 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Raffaele Pellegrino | -- | 3063 | 2023-09-27 20:57:42 | | | |
| 2 | Lindsay Dong | Meta information modification | 3063 | 2023-09-28 07:20:26 | | |
Video Upload Options
The melanocortin system is a complex set of molecular mediators and receptors involved in many physiological and homeostatic processes. These include the regulation of melanogenesis, steroidogenesis, neuromodulation and the modulation of inflammatory processes. In the latter context, the system has assumed importance in conditions of chronic digestive inflammation, such as inflammatory bowel diseases (IBD), in which numerous experiences have been accumulated in mouse models of colitis.
1. Introduction
2. α-MSH as a Key Melanocortin in the Modulation of Inflammatory Processes
3. The Receptors of the Melanocortin System: The Basis for Their Role in Peripheral and Systemic Inflammation
3.1. MC1R
3.2. MC2R
3.3. MC3R
3.4. MC4R
3.5. MC5R
4. The Role of the Melanocortin System in IBD
4.1. MC1R Mediates and Improves Intestinal Inflammation in Major Models of Experimental Colitis, and Some of Its Agonists Are Being Carefully Studied as Potential Therapeutic Agents in IBD
4.2. MC2R Is Involved in the Interaction between UVA and UVB and Murine DSS-Induced Colitis
4.3. Colic Expression of MC3R and MC4R Appears to Differ According to Disease Activity in IBD: Initial Experience and Scarce Evidence
Indeed, despite the more central localisation of MC4R, its role and peripheral localisations are increasingly emerging. Panaro et al. [52] showed how MC4R is expressed in different portions of the gastrointestinal tract, such as the stomach, small intestine and descending colon. Several cytotypes, such as small intestine cells positive for cholecystokinin, gastric inhibitory peptide, and glucagon-like peptide 1 of mice, showed high expression of the mRNA coding for this receptor. By stimulating it with α-MSH administration, its functions seem related to a paracrine inhibition of electrolyte secretion.
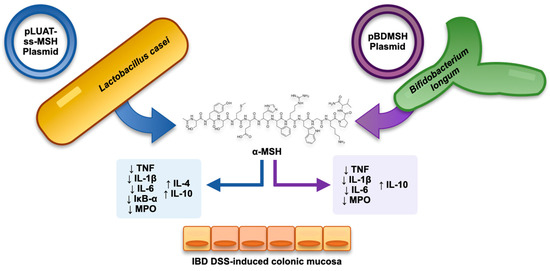
4.4. Combining Recombinant Bacteria and α-MSH as a Strategy in Experimental Colitis
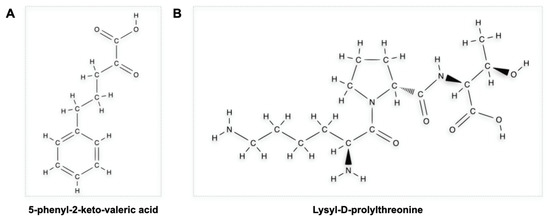
4.5. Exploiting the Similarity with KPV Tripeptide of the C-Terminal End of α-MSH: The Anti-Inflammatory Role of KPV-like Tripeptide KdPT
4.6. Not Only α-MSH: What Potential of β-MSH?
4.7. Prospects
5. Conclusions
References
- Catania, A. The Melanocortin System in Leukocyte Biology. J. Leukoc. Biol. 2007, 81, 383–392.
- Holder, J.R.; Haskell-Luevano, C. Melanocortin Ligands: 30 Years of Structure-Activity Relationship (SAR) Studies. Med. Res. Rev. 2004, 24, 325–356.
- Yang, Y.; Harmon, C.M. Molecular Signatures of Human Melanocortin Receptors for Ligand Binding and Signaling. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2436–2447.
- Videira, I.F.d.S.; Moura, D.F.L.; Magina, S. Mechanisms Regulating Melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83.
- Lam, C.; Getting, S. Melanocortin Receptor Type 3 as a Potential Target for Anti-Inflammatory Therapy. CDTIA 2004, 3, 311–315.
- Kawai, T.; Akira, S. Signaling to NF-KappaB by Toll-like Receptors. Trends Mol. Med. 2007, 13, 460–469.
- Lawrence, T. The Nuclear Factor NF-KappaB Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651.
- Thoma, A.; Lightfoot, A.P. NF-KB and Inflammatory Cytokine Signalling: Role in Skeletal Muscle Atrophy. Adv. Exp. Med. Biol. 2018, 1088, 267–279.
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB System. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241.
- Cheng, Z.; Taylor, B.; Ourthiague, D.R.; Hoffmann, A. Distinct Single-Cell Signaling Characteristics Are Conferred by the MyD88 and TRIF Pathways during TLR4 Activation. Sci. Signal. 2015, 8, ra69.
- Cui, H.-S.; Hayasaka, S.; Zhang, X.-Y.; Chi, Z.-L.; Hayasaka, Y. Effect of Alpha-Melanocyte-Stimulating Hormone on Interleukin 8 and Monocyte Chemotactic Protein 1 Expression in a Human Retinal Pigment Epithelial Cell Line. Ophthalmic Res. 2005, 37, 279–288.
- Haycock, J.W.; Wagner, M.; Morandini, R.; Ghanem, G.; Rennie, I.G.; Mac Neil, S. Alpha-Melanocyte-Stimulating Hormone Inhibits NF-KappaB Activation in Human Melanocytes and Melanoma Cells. J. Investig. Dermatol. 1999, 113, 560–566.
- Buggy, J.J. Binding of Alpha-Melanocyte-Stimulating Hormone to Its G-Protein-Coupled Receptor on B-Lymphocytes Activates the Jak/STAT Pathway. Biochem. J. 1998, 331 Pt 1, 211–216.
- Rogler, G.; Brand, K.; Vogl, D.; Page, S.; Hofmeister, R.; Andus, T.; Knuechel, R.; Baeuerle, P.A.; Schölmerich, J.; Gross, V. Nuclear Factor KappaB Is Activated in Macrophages and Epithelial Cells of Inflamed Intestinal Mucosa. Gastroenterology 1998, 115, 357–369.
- Pallone, F.; Monteleone, G. Mechanisms of Tissue Damage in Inflammatory Bowel Disease. Curr. Opin. Gastroenterol. 2001, 17, 307–312.
- Chang, S.H.; Jung, E.J.; Lim, D.G.; Park, Y.H.; Wee, Y.M.; Kim, J.H.; Kim, Y.H.; Choi, M.Y.; Koo, S.K.; Choi, K.D.; et al. Anti-Inflammatory Action of Alpha-Melanocyte Stimulating Hormone (Alpha-MSH) in Anti-CD3/CD28-Mediated Spleen and CD4(+)CD25(-) T Cells and a Partial Participation of IL-10. Immunol. Lett. 2008, 118, 44–48.
- Nishida, T.; Taylor, A.W. Specific Aqueous Humor Factors Induce Activation of Regulatory T Cells. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2268–2274.
- Taylor, A.W.; Streilein, J.W.; Cousins, S.W. Alpha-Melanocyte-Stimulating Hormone Suppresses Antigen-Stimulated T Cell Production of Gamma-Interferon. Neuroimmunomodulation 1994, 1, 188–194.
- Auriemma, M.; Brzoska, T.; Klenner, L.; Kupas, V.; Goerge, T.; Voskort, M.; Zhao, Z.; Sparwasser, T.; Luger, T.A.; Loser, K. α-MSH-Stimulated Tolerogenic Dendritic Cells Induce Functional Regulatory T Cells and Ameliorate Ongoing Skin Inflammation. J. Investig. Dermatol. 2012, 132, 1814–1824.
- Saporiti, F.; Piacentini, L.; Alfieri, V.; Bono, E.; Ferrari, F.; Chiesa, M.; Colombo, G.I. Melanocortin-1 Receptor Positively Regulates Human Artery Endothelial Cell Migration. Cell. Physiol. Biochem. 2019, 52, 1339–1360.
- Seaton, M.E.; Parent, B.A.; Sood, R.F.; Wurfel, M.M.; Muffley, L.A.; O’Keefe, G.E.; Gibran, N.S. Melanocortin-1 Receptor Polymorphisms and the Risk of Complicated Sepsis After Trauma: A Candidate Gene Association Study. Shock 2017, 47, 79–85.
- Rossi, S.; Maisto, R.; Gesualdo, C.; Trotta, M.C.; Ferraraccio, F.; Kaneva, M.K.; Getting, S.J.; Surace, E.; Testa, F.; Simonelli, F.; et al. Corrigendum to “Activation of Melanocortin Receptors MC1 and MC5 Attenuates Retinal Damage in Experimental Diabetic Retinopathy”. Mediat. Inflamm. 2021, 2021, 1–2.
- Maisto, R.; Gesualdo, C.; Trotta, M.C.; Grieco, P.; Testa, F.; Simonelli, F.; Barcia, J.M.; D’Amico, M.; Di Filippo, C.; Rossi, S. Melanocortin Receptor Agonists MCR1-5 Protect Photoreceptors from High-Glucose Damage and Restore Antioxidant Enzymes in Primary Retinal Cell Culture. J. Cell. Mol. Med. 2017, 21, 968–974.
- Andersen, M.; Nagaev, I.; Meyer, M.K.; Nagaeva, O.; Wikberg, J.; Mincheva-Nilsson, L.; Andersen, G.N. Melanocortin 2, 3 and 4 Receptor Gene Expressions Are Downregulated in CD8+ T Cytotoxic Lymphocytes and CD19+ B Lymphocytes in Rheumatoid Arthritis Responding to TNF-α Inhibition. Scand. J. Immunol. 2017, 86, 31–39.
- Getting, S.J.; Perretti, M. MC3-R as a Novel Target for Antiinflammatory Therapy. Drug News Perspect. 2000, 13, 19–27.
- Getting, S.J.; Gibbs, L.; Clark, A.J.; Flower, R.J.; Perretti, M. POMC Gene-Derived Peptides Activate Melanocortin Type 3 Receptor on Murine Macrophages, Suppress Cytokine Release, and Inhibit Neutrophil Migration in Acute Experimental Inflammation. J. Immunol. 1999, 162, 7446–7453.
- Getting, S.J.; Di Filippo, C.; Christian, H.C.; Lam, C.W.; Rossi, F.; D’Amico, M.; Perretti, M. MC-3 Receptor and the Inflammatory Mechanisms Activated in Acute Myocardial Infarct. J. Leukoc. Biol. 2004, 76, 845–853.
- Catania, A. Neuroprotective Actions of Melanocortins: A Therapeutic Opportunity. Trends Neurosci. 2008, 31, 353–360.
- Mountjoy, K.G.; Mortrud, M.T.; Low, M.J.; Simerly, R.B.; Cone, R.D. Localization of the Melanocortin-4 Receptor (MC4-R) in Neuroendocrine and Autonomic Control Circuits in the Brain. Mol. Endocrinol. 1994, 8, 1298–1308.
- Tracey, K.J. Physiology and Immunology of the Cholinergic Antiinflammatory Pathway. J. Clin. Investig. 2007, 117, 289–296.
- Zila, I.; Mokra, D.; Kopincova, J.; Kolomaznik, M.; Javorka, M.; Calkovska, A. Vagal-Immune Interactions Involved in Cholinergic Anti-Inflammatory Pathway. Physiol. Res. 2017, 66, S139–S145.
- Giuliani, D.; Ottani, A.; Altavilla, D.; Bazzani, C.; Squadrito, F.; Guarini, S. Melanocortins and the Cholinergic Anti-Inflammatory Pathway. Adv. Exp. Med. Biol. 2010, 681, 71–87.
- Tracey, K.J. The Inflammatory Reflex. Nature 2002, 420, 853–859.
- Kamermans, A.; Verhoeven, T.; van Het Hof, B.; Koning, J.J.; Borghuis, L.; Witte, M.; van Horssen, J.; de Vries, H.E.; Rijnsburger, M. Setmelanotide, a Novel, Selective Melanocortin Receptor-4 Agonist Exerts Anti-Inflammatory Actions in Astrocytes and Promotes an Anti-Inflammatory Macrophage Phenotype. Front. Immunol. 2019, 10, 2312.
- Maisto, R.; Oltra, M.; Vidal-Gil, L.; Martínez-Gil, N.; Sancho-Pellúz, J.; Filippo, C.D.; Rossi, S.; D’Amico, M.; Barcia, J.M.; Romero, F.J. ARPE-19-Derived VEGF-Containing Exosomes Promote Neovascularization in HUVEC: The Role of the Melanocortin Receptor 5. Cell Cycle 2019, 18, 413–424.
- Hugot, J.P.; Chamaillard, M.; Zouali, H.; Lesage, S.; Cézard, J.P.; Belaiche, J.; Almer, S.; Tysk, C.; O’Morain, C.A.; Gassull, M.; et al. Association of NOD2 Leucine-Rich Repeat Variants with Susceptibility to Crohn’s Disease. Nature 2001, 411, 599–603.
- Ogura, Y.; Bonen, D.K.; Inohara, N.; Nicolae, D.L.; Chen, F.F.; Ramos, R.; Britton, H.; Moran, T.; Karaliuskas, R.; Duerr, R.H.; et al. A Frameshift Mutation in NOD2 Associated with Susceptibility to Crohn’s Disease. Nature 2001, 411, 603–606.
- Maaser, C. Crucial Role of the Melanocortin Receptor MC1R in Experimental Colitis. Gut 2006, 55, 1415–1422.
- Kannengiesser, K.; Maaser, C.; Heidemann, J.; Luegering, A.; Ross, M.; Brzoska, T.; Bohm, M.; Luger, T.A.; Domschke, W.; Kucharzik, T. Melanocortin-Derived Tripeptide KPV Has Anti-Inflammatory Potential in Murine Models of Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2008, 14, 324–331.
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2023 Update. Nucleic Acids Res. 2023, 51, D1373–D1380.
- Luger, T.A.; Scholzen, T.E.; Brzoska, T.; Böhm, M. New Insights into the Functions of Alpha-MSH and Related Peptides in the Immune System. Ann. N. Y. Acad. Sci. 2003, 994, 133–140.
- Xiao, B.; Xu, Z.; Viennois, E.; Zhang, Y.; Zhang, Z.; Zhang, M.; Han, M.K.; Kang, Y.; Merlin, D. Orally Targeted Delivery of Tripeptide KPV via Hyaluronic Acid-Functionalized Nanoparticles Efficiently Alleviates Ulcerative Colitis. Mol. Ther. 2017, 25, 1628–1640.
- Laroui, H.; Dalmasso, G.; Nguyen, H.T.T.; Yan, Y.; Sitaraman, S.V.; Merlin, D. Drug-Loaded Nanoparticles Targeted to the Colon with Polysaccharide Hydrogel Reduce Colitis in a Mouse Model. Gastroenterology 2010, 138, 843–853.
- Dalmasso, G.; Charrier-Hisamuddin, L.; Nguyen, H.T.T.; Yan, Y.; Sitaraman, S.; Merlin, D. PepT1-Mediated Tripeptide KPV Uptake Reduces Intestinal Inflammation. Gastroenterology 2008, 134, 166–178.
- Steel, A.; Nussberger, S.; Romero, M.F.; Boron, W.F.; Boyd, C.A.; Hediger, M.A. Stoichiometry and PH Dependence of the Rabbit Proton-Dependent Oligopeptide Transporter PepT1. J. Physiol. 1997, 498 Pt 3, 563–569.
- Shen, H.; Smith, D.E.; Yang, T.; Huang, Y.G.; Schnermann, J.B.; Brosius, F.C. Localization of PEPT1 and PEPT2 Proton-Coupled Oligopeptide Transporter MRNA and Protein in Rat Kidney. Am. J. Physiol. 1999, 276, F658–F665.
- Daniel, H. Molecular and Integrative Physiology of Intestinal Peptide Transport. Annu. Rev. Physiol. 2004, 66, 361–384.
- Merlin, D.; Si-Tahar, M.; Sitaraman, S.V.; Eastburn, K.; Williams, I.; Liu, X.; Hediger, M.A.; Madara, J.L. Colonic Epithelial HPepT1 Expression Occurs in Inflammatory Bowel Disease: Transport of Bacterial Peptides Influences Expression of MHC Class 1 Molecules. Gastroenterology 2001, 120, 1666–1679.
- Spana, C.; Taylor, A.W.; Yee, D.G.; Makhlina, M.; Yang, W.; Dodd, J. Probing the Role of Melanocortin Type 1 Receptor Agonists in Diverse Immunological Diseases. Front. Pharmacol. 2019, 9, 1535.
- Dodd, J.; Jordan, R.; Makhlina, M.; Barnett, K.; Roffel, A.; Spana, C.; Obr, A.; Dhingra, P.; Kayne, P.S. A Novel Oral Formulation of the Melanocortin-1 Receptor Agonist PL8177 Resolves Inflammation in Preclinical Studies of Inflammatory Bowel Disease and Is Gut Restricted in Rats, Dogs, and Humans. Front. Immunol. 2023, 14, 1083333.
- Hiramoto, K.; Yamate, Y.; Sato, E.F. The Effects of Ultraviolet Eye Irradiation on Dextran Sodium Sulfate-Induced Ulcerative Colitis in Mice. Photochem. Photobiol. 2016, 92, 728–734.
- Panaro, B.L.; Tough, I.R.; Engelstoft, M.S.; Matthews, R.T.; Digby, G.J.; Møller, C.L.; Svendsen, B.; Gribble, F.; Reimann, F.; Holst, J.J.; et al. The Melanocortin-4 Receptor Is Expressed in Enteroendocrine L Cells and Regulates the Release of Peptide YY and Glucagon-like Peptide 1 In Vivo. Cell Metab. 2014, 20, 1018–1029.
- Wei, P.; Yang, Y.; Liu, Z.; Huang, J.; Gong, Y.; Sun, H. Oral Bifidobacterium Longum Expressing Alpha-Melanocyte-Stimulating Hormone to Fight Experimental Colitis. Drug Deliv. 2016, 23, 2058–2064.
- Wei, P.; Yang, Y.; Ding, Q.; Li, X.; Sun, H.; Liu, Z.; Huang, J.; Gong, Y. Oral Delivery of Bifidobacterium Longum Expressing α-Melanocyte-Stimulating Hormone to Combat Ulcerative Colitis. J. Med. Microbiol. 2016, 65, 160–168.
- Bettenworth, D.; Buyse, M.; Böhm, M.; Mennigen, R.; Czorniak, I.; Kannengiesser, K.; Brzoska, T.; Luger, T.A.; Kucharzik, T.; Domschke, W.; et al. The Tripeptide KdPT Protects from Intestinal Inflammation and Maintains Intestinal Barrier Function. Am. J. Pathol. 2011, 179, 1230–1242.
- Spencer, J.D.; Schallreuter, K.U. Regulation of Pigmentation in Human Epidermal Melanocytes by Functional High-Affinity Beta-Melanocyte-Stimulating Hormone/Melanocortin-4 Receptor Signaling. Endocrinology 2009, 150, 1250–1258.
- Luger, T.A.; Scholzen, T.; Brzoska, T.; Becher, E.; Slominski, A.; Paus, R. Cutaneous Immunomodulation and Coordination of Skin Stress Responses by Alpha-Melanocyte-Stimulating Hormone. Ann. N. Y. Acad. Sci. 1998, 840, 381–394.
- Bradamante, M.; Turčić, P.; Stambuk, N.; Konjevoda, P.; Aralica, G.; Alerić, I.; Kozmar, A. Cytoprotective Effects of β-Melanocortin in the Rat Gastrointestinal Tract. Molecules 2012, 17, 11680–11692.
- Yeo, G.S.H.; Chao, D.H.M.; Siegert, A.-M.; Koerperich, Z.M.; Ericson, M.D.; Simonds, S.E.; Larson, C.M.; Luquet, S.; Clarke, I.; Sharma, S.; et al. The Melanocortin Pathway and Energy Homeostasis: From Discovery to Obesity Therapy. Mol. Metab. 2021, 48, 101206.

