 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Caterina Tezze | -- | 4415 | 2023-10-19 17:48:24 | | | |
| 2 | Lindsay Dong | + 2 word(s) | 4417 | 2023-10-23 03:24:34 | | | | |
| 3 | Lindsay Dong | Meta information modification | 4417 | 2023-10-30 09:44:10 | | |
Video Upload Options
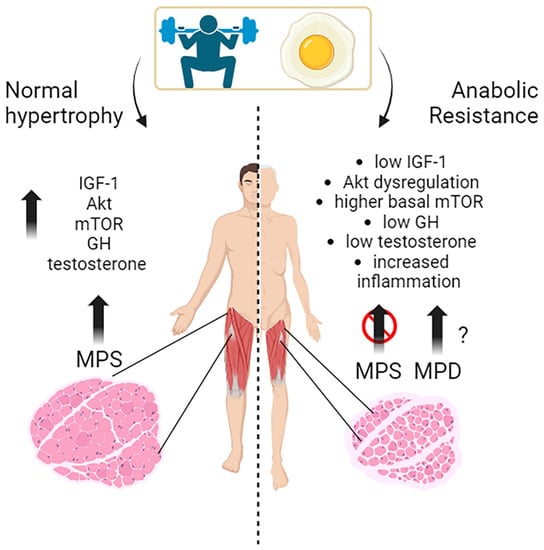
Muscle growth, or muscle hypertrophy, is a complex process regulated by several molecular pathways. The IGF-1/PI3K/Akt/mTOR pathway is a vital signaling cascade in muscle growth that involves various interconnected mechanisms. Its activation increases protein synthesis, reduces protein degradation, and improves cell growth. Akt activation is crucial in promoting muscle protein synthesis in response to exercise and nutrient intake in young individuals. The timing of exercise and protein intake also affect Akt activation and subsequent muscle protein synthesis. While exercise alone did not increase Akt and mTOR phosphorylation, protein ingestion afterward did so in a dose-dependent manner. Growth hormone (GH) promotes the uptake of essential nutrients, such as glucose and amino acids, into muscle cells for energy production and protein synthesis. Testosterone is one of the most potent naturally secreted androgenic-anabolic hormones, and its biological effects include promoting muscle growth.
1. The Molecular Mechanisms behind Muscles Growth in Young Subjects: Exercise, Nutrients, and Hormones
2. Effects of Substrates and Exercise on Skeletal Muscle Protein Synthesis in Young, Middle-Aged Subjects
2.1. Effects of Proteins and Amino Acids
2.2. Effects of Exercise and Nutrition on Muscle Protein Synthesis and Accretion
2.3. Effect of Other Substrates
Glucose
Lipids and Ketones
Other Nutritional Interventions
3. Hormones and Related Drug Interventions
3.1. Insulin
3.2. Glucocorticoids
3.3. Human Growth Hormone (hGH) and IGF-1
3.4. Catecholamines
3.5. Estrogens
3.6. Androgens
Testosterone is a potent anabolic stimulus primarily through improvement in the re-utilization of amino acids released from protein degradation [83] (see also the above paragraph), and this will be further discussed below. Testosterone and progesterone, but not estradiol, stimulated muscle protein synthesis in postmenopausal women [84].
4. Exercise
4.1. Resistance Exercise Can Be Further Classified into Two Main Categories
4.2. Effects of Exercise in Conjunction with Nutrient Intake
4.3. Exercise–Insulin Interaction
References
- Yoshida, T.; Delafontaine, P. Mechanisms of IGF-1-Mediated Regulation of Skeletal Muscle Hypertrophy and Atrophy. Cells 2020, 9, 1970.
- Drummond, M.J.; Fry, C.S.; Glynn, E.L.; Dreyer, H.C.; Dhanani, S.; Timmerman, K.L.; Volpi, E.; Rasmussen, B.B. Rapamycin Administration in Humans Blocks the Contraction-Induced Increase in Skeletal Muscle Protein Synthesis: Rapamycin Blocks Protein Synthesis in Human Muscle. J. Physiol. 2009, 587, 1535–1546.
- Francaux, M.; Demeulder, B.; Naslain, D.; Fortin, R.; Lutz, O.; Caty, G.; Deldicque, L. Aging Reduces the Activation of the MTORC1 Pathway after Resistance Exercise and Protein Intake in Human Skeletal Muscle: Potential Role of REDD1 and Impaired Anabolic Sensitivity. Nutrients 2016, 8, 47.
- Léger, B.; Cartoni, R.; Praz, M.; Lamon, S.; Dériaz, O.; Crettenand, A.; Gobelet, C.; Rohmer, P.; Konzelmann, M.; Luthi, F.; et al. Akt Signalling through GSK-3beta, MTOR and Foxo1 Is Involved in Human Skeletal Muscle Hypertrophy and Atrophy. J. Physiol. 2006, 576, 923–933.
- Di Camillo, B.; Eduati, F.; Nair, S.K.; Avogaro, A.; Toffolo, G.M. Leucine Modulates Dynamic Phosphorylation Events in Insulin Signaling Pathway and Enhances Insulin-Dependent Glycogen Synthesis in Human Skeletal Muscle Cells. BMC Cell. Biol. 2014, 15, 9.
- Kakigi, R.; Yoshihara, T.; Ozaki, H.; Ogura, Y.; Ichinoseki-Sekine, N.; Kobayashi, H.; Naito, H. Whey Protein Intake after Resistance Exercise Activates MTOR Signaling in a Dose-Dependent Manner in Human Skeletal Muscle. Eur. J Appl. Physiol. 2014, 114, 735–742.
- Jacquemin, V.; Furling, D.; Bigot, A.; Butler-Browne, G.S.; Mouly, V. IGF-1 Induces Human Myotube Hypertrophy by Increasing Cell Recruitment. Exp. Cell Res. 2004, 299, 148–158.
- Barclay, R.D.; Burd, N.A.; Tyler, C.; Tillin, N.A.; Mackenzie, R.W. The Role of the IGF-1 Signaling Cascade in Muscle Protein Synthesis and Anabolic Resistance in Aging Skeletal Muscle. Front. Nutr. 2019, 6, 146.
- Hansen, M.; Koskinen, S.O.; Petersen, S.G.; Doessing, S.; Frystyk, J.; Flyvbjerg, A.; Westh, E.; Magnusson, S.P.; Kjaer, M.; Langberg, H. Ethinyl Oestradiol Administration in Women Suppresses Synthesis of Collagen in Tendon in Response to Exercise: Oral Contraceptives and Tendon Collagen. J. Physiol. 2008, 586, 3005–3016.
- Perrini, S.; Laviola, L.; Carreira, M.C.; Cignarelli, A.; Natalicchio, A.; Giorgino, F. The GH/IGF1 Axis and Signaling Pathways in the Muscle and Bone: Mechanisms Underlying Age-Related Skeletal Muscle Wasting and Osteoporosis. J. Endocrinol. 2010, 205, 201–210.
- Chikani, V.; Ho, K.K.Y. Action of GH on Skeletal Muscle Function: Molecular and Metabolic Mechanisms. J. Mol. Endocrinol. 2014, 52, R107–R123.
- West, D.W.D.; Phillips, S.M. Anabolic Processes in Human Skeletal Muscle: Restoring the Identities of Growth Hormone and Testosterone. Physician Sports Med. 2010, 38, 97–104.
- Florini, J.R. Effects of Testosterone on Qualitative Pattern of Protein Synthesis in Skeletal Muscle. Biochemistry 1970, 9, 909–912.
- Mauras, N.; Hayes, V.; Welch, S.; Rini, A.; Helgeson, K.; Dokler, M.; Veldhuis, J.D.; Urban, R.J. Testosterone Deficiency in Young Men: Marked Alterations in Whole Body Protein Kinetics, Strength, and Adiposity. J. Clin. Endocrinol. Metab. 1998, 83, 1886–1892.
- Demling, R.H.; Orgill, D.P. The Anticatabolic and Wound Healing Effects of the Testosterone Analog Oxandrolone after Severe Burn Injury. J. Crit. Care 2000, 15, 12–17.
- Vingren, J.L.; Kraemer, W.J.; Ratamess, N.A.; Anderson, J.M.; Volek, J.S.; Maresh, C.M. Testosterone Physiology in Resistance Exercise and Training: The Up-Stream Regulatory Elements. Sports Med. 2010, 40, 1037–1053.
- Vierck, J. Satellite Cell Regulation Following Myotrauma Caused by Resistance Exercise. Cell Biol. Int. 2000, 24, 263–272.
- Bhasin, S.; Woodhouse, L.; Casaburi, R.; Singh, A.B.; Bhasin, D.; Berman, N.; Chen, X.; Yarasheski, K.E.; Magliano, L.; Dzekov, C.; et al. Testosterone Dose-Response Relationships in Healthy Young Men. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E1172–E1181.
- Plotkin, D.L.; Delcastillo, K.; Van Every, D.W.; Tipton, K.D.; Aragon, A.A.; Schoenfeld, B.J. Isolated Leucine and Branched-Chain Amino Acid Supplementation for Enhancing Muscular Strength and Hypertrophy: A Narrative Review. Int. J. Sport Nutr. Exerc. Metab. 2021, 31, 292–301.
- Tezze, C.; Amendolagine, F.I.; Nogara, L.; Baraldo, M.; Ciciliot, S.; Arcidiacono, D.; Zaramella, A.; Masiero, G.; Ferrarese, G.; Realdon, S.; et al. A Combination of Metformin and Galantamine Exhibits Synergistic Benefits in the Treatment of Sarcopenia. JCI Insight 2023, 8, e168787.
- Malena, A.; Pennuto, M.; Tezze, C.; Querin, G.; D’Ascenzo, C.; Silani, V.; Cenacchi, G.; Scaramozza, A.; Romito, S.; Morandi, L.; et al. Androgen-Dependent Impairment of Myogenesis in Spinal and Bulbar Muscular Atrophy. Acta Neuropathol. 2013, 126, 109–121.
- Tezze, C.; Romanello, V.; Sandri, M. FGF21 as Modulator of Metabolism in Health and Disease. Front. Physiol. 2019, 10, 419.
- Tezze, C.; Romanello, V.; Desbats, M.A.; Fadini, G.P.; Albiero, M.; Favaro, G.; Ciciliot, S.; Soriano, M.E.; Morbidoni, V.; Cerqua, C.; et al. Age-Associated Loss of OPA1 in Muscle Impacts Muscle Mass, Metabolic Homeostasis, Systemic Inflammation, and Epithelial Senescence. Cell Metab. 2017, 25, 1374–1389.
- Solagna, F.; Tezze, C.; Lindenmeyer, M.T.; Lu, S.; Wu, G.; Liu, S.; Zhao, Y.; Mitchell, R.; Meyer, C.; Omairi, S.; et al. Pro-Cachectic Factors Link Experimental and Human Chronic Kidney Disease to Skeletal Muscle Wasting Programs. J. Clin. Investig. 2021, 131, e135821.
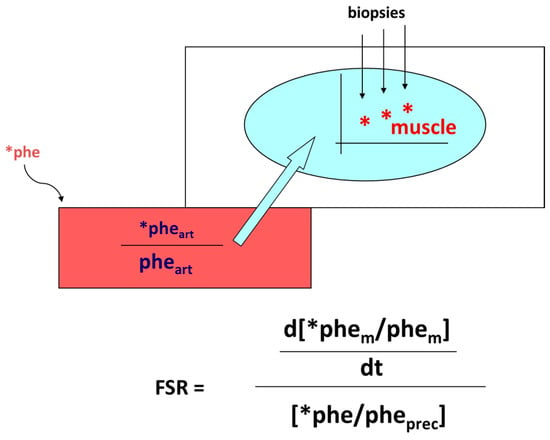
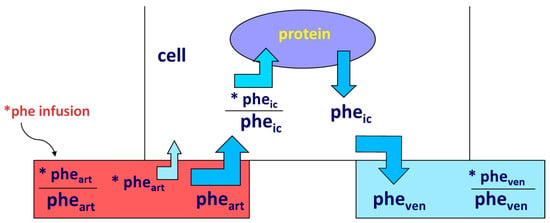
- Smith, K.; Rennie, M.J. The Measurement of Tissue Protein Turnover. Baillière’s Clin. Endocrinol. Metab. 1996, 10, 469–495.
- Gorissen, S.H.M.; Rémond, D.; Van Loon, L.J.C. The Muscle Protein Synthetic Response to Food Ingestion. Meat Sci. 2015, 109, 96–100.
- Wolfe, R.R. Regulation of Muscle Protein by Amino Acids. J. Nutr. 2002, 132, 3219S–3224S.
- Tang, J.E.; Moore, D.R.; Kujbida, G.W.; Tarnopolsky, M.A.; Phillips, S.M. Ingestion of Whey Hydrolysate, Casein, or Soy Protein Isolate: Effects on Mixed Muscle Protein Synthesis at Rest and Following Resistance Exercise in Young Men. J. Appl. Physiol. 2009, 107, 987–992.
- Bennet, W.M.; Connacher, A.A.; Scrimgeour, C.M.; Rennie, M.J. The Effect of Amino Acid Infusion on Leg Protein Turnover Assessed by L-Phenylalanine and L-Leucine Exchange. Eur. J. Clin. Investig. 2008, 20, 41–50.
- Tessari, P.; Barazzoni, R.; Zanetti, M.; Kiwanuka, E.; Tiengo, A. The Role of Substrates in the Regulation of Protein Metabolism. Baillière’s Clin. Endocrinol. Metab. 1996, 10, 511–532.
- Nygren, J.; Nair, K.S. Differential Regulation of Protein Dynamics in Splanchnic and Skeletal Muscle Beds by Insulin and Amino Acids in Healthy Human Subjects. Diabetes 2003, 52, 1377–1385.
- Volpi, E.; Kobayashi, H.; Sheffield-Moore, M.; Mittendorfer, B.; Wolfe, R.R. Essential Amino Acids Are Primarily Responsible for the Amino Acid Stimulation of Muscle Protein Anabolism in Healthy Elderly Adults. Am. J. Clin. Nutr. 2003, 78, 250–258.
- Kimball, S.R.; Jefferson, L.S. Regulation of Global and Specific MRNA Translation by Oral Administration of Branched-Chain Amino Acids. Biochem. Biophys. Res. Commun. 2004, 313, 423–427.
- Ham, D.J.; Caldow, M.K.; Lynch, G.S.; Koopman, R. Leucine as a Treatment for Muscle Wasting: A Critical Review. Clin. Nutr. 2014, 33, 937–945.
- Tessari, P.; Zanetti, M.; Barazzoni, R.; Vettore, M.; Michielan, F. Mechanisms of Postprandial Protein Accretion in Human Skeletal Muscle. Insight from Leucine and Phenylalanine Forearm Kinetics. J. Clin. Investig. 1996, 98, 1361–1372.
- Gwin, J.A.; Church, D.D.; Wolfe, R.R.; Ferrando, A.A.; Pasiakos, S.M. Muscle Protein Synthesis and Whole-Body Protein Turnover Responses to Ingesting Essential Amino Acids, Intact Protein, and Protein-Containing Mixed Meals with Considerations for Energy Deficit. Nutrients 2020, 12, 2457.
- Witard, O.C.; Jackman, S.R.; Breen, L.; Smith, K.; Selby, A.; Tipton, K.D. Myofibrillar Muscle Protein Synthesis Rates Subsequent to a Meal in Response to Increasing Doses of Whey Protein at Rest and after Resistance Exercise. Am. J. Clin. Nutr. 2014, 99, 86–95.
- Moore, D.R.; Tang, J.E.; Burd, N.A.; Rerecich, T.; Tarnopolsky, M.A.; Phillips, S.M. Differential Stimulation of Myofibrillar and Sarcoplasmic Protein Synthesis with Protein Ingestion at Rest and after Resistance Exercise. J. Physiol. 2009, 587, 897–904.
- Robinson, M.J.; Burd, N.A.; Breen, L.; Rerecich, T.; Yang, Y.; Hector, A.J.; Baker, S.K.; Phillips, S.M. Dose-Dependent Responses of Myofibrillar Protein Synthesis with Beef Ingestion Are Enhanced with Resistance Exercise in Middle-Aged Men. Appl. Physiol. Nutr. Metab. 2013, 38, 120–125.
- Paddon-Jones, D.; Sheffield-Moore, M.; Zhang, X.-J.; Volpi, E.; Wolf, S.E.; Aarsland, A.; Ferrando, A.A.; Wolfe, R.R. Amino Acid Ingestion Improves Muscle Protein Synthesis in the Young and Elderly. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E321–E328.
- Fujita, S.; Volpi, R. Nutrition and sarcopenia of ageing. Nutr. Res. Rev. 2004, 17, 69–76.
- Cuthbertson, D.; Smith, K.; Babraj, J.; Leese, G.; Waddell, T.; Atherton, P.; Wackerhage, H.; Taylor, P.M.; Rennie, M.J. Anabolic Signaling Deficits Underlie Amino Acid Resistance of Wasting, Aging Muscle. FASEB J. 2005, 19, 422–424.
- Deutz, N.E.P.; Bauer, J.M.; Barazzoni, R.; Biolo, G.; Boirie, Y.; Bosy-Westphal, A.; Cederholm, T.; Cruz-Jentoft, A.; Krznariç, Z.; Nair, K.S.; et al. Protein Intake and Exercise for Optimal Muscle Function with Aging: Recommendations from the ESPEN Expert Group. Clin. Nutr. 2014, 33, 929–936.
- Van Vliet, S.; Beals, J.W.; Holwerda, A.M.; Emmons, R.S.; Goessens, J.P.; Paluska, S.A.; De Lisio, M.; Van Loon, L.J.C.; Burd, N.A. Time-Dependent Regulation of Postprandial Muscle Protein Synthesis Rates after Milk Protein Ingestion in Young Men. J. Appl. Physiol. 2019, 127, 1792–1801.
- Monteyne, A.J.; Dunlop, M.V.; Machin, D.J.; Coelho, M.O.C.; Pavis, G.F.; Porter, C.; Murton, A.J.; Abdelrahman, D.R.; Dirks, M.L.; Stephens, F.B.; et al. A Mycoprotein-Based High-Protein Vegan Diet Supports Equivalent Daily Myofibrillar Protein Synthesis Rates Compared with an Isonitrogenous Omnivorous Diet in Older Adults: A Randomised Controlled Trial. Br. J. Nutr. 2021, 126, 674–684.
- Boirie, Y.; Dangin, M.; Gachon, P.; Vasson, M.-P.; Maubois, J.-L.; Beaufrère, B. Slow and Fast Dietary Proteins Differently Modulate Postprandial Protein Accretion. Proc. Natl. Acad. Sci. USA 1997, 94, 14930–14935.
- Dangin, M.; Boirie, Y.; Garcia-Rodenas, C.; Gachon, P.; Fauquant, J.; Callier, P.; Ballèvre, O.; Beaufrère, B. The Digestion Rate of Protein Is an Independent Regulating Factor of Postprandial Protein Retention. Am. J. Physiol. -Endocrinol. Metab. 2001, 280, E340–E348.
- Reidy, P.T.; Walker, D.K.; Dickinson, J.M.; Gundermann, D.M.; Drummond, M.J.; Timmerman, K.L.; Cope, M.B.; Mukherjea, R.; Jennings, K.; Volpi, E.; et al. Soy-Dairy Protein Blend and Whey Protein Ingestion after Resistance Exercise Increases Amino Acid Transport and Transporter Expression in Human Skeletal Muscle. J. Appl. Physiol. 2014, 116, 1353–1364.
- Tipton, K.D.; Wolfe, R.R. Exercise, Protein Metabolism, and Muscle Growth. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 109–132.
- Churchward-Venne, T.A.; Murphy, C.H.; Longland, T.M.; Phillips, S.M. Role of Protein and Amino Acids in Promoting Lean Mass Accretion with Resistance Exercise and Attenuating Lean Mass Loss during Energy Deficit in Humans. Amino Acids 2013, 45, 231–240.
- Pennings, B.; Boirie, Y.; Senden, J.M.; Gijsen, A.P.; Kuipers, H.; Van Loon, L.J. Whey Protein Stimulates Postprandial Muscle Protein Accretion More Effectively than Do Casein and Casein Hydrolysate in Older Men. Am. J. Clin. Nutr. 2011, 93, 997–1005.
- Macnaughton, L.S.; Wardle, S.L.; Witard, O.C.; McGlory, C.; Hamilton, D.L.; Jeromson, S.; Lawrence, C.E.; Wallis, G.A.; Tipton, K.D. The Response of Muscle Protein Synthesis Following Whole-Body Resistance Exercise Is Greater Following 40 g than 20 g of Ingested Whey Protein. Physiol. Rep. 2016, 4, e12893.
- Churchward-Venne, T.A.; Pinckaers, P.J.M.; Smeets, J.S.J.; Betz, M.W.; Senden, J.M.; Goessens, J.P.B.; Gijsen, A.P.; Rollo, I.; Verdijk, L.B.; van Loon, L.J.C. Dose-Response Effects of Dietary Protein on Muscle Protein Synthesis during Recovery from Endurance Exercise in Young Men: A Double-Blind Randomized Trial. Am. J. Clin. Nutr. 2020, 112, 303–317.
- Breen, L.; Phillips, S.M. Skeletal Muscle Protein Metabolism in the Elderly: Interventions to Counteract the “anabolic Resistance” of Ageing. Nutr. Metab. 2011, 8, 68.
- Reeds, P.J.; Burrin, D.G.; Davis, T.A.; Stoll, B. Amino Acid Metabolism and the Energetics of Growth. Arch. Anim. Nutr. 1998, 51, 187–197.
- Smiles, W.J.; Hawley, J.A.; Camera, D.M. Effects of Skeletal Muscle Energy Availability on Protein Turnover Responses to Exercise. J. Exp. Biol. 2016, 219, 214–225.
- Hedden, M.P.; Buse, M.G. Effects of Glucose, Pyruvate, Lactate, and Amino Acids on Muscle Protein Synthesis. Am. J. Physiol. 1982, 242, E184–E192.
- Goichon, A.; Coëffier, M.; Claeyssens, S.; Lecleire, S.; Cailleux, A.-F.; Bôle-Feysot, C.; Chan, P.; Donnadieu, N.; Lerebours, E.; Lavoinne, A.; et al. Effects of an Enteral Glucose Supply on Protein Synthesis, Proteolytic Pathways, and Proteome in Human Duodenal Mucosa. Am. J. Clin. Nutr. 2011, 94, 784–794.
- Bowtell, J.L.; Leese, G.P.; Smith, K.; Watt, P.W.; Nevill, A.; Rooyackers, O.; Wagenmakers, A.J.; Rennie, M.J. Effect of Oral Glucose on Leucine Turnover in Human Subjects at Rest and during Exercise at Two Levels of Dietary Protein. J. Physiol. 2000, 525 Pt 1, 271–281.
- Thomsen, H.H.; Rittig, N.; Johannsen, M.; Møller, A.B.; Jørgensen, J.O.; Jessen, N.; Møller, N. Effects of 3-Hydroxybutyrate and Free Fatty Acids on Muscle Protein Kinetics and Signaling during LPS-Induced Inflammation in Humans: Anticatabolic Impact of Ketone Bodies. Am. J. Clin. Nutr. 2018, 108, 857–867.
- Rodriguez, N.; Schwenk, W.F.; Beaufrere, B.; Miles, J.M.; Haymond, M.W. Trioctanoin Infusion Increases in Vivo Leucine Oxidation: A Lesson in Isotope Modeling. Am. J. Physiol. -Endocrinol. Metab. 1986, 251, E343–E348.
- Haymond, M.W.; Tessari, P.; Beaufrere, B.; Rodriguez, N.; Bailey, J.; Miles, J.M. Effects of Parenteral Lipid on Leucine Metabolism: Dependence of Fatty Acid Chain Length. JPEN J. Parenter. Enter. Nutr. 1988, 12, 94S–97S.
- Lang, C.H. Elevated Plasma Free Fatty Acids Decrease Basal Protein Synthesis, but Not the Anabolic Effect of Leucine, in Skeletal Muscle. Am. J. Physiol. -Endocrinol. Metab. 2006, 291, E666–E674.
- Tsintzas, K.; Jones, R.; Pabla, P.; Mallinson, J.; Barrett, D.A.; Kim, D.-H.; Cooper, S.; Davies, A.; Taylor, T.; Chee, C.; et al. Effect of Acute and Short-Term Dietary Fat Ingestion on Postprandial Skeletal Muscle Protein Synthesis Rates in Middle-Aged, Overweight, and Obese Men. Am. J. Physiol. -Endocrinol. Metab. 2020, 318, E417–E429.
- Furst, T.; Massaro, A.; Miller, C.; Williams, B.T.; LaMacchia, Z.M.; Horvath, P.J. β-Alanine Supplementation Increased Physical Performance and Improved Executive Function Following Endurance Exercise in Middle Aged Individuals. J. Int. Soc. Sports Nutr. 2018, 15, 32.
- Artioli, G.G.; Gualano, B.; Smith, A.; Stout, J.; Lancha, A.H. Role of β-Alanine Supplementation on Muscle Carnosine and Exercise Performance. Med. Sci. Sports Exerc. 2010, 42, 1162–1173.
- Parise, G.; Mihic, S.; MacLennan, D.; Yarasheski, K.E.; Tarnopolsky, M.A. Effects of Acute Creatine Monohydrate Supplementation on Leucine Kinetics and Mixed-Muscle Protein Synthesis. J. Appl. Physiol. 2001, 91, 1041–1047.
- Tipton, K.D.; Ferrando, A.A. Improving Muscle Mass: Response of Muscle Metabolism to Exercise, Nutrition and Anabolic Agents. Essays Biochem. 2008, 44, 85–98.
- Tessari, P.; Inchiostro, S.; Biolo, G.; Marescotti, M.C.; Fantin, G.; Boscarato, M.T.; Merola, G.; Mantero, F.; Tiengo, A. Leucine Kinetics and the Effects of Hyperinsulinemia in Patients With Cushing’s Syndrome. J. Clin. Endocrinol. Metab. 1989, 68, 256–262.
- Horber, F.F.; Haymond, M.W. Human Growth Hormone Prevents the Protein Catabolic Side Effects of Prednisone in Humans. J. Clin. Investig. 1990, 86, 265–272.
- Schakman, O.; Kalista, S.; Barbé, C.; Loumaye, A.; Thissen, J.P. Glucocorticoid-Induced Skeletal Muscle Atrophy. Int. J. Biochem. Cell Biol. 2013, 45, 2163–2172.
- Louard, R.J.; Bhushan, R.; Gelfand, R.A.; Barrett, E.J.; Sherwin, R.S. Glucocorticoids Antagonize Insulin’s Antiproteolytic Action on Skeletal Muscle in Humans. J. Clin. Endocrinol. Metab. 1994, 79, 278–284.
- Fryburg, D.A.; Gelfand, R.A.; Barrett, E.J. Growth Hormone Acutely Stimulates Forearm Muscle Protein Synthesis in Normal Humans. Am. J. Physiol. -Endocrinol. Metab. 1991, 260, E499–E504.
- Haymond, M.W.; Horber, F.; De Feo, P.; Kahn, S.E.; Mauras, N. Effect of Human Growth Hormone and Insulin-Like Growth Factor I on Whole-Body Leucine and Estimates of Protein Metabolism. Horm. Res. 1993, 40, 92–94.
- Kandalla, P.K.; Goldspink, G.; Butler-Browne, G.; Mouly, V. Mechano Growth Factor E Peptide (MGF-E), Derived from an Isoform of IGF-1, Activates Human Muscle Progenitor Cells and Induces an Increase in Their Fusion Potential at Different Ages. Mech. Ageing Dev. 2011, 132, 154–162.
- Górecki, D.C.; Beręsewicz, M.; Zabłocka, B. Neuroprotective Effects of Short Peptides Derived from the Insulin-like Growth Factor 1. Neurochem. Int. 2007, 51, 451–458.
- Matthews, D.E.; Pesola, G.; Campbell, R.G. Effect of epinephrine on amino acid and energy metabolism in humans. Am. J. Physiol. 1990, 258 Pt 1, E948–E956.
- Schiefermeier, M.; Ratheiser, K.; Zauner, C.; Roth, E.; Eichler, H.; Matthews, D. Epinephrine Does Not Impair Utilization of Exogenous Amino Acids in Humans. Am. J. Clin. Nutr. 1997, 65, 1765–1773.
- Kraenzlin, M.E.; Keller, U.; Keller, A.; Thélin, A.; Arnaud, M.J.; Stauffacher, W. Elevation of Plasma Epinephrine Concentrations Inhibits Proteolysis and Leucine Oxidation in Man via Beta-Adrenergic Mechanisms. J. Clin. Investig. 1989, 84, 388–393.
- Yoh, K.; Ikeda, K.; Horie, K.; Inoue, S. Roles of Estrogen, Estrogen Receptors, and Estrogen-Related Receptors in Skeletal Muscle: Regulation of Mitochondrial Function. Int. J. Mol. Sci. 2023, 24, 1853.
- Boland, R.; Vasconsuelo, A.; Milanesi, L.; Ronda, A.C.; de Boland, A.R. 17beta-Estradiol Signaling in Skeletal Muscle Cells and Its Relationship to Apoptosis. Steroids 2008, 73, 859–863.
- Tsai, W.-J.A.; McCormick, K.M.; Brazeau, D.A.; Brazeau, G.A. Estrogen Effects on Skeletal Muscle Insulin-like Growth Factor 1 and Myostatin in Ovariectomized Rats. Exp. Biol. Med. 2007, 232, 1314–1325.
- Cho, E.-J.; Choi, Y.; Jung, S.-J.; Kwak, H.-B. Role of Exercise in Estrogen Deficiency-Induced Sarcopenia. J. Exerc. Rehabil. 2022, 18, 2–9.
- Smith, G.I.; Yoshino, J.; Reeds, D.N.; Bradley, D.; Burrows, R.E.; Heisey, H.D.; Moseley, A.C.; Mittendorfer, B. Testosterone and Progesterone, But Not Estradiol, Stimulate Muscle Protein Synthesis in Postmenopausal Women. J. Clin. Endocrinol. Metab. 2014, 99, 256–265.
- Biolo, G.; Tipton, K.D.; Klein, S.; Wolfe, R.R. An Abundant Supply of Amino Acids Enhances the Metabolic Effect of Exercise on Muscle Protein. Am. J. Physiol. -Endocrinol. Metab. 1997, 273, E122–E129.
- Norton, L.E.; Layman, D.K. Leucine Regulates Translation Initiation of Protein Synthesis in Skeletal Muscle after Exercise. J. Nutr. 2006, 136, 533S–537S.
- Herman, J.R.; Rana, S.R.; Chleboun, G.S.; Gilders, R.M.; Hageman, F.C.; Hikida, R.S.; Kushnick, M.R.; Ragg, K.E.; Staron, R.S.; Toma, K. Correlation Between Muscle Fiber Cross-Sectional Area And Strength Gain Using Three Different Resistance-Training Programs In College-Aged Women. J. Strength Cond. Res. 2010, 24, 1.
- Jones, E.J.; Bishop, P.A.; Woods, A.K.; Green, J.M. Cross-Sectional Area and Muscular Strength: A Brief Review. Sports Med. 2008, 38, 987–994.
- Schoenfeld, B.J. The Mechanisms of Muscle Hypertrophy and Their Application to Resistance Training. J. Strength Cond. Res. 2010, 24, 2857–2872.
- Kumar, V.; Selby, A.; Rankin, D.; Patel, R.; Atherton, P.; Hildebrandt, W.; Williams, J.; Smith, K.; Seynnes, O.; Hiscock, N.; et al. Age-Related Differences in the Dose-Response Relationship of Muscle Protein Synthesis to Resistance Exercise in Young and Old Men: Age-Related Effects of Exercise on Muscle Anabolism. J. Physiol. 2009, 587, 211–217.
- Mayhew, D.L.; Kim, J.; Cross, J.M.; Ferrando, A.A.; Bamman, M.M. Translational Signaling Responses Preceding Resistance Training-Mediated Myofiber Hypertrophy in Young and Old Humans. J. Appl. Physiol. 2009, 107, 1655–1662.
- Phillips, S.M.; Tipton, K.D.; Aarsland, A.; Wolf, S.E.; Wolfe, R.R. Mixed Muscle Protein Synthesis and Breakdown after Resistance Exercise in Humans. Am. J. Physiol. -Endocrinol. Metab. 1997, 273, E99–E107.
- Yarasheski, K.E.; Pak-Loduca, J.; Hasten, D.L.; Obert, K.A.; Brown, M.B.; Sinacore, D.R. Resistance Exercise Training Increases Mixed Muscle Protein Synthesis Rate in Frail Women and Men >/=76 Yr Old. Am. J. Physiol. 1999, 277, E118–E125.
- Yarasheski, K.E.; Zachwieja, J.J.; Bier, D.M. Acute Effects of Resistance Exercise on Muscle Protein Synthesis Rate in Young and Elderly Men and Women. Am. J. Physiol. 1993, 265, E210–E214.
- Sheffield-Moore, M.; Paddon-Jones, D.; Sanford, A.P.; Rosenblatt, J.I.; Matlock, A.G.; Cree, M.G.; Wolfe, R.R. Mixed Muscle and Hepatic Derived Plasma Protein Metabolism Is Differentially Regulated in Older and Younger Men Following Resistance Exercise. Am. J. Physiol. -Endocrinol. Metab. 2005, 288, E922–E929.
- Küüsmaa, M.; Schumann, M.; Sedliak, M.; Kraemer, W.J.; Newton, R.U.; Malinen, J.-P.; Nyman, K.; Häkkinen, A.; Häkkinen, K. Effects of Morning versus Evening Combined Strength and Endurance Training on Physical Performance, Muscle Hypertrophy, and Serum Hormone Concentrations. Appl. Physiol. Nutr. Metab. 2016, 41, 1285–1294.
- Biolo, G.; Williams, B.D.; Fleming, R.Y.; Wolfe, R.R. Insulin Action on Muscle Protein Kinetics and Amino Acid Transport during Recovery after Resistance Exercise. Diabetes 1999, 48, 949–957.

