallor, A.C.; Villareal, N.C.; Lewis, J.S.; Patterson, T.F. Development of Caspofungin Resistance Following Prolonged Therapy for Invasive Candidiasis Secondary to Candida Glabrata Infection. Antimicrob. Agents Chemother. 2008, 52, 3783–3785.
- Munro, C.A. Fungal Echinocandin Resistance. F1000 Biol. Rep. 2010, 2, 117–126.
- Park, S.; Kelly, R.; Kahn, J.N.; Robles, J.; Hsu, M.J.; Register, E.; Li, W.; Vyas, V.; Fan, H.; Abruzzo, G.; et al. Specific Substitutions in the Echinocandin Target Fks1p Account for Reduced Susceptibility of Rare Laboratory and Clinical Candida sp. Isolates. Antimicrob. Agents Chemother. 2005, 49, 3264–3273.
- Perlin, D.S. Resistance to Echinocandin-Class Antifungal Drugs. Drug Resist. Updates 2007, 10, 121–130.
- Hakki, M.; Staab, J.F.; Marr, K.A. Emergence of a Candida Krusei Isolate with Reduced Susceptibility to Caspofungin during Therapy. Antimicrob. Agents Chemother. 2006, 50, 2522–2524.
- Kahn, J.N.; Garcia-Effron, G.; Hsu, M.J.; Park, S.; Marr, K.A.; Perlin, D.S. Acquired Echinocandin Resistance in a Candida Krusei Isolate Due to Modification of Glucan Synthase. Antimicrob. Agents Chemother. 2007, 51, 1876–1878.
- Garcia-Effron, G.; Park, S.; Perlin, D.S. Correlating Echinocandin MIC and Kinetic Inhibition of Fks1 Mutant Glucan Synthases for Candida Albicans: Implications for Interpretive Breakpoints. Antimicrob. Agents Chemother. 2009, 53, 112–122.
- Sharma, D.; Paul, R.A.; Rudramurthy, S.M.; Kashyap, N.; Bhattacharya, S.; Soman, R.; Shankarnarayan, S.A.; Chavan, D.; Singh, S.; Das, P.; et al. Impact of FKS1 Genotype on Echinocandin In Vitro Susceptibility in Candida Auris and In Vivo Response in a Murine Model of Infection. Antimicrob. Agents Chemother. 2022, 66, e0165221.
- Balashov, S.V.; Park, S.; Perlin, D.S. Assessing Resistance to the Echinocandin Antifungal Drug Caspofungin in Candida Albicans by Profiling Mutations in FKS1. Antimicrob. Agents Chemother. 2006, 50, 2058–2063.
- Katiyar, S.; Pfaller, M.; Edlind, T. Candida Albicans and Candida Glabrata Clinical Isolates Exhibiting Reduced Echinocandin Susceptibility. Antimicrob. Agents Chemother. 2006, 50, 2892–2894.
- Pardini, G.; De Groot, P.W.J.; Coste, A.T.; Karababa, M.; Klis, F.M.; De Koster, C.G.; Sanglard, D. The CRH Family Coding for Cell Wall Glycosylphosphatidylinositol Proteins with a Predicted Transglycosidase Domain Affects Cell Wall Organization and Virulence of Candida Albicans. J. Biol. Chem. 2006, 281, 40399–40411.
- Fonzi, W.A. PHR1 and PHR2 of Candida Albicans Encode Putative Glycosidases Required for Proper Cross-Linking of β-1,3- and β-1,6-Glucans. J. Bacteriol. 1999, 181, 7070–7079.
- Yu, Z.W.; Quinn, P.J. Solvation Effects of Dimethyl Sulphoxide on the Structure of Phospholipid Bilayers. Biophys. Chem. 1998, 70, 35–39.
- De Cesare, G.B.; Hafez, A.; Stead, D.; Llorens, C.; Munro, C.A. Biomarkers of Caspofungin Resistance in Candida Albicans Isolates: A Proteomic Approach. Virulence 2022, 13, 1005–1018.
- Caplan, T.; Polvi, E.J.; Xie, J.L.; Buckhalter, S.; Leach, M.D.; Robbins, N.; Cowen, L.E. Functional Genomic Screening Reveals Core Modulators of Echinocandin Stress Responses in Candida Albicans. Cell Rep. 2018, 23, 2292–2298.
- Kelly, J.; Kavanagh, K. Proteomic Analysis of Proteins Released from Growth-Arrested Candida Albicans Following Exposure to Caspofungin. Med. Mycol. 2010, 48, 598–605.
- Lafayette, S.L.; Collins, C.; Zaas, A.K.; Schell, W.A.; Betancourt-Quiroz, M.; Leslie Gunatilaka, A.A.; Perfect, J.R.; Cowen, L.E. PKC Signaling Regulates Drug Resistance of the Fungal Pathogen Candida Albicans via Circuitry Comprised of Mkc1, Calcineurin, and Hsp90. PLoS Pathog. 2010, 6, e1001069.
- Cowen, L.E.; Steinbach, W.J. Stress, Drugs, and Evolution: The Role of Cellular Signaling in Fungal Drug Resistance. Eukaryot. Cell 2008, 7, 747–764.
- Erbs, P.; Exinger, F.; Jund, R. Characterization of the Saccaromyces Cerevisiae FCY1 Gene Encoding Cytosine Deaminase and Its Homologue FCA1 of Candida Albicans. Curr. Genet. 1997, 31, 1–6.
- McManus, B.A.; Moran, G.P.; Higgins, J.A.; Sullivan, D.J.; Coleman, D.C. A Ser29Leu Substitution in the Cytosine Deaminase Fca1p Is Responsible for Clade-Specific Flucytosine Resistance in Candida Dubliniensis. Antimicrob. Agents Chemother. 2009, 53, 4678–4685.
- Lestrade, P.P.; Bentvelsen, R.G.; Schauwvlieghe, A.F.A.D.; Schalekamp, S.; Van Der Velden, W.J.F.M.; Kuiper, E.J.; Van Paassen, J.; Van Der Hoven, B.; Van Der Lee, H.A.; Melchers, W.J.G.; et al. Voriconazole Resistance and Mortality in Invasive Aspergillosis: A Multicenter Retrospective Cohort Study. Clin. Infect. Dis. 2019, 68, 1463–1471.
- Jund, R.; Lacroute, F. Genetic and Physiological Aspects of Resistance to 5-Fluoropyrimidines in Saccharomyces Cerevisiae. J. Bacteriol. 1970, 102, 607–615.
- Chevallier, M.R.; Jund, R.; Lacroute, F. Characterization of Cytosine Permeation in Saccharomyces Cerevisiae. J. Bacteriol. 1975, 122, 629–641.
- Kern, L.; de Montigny, J.; Lacroute, F.; Jund, R. Regulation of the Pyrimidine Salvage Pathway by the FUR1 Gene Product of Saccharomyces Cerevisiae. Curr. Genet. 1991, 19, 333–337.
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm Formation by the Fungal Pathogen Candida Albicans: Development, Architecture, and Drug Resistance. J. Bacteriol. 2001, 183, 5385–5394.
- Rajendran, R.; Sherry, L.; Nile, C.J.; Sherriff, A.; Johnson, E.M.; Hanson, M.F.; Williams, C.; Munro, C.A.; Jones, B.J.; Ramage, G. Biofilm Formation Is a Risk Factor for Mortality in Patients with Candida Albicans Bloodstream Infection-Scotland, 2012–2013. Clin. Microbiol. Infect. 2016, 22, 87–93.
- Ramage, G.; Vande Walle, K.; Wickes, B.L.; López-Ribot, J.L. Standardized Method for in Vitro Antifungal Susceptibility Testing of Candida Albicans Biofilms. Antimicrob. Agents Chemother. 2001, 45, 2475–2479.
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C. Fungal Biofilm Resistance. Int. J. Microbiol. 2012, 2012, 1–14.
- Fan, F.M.; Liu, Y.; Liu, Y.Q.; Lv, R.X.; Sun, W.; Ding, W.J.; Cai, Y.X.; Li, W.W.; Liu, X.; Qu, W. Candida Albicans Biofilms: Antifungal Resistance, Immune Evasion, and Emerging Therapeutic Strategies. Int. J. Antimicrob. Agents 2022, 60, 106673.
- Perumal, P.; Mekala, S.; Chaffin, W.L.J. Role for Cell Density in Antifungal Drug Resistance in Candida Albicans Biofilms. Antimicrob. Agents Chemother. 2007, 51, 2454–2463.
- Nett, J.; Lincoln, L.; Marchillo, K.; Massey, R.; Holoyda, K.; Hoff, B.; VanHandel, M.; Andes, D. Putative Role of β-1,3 Glucans in Candida Albicans Biofilm Resistance. Antimicrob. Agents Chemother. 2007, 51, 510–520.
- Nett, J.E.; Sanchez, H.; Cain, M.T.; Andes, D.R. Genetic Basis of Candida Biofilm Resistance Due to Drug-Sequestering Matrix Glucan. J. Infect. Dis. 2010, 202, 171–175.
- Tan, Y.; Ma, S.; Leonhard, M.; Moser, D.; Schneider-Stickler, B. β-1,3-Glucanase Disrupts Biofilm Formation and Increases Antifungal Susceptibility of Candida Albicans DAY185. Int. J. Biol. Macromol. 2018, 108, 942–946.
- Al-Fattani, M.A.; Douglas, L.J. Biofilm Matrix of Candida Albicans and Candida Tropicalis: Chemical Composition and Role in Drug Resistance. J. Med. Microbiol. 2006, 55, 999–1008.
- Nailis, H.; Vandenbosch, D.; Deforce, D.; Nelis, H.J.; Coenye, T. Transcriptional Response to Fluconazole and Amphotericin B in Candida Albicans Biofilms. Res. Microbiol. 2010, 161, 284–292.
- Borecká-Melkusová, S.; Moran, G.P.; Sullivan, D.J.; Kucharíková, S.; Chorvát, D.; Bujdáková, H. The Expression of Genes Involved in the Ergosterol Biosynthesis Pathway in Candida Albicans and Candida Dubliniensis Biofilms Exposed to Fluconazole. Mycoses 2009, 52, 118–128.
- Katragkou, A.; Chatzimoschou, A.; Simitsopoulou, M.; Dalakiouridou, M.; Diza-Mataftsi, E.; Tsantali, C.; Roilides, E. Differential Activities of Newer Antifungal Agents against Candida Albicans and Candida Parapsilosis Biofilms. Antimicrob. Agents Chemother. 2008, 52, 357–360.
- Rossignol, T.; Ding, C.; Guida, A.; D’Enfert, C.; Higgins, D.G.; Butler, G. Correlation between Biofilm Formation and the Hypoxic Response in Candida Parapsilosis. Eukaryot. Cell 2009, 8, 550–559.
- Knot, P.D.; Suci, P.A.; Miller, R.L.; Nelson, R.D.; Tyler, B.J. A Small Subpopulation of Blastospores in Candida Albicans Biofilms Exhibit Resistance to Amphotericin B Associated with Differential Regulation of Ergosterol and β-1,6-Glucan Pathway Genes. Antimicrob. Agents Chemother. 2006, 50, 3708–3716.
- Lepak, A.; Nett, J.; Lincoln, L.; Marchillo, K.; Andes, D. Time Course of Microbiologic Outcome and Gene Expression in Candida Albicans during and Following in Vitro and in Vivo Exposure to Fluconazole. Antimicrob. Agents Chemother. 2006, 50, 1311–1319.
- Mukherjee, P.K.; Chandra, J.; Kuhn, D.M.; Ghannoum, M.A. Mechanism of Fluconazole Resistance in Candida Albicans Biofilms: Phase-Specific Role of Efflux Pumps and Membrane Sterols. Infect. Immun. 2003, 71, 4333–4340.
- Ramage, G.; Bachmann, S.; Patterson, T.F.; Wickes, B.L.; López-Ribot, J.L. Investigation of Multidrug Efflux Pumps in Relation to Fluconazole Resistance in Candida Albicans Biofilms. J. Antimicrob. Chemother. 2002, 49, 973–980.
- Mateus, C.; Crow, S.A.; Ahearn, D.G. Adherence of Candida Albicans to Silicone Induces Immediate Enhanced Tolerance to Fluconazole. Antimicrob. Agents Chemother. 2004, 48, 3358–3366.
- Bizerra, F.C.; Nakamura, C.V.; De Poersch, C.; Estivalet Svidzinski, T.I.; Borsato Quesada, R.M.; Goldenberg, S.; Krieger, M.A.; Yamada-Ogatta, S.F. Characteristics of Biofilm Formation by Candida Tropicalis and Antifungal Resistance. FEMS Yeast Res. 2008, 8, 442–450.
- Song, J.W.; Shin, J.H.; Kee, S.J.; Kim, S.H.; Shin, M.G.; Suh, S.P.; Ryang, D.W. Expression of CgCDR1, CgCDR2, and CgERG11 in Candida Glabrata Biofilms Formed by Bloodstream Isolates. Med. Mycol. 2009, 47, 545–548.
- Andes, D.; Nett, J.; Oschel, P.; Albrecht, R.; Marchillo, K.; Pitula, A. Development and Characterization of an in Vivo Central Venous Catheter Candida Albicans Biofilm Model. Infect. Immun. 2004, 72, 6023–6031.
- Marichal, P.; Koymans, L.; Willemsens, S.; Bellens, D.; Verhasselt, P.; Luyten, W.; Borgers, M.; Ramaekers, F.C.S.; Odds, F.C.; Bossche, H. Vanden Contribution of Mutations in the Cytochrome P450 14α-Demethylase (Erg11p, Cyp51p) to Azole Resistance in Candida Albicans. Microbiol. 1999, 145, 2701–2713.
- Xu, Z.; Zhang, L.X.; Zhang, J.D.; Cao, Y.B.; Yu, Y.Y.; Wang, D.J.; Ying, K.; Chen, W.S.; Jiang, Y.Y. CDNA Microarray Analysis of Differential Gene Expression and Regulation in Clinically Drug-Resistant Isolates of Candida Albicans from Bone Marrow Transplanted Patients. Int. J. Med. Microbiol. 2006, 296, 421–434.

