Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Satoru Matsuda | -- | 1800 | 2023-09-07 15:51:12 | | | |
| 2 | Peter Tang | Meta information modification | 1800 | 2023-09-08 04:45:41 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Kitagishi, Y.; Matsuda, S. Function of RUFY Family Proteins with RUN Domain. Encyclopedia. Available online: https://encyclopedia.pub/entry/48934 (accessed on 29 July 2026).
Kitagishi Y, Matsuda S. Function of RUFY Family Proteins with RUN Domain. Encyclopedia. Available at: https://encyclopedia.pub/entry/48934. Accessed July 29, 2026.
Kitagishi, Yasuko, Satoru Matsuda. "Function of RUFY Family Proteins with RUN Domain" Encyclopedia, https://encyclopedia.pub/entry/48934 (accessed July 29, 2026).
Kitagishi, Y., & Matsuda, S. (2023, September 07). Function of RUFY Family Proteins with RUN Domain. In Encyclopedia. https://encyclopedia.pub/entry/48934
Kitagishi, Yasuko and Satoru Matsuda. "Function of RUFY Family Proteins with RUN Domain." Encyclopedia. Web. 07 September, 2023.
Copy Citation
Cell survival, homeostasis and cell polarity rely on the control of membrane trafficking pathways. The RUN domain (comprised of the RPIP8, UNC-14, and NESCA proteins) has been suggested to be implicated in small GTPase-mediated membrane trafficking and cell polarity. Accumulating evidence supports the hypothesis that the RUN domain-containing proteins might be responsible for an interaction with a filamentous network linked to actin cytoskeleton and/or microtubules. In addition, several downstream molecules of PI3K are involved in regulation of the membrane trafficking by interacting with vesicle-associated RUN proteins such as RUFY family proteins.
RUFY family
RUN domain
membrane trafficking
small GTPase
NESCA
Rab
Rap
1. Introduction
Asymmetric organization of cellular components and structures is called cell polarity. Establishment of the cell polarity involves many processes including signaling cascade [1], membrane trafficking events, [2] and cytoskeletal dynamics [3], which are implicated in differentiation, proliferation and morphogenesis of various cellular organisms [4]. Cell polarization is required in order for a cell to function properly. For example, the presence of axon in neuronal cells determines the directional flow of the signal (Figure 1). It is well known that initial trafficking of down-regulated protein is performed by clathrin-mediated internalization and sorting into the lumen of the endosomes [5] (Figure 2). Dysregulation of cell polarity can cause developmental disorder and cancer [6]. Studies have revealed links between cell polarity establishment and cellular membrane traffic [2][7], and the function of endosome-associated proteins has been implicated in cell polarity [8].
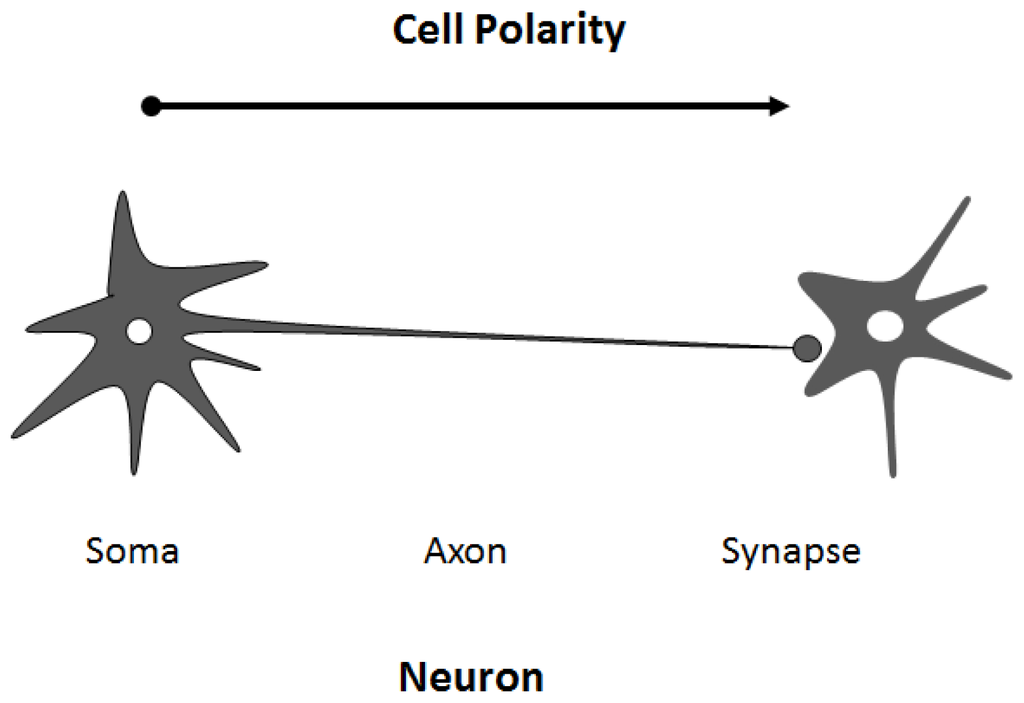
Figure 1. Cell polarity is observed in many cases. For example, neuronal cells are asymmetric with discrete regions responsible for different roles that may underlie the generation of specific compartments within cells, which is distinct in biochemical composition, function, and structure. An arrow indicates the direction of polarity.
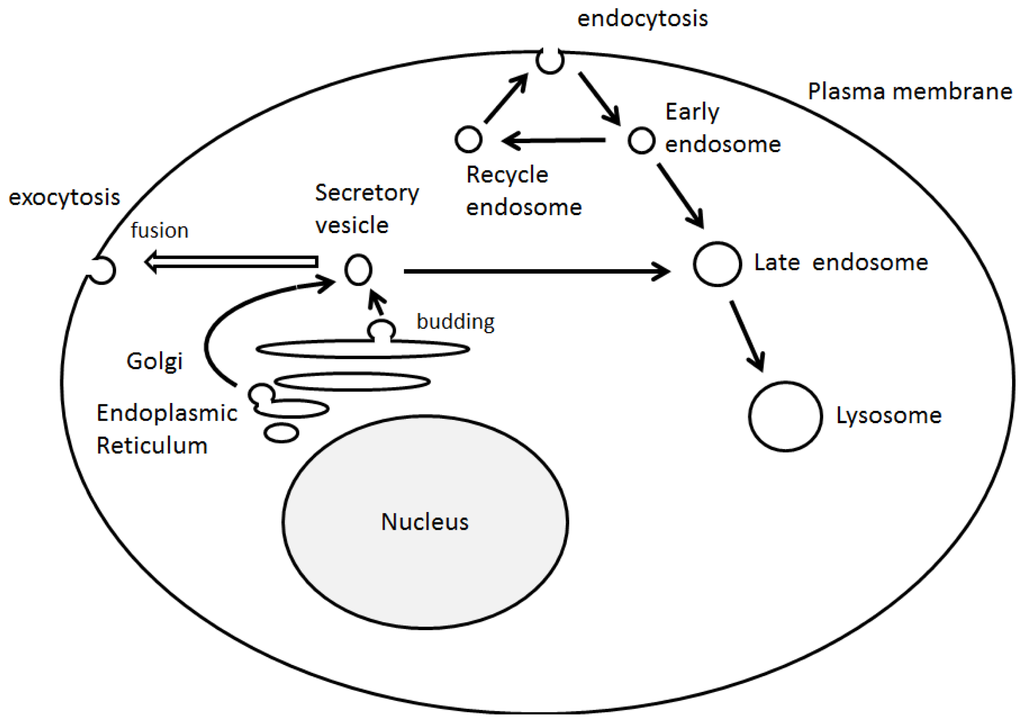
Figure 2. Schematic illustration of intracellular vesicle transport. The model shows several pathways used by biosynthetic secretory and internalized endocytotic cargoes to reach their destinations. Note that some critical trafficking routes have been omitted for clarity.
2. RUN Domain Binds Several Signaling Molecules
The RUN domains, named after RPIP8, UNC-14, and NESCA proteins [9], might function as effectors of the small GTPase superfamily [10][11]. Because many RUN domain-containing proteins are involved in the signaling of small GTPases, including members of the Rap and Rab family, the RUN domain has been suggested to be involved in membrane trafficking [11]. The RUN domain encloses hydrophobic amino acids in the conserved positions, which is a conserved protein motif that consists of approximately 200 amino acids with binding activity to small GTP-binding proteins [9]. The sequence analysis has predicted that the RUN domain is composed of several conserved blocks, which constitute the core of a globular structure. The overall crystal structure of the RUN domain adopts a single globular fold consisting of eight alpha-helices [12]. However, it is not the only function of the RUN domain to bind a GTPase. The RUN domain binds to some molecules, including motor proteins, and the RUN domain might be responsible for an interaction with a filamentous network [11]. NESCA, a signaling adapter protein in the NGF-mediated signaling pathway, contains a RUN domain at the N-terminus [10]. The RUN domain of NESCA comprises nine helices, resembling the other RUN domain-containing proteins [13]. Mutational analyses have demonstrated that the RUN domain is an important structural determinant for the nuclear translocation of NESCA and that the nuclear redistribution of NESCA is essential to its neurite outgrowth-promoting properties [10]. However, the RUN domain of NESCA has different surface electrostatic distributions at the putative GTPase-interacting interface compared to the other RUN domains [13]. The RUN domain of NESCA can bind H-Ras, a downstream signaling molecule of TrkA, as well as TrkA itself, suggesting that the NESCA participates in the NGF-TrkA signaling pathway [13].
The RUN domain-containing proteins have been shown to promote endosomal fusion and are important for vesicular transport. In addition, the RUN domains appear to be required for localization to detergent-insoluble endosomal microdomains [10][11]. The physical interaction between RUN proteins and filamentous materials has been confirmed by several biochemical experiments using wild type and mutant proteins [11]. The association among small GTPases, RUN proteins, and motor proteins might reflect a novel function for these proteins in the transport of vesicular cargoes in cells. It has been reported that FYCO1 functions as an adapter linking autophagosomes to microtubule molecular motors and the Rab7, which is implicated in the phagosomal transport and fusion [14]. Kinesin-1 is a heterotetramer composed of kinesin heavy chain and kinesin light chain. UNC-14, a RUN domain protein binds to the kinesin-1 and regulates synaptic vesicle localization [15]. UNC-14 is also predicted to play an important role in multiple Ras-like GTPase signaling pathways [16]. Because RUN domains are often found in proteins involved in the regulation of Rab family small GTPases, the RUN domain has been suggested to be involved in the Rab-mediated membrane trafficking. It seems there is a common function underlying the mechanism for association of RUN domain to small GTPases and motor proteins.
3. Function of RUFY Family Proteins with the RUN Domain
The RUFY, designated as the RUN and FYVE domain-containing protein family, contains an amino-terminal RUN domain, and a carboxyl-terminal FYVE domain, which associate with phosphatidylinositol 3-phosphate in membranes of early endosomes [17]. Actually, RUFY proteins are localized predominantly to the early endosomes. RUFY proteins are often tyrosine-phosphorylated and the mutant lacking the phosphorylation sites fails to go to the endosomes [17]. It has been suggested Rab10, Rab11, Rab14, and RUFY proteins play an important role in the Glut4 trafficking in adipocytes and in skeletal muscle [18]. Sequence and genome analysis has revealed that a RUFY family consists of 4 members of proteins (Figure 3).
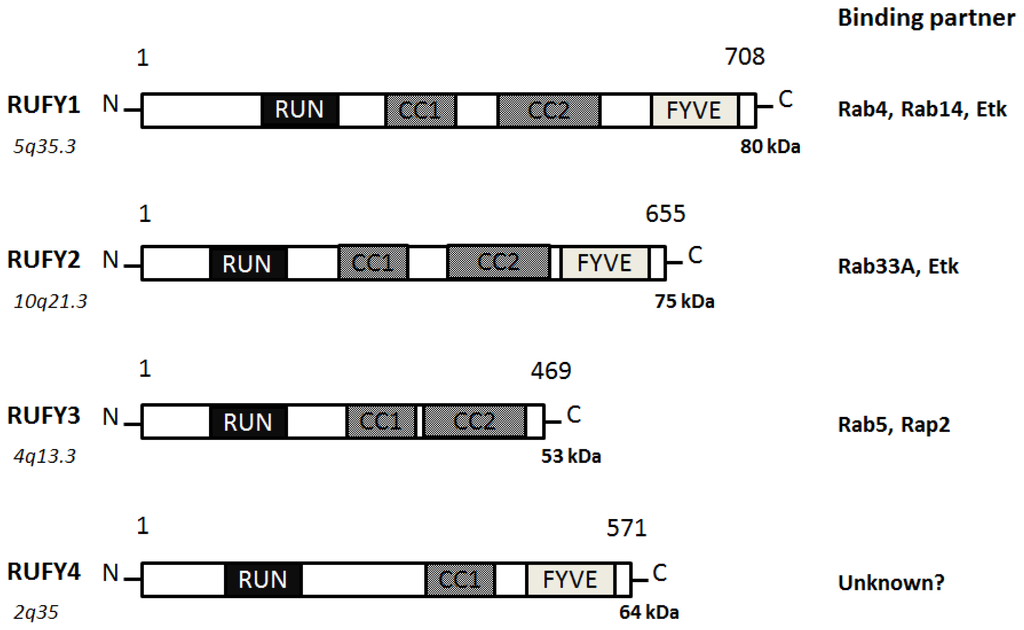
Figure 3. Schematic diagram indicating the domain structures of the RUFY1, RUFY2, RUFY3, and RUFY4 proteins. The functionally important sites and their interaction proteins are shown. Genomic locations of each the genes and approximate molecular mass of each the proteins are also shown at the both ends. RUN = RPIP8, UNC-14, and NESCA proteins, FYVE = Fab-1, YGL023, Vps27, and EEA1 proteins, CC = Coiled Coil.
RUFY1, also known as RABIP4 or ZFYVE12, is a 708-amino acids protein that localizes to the cytoplasm and early endosome membrane [17][19]. RUFY1 has been identified as downstream effector of Etk protein kinase, which is highly expressed in testis, lung, brain and kidney. RUFY1 functions to bind PIP3-containing phospholipid vesicles and participates in early endosomal membrane trafficking [17]. The downstream effects of PI3K signaling are mediated by proteins containing a PIP3-binding module indicated by the FYVE finger domain. Many FYVE domain-containing proteins are localized at the endosomes and play an important role in endocytosis [20]. Through the SH3 and SH2 domains of Etk, the Etk interacts with RUFY1, then phosphorylates Tyr-281 and Tyr-292 of RUFY1, which is essential for the endosomal localization [17]. The Etk plays an important role in the regulation of endocytosis as a downstream effector of PI3K. Two coiled coil domains also determine endosomal localization of RUFY1 [21]. The PI3K inhibitor wortmannin blocks the endosomal localization of RUFY1 [21]. Rab14 engages in a GTP-dependent interaction with RUFY1 [22]. The active Rab14 regulates RUFY1 recruitment onto endosomal membranes, and Rab4 allows endosomal fusion [22]. The Rab14 seems to be the primary determinant of RUFY1 recruitment to the endosomes, and the FYVE domain may assist RUFY1 targeting to PIP3-enriched early endosomes [22]. Both Rab14 and RUFY1 are involved in Rab4-dependent recycling endosome, and enlargement of the early endosomes mediated by RUFY1 needs the interaction with Rab4 [23]. The RUFY1 is present in the sorting endosomes, while Rab4 is present both on sorting and recycling endosomes [24]. This would provide directional trafficking away from the recycling endosomes to the sorting endosomes. RUFY1 can also modify the kinetic parameters of Glut1 protein recycling [24].
RUFY2, also known as RABIP4R or ZFYVE13, contains a RUN domain and a carboxy terminal FYVE zinc finger, separated by two coiled coil domains [25]. Localizing to nucleus, RUFY2 is expressed in brain, lung and testis. RUFY2 as well as RUFY1 interacts with the Etk that is a tyrosine kinase involved in regulation of various cellular processes [17]. The carboxyl domain of RUFY2 binds to a negative form of Rab33A [26]. RUFY3, also known as RIPX or SINGAR1, is diffusely localized in hippocampal neurons and accumulated in the growth cones and axons [27]. RUFY3 ensures the robustness of neuronal polarity by suppressing formation of surplus axons. RUFY3 also contains the RUN domain and seems to play important roles in multiple Ras-like GTPase signaling pathways. Rab5 engages in a GTP-dependent interaction with RUFY3. RUFY3 can bind to the active Rab5 and weakly associates to Rap2 [28]. RUFY3 may function as a docking protein for distinct two small GTPases. It has been reported that oxidized LDL-containing immune complexes affect the gene expression of RUFY3 in human U937 monocytic cells [29]. RUFY4 is a 571-amino acids protein that contains a RUN domain and a FYVE zinc finger domain [30]. RUFY4 is also believed to be involved with zinc ion binding. As poorly characterized for RUFY2 and for RUFY4, little is known about the precise intracellular functions for these proteins.
4. Regulation of Cell Polarity and Membrane Trafficking
Cell polarity and vesicle sorting are significant processes that influence normal cell function including cell adhesion, migration, and neurotransmission. Endosomal membrane trafficking is a spatiotemporally regulated process that confirms suitable delivery of cargo via the pathway. Endosomes can bud inwardly from the membranes to form vesicles, which receive cargo from the cell surface via endocytosis and biosynthetic cargo from the late Golgi complex [31]. The endocytic trafficking has been shown to be a critical component of many signaling pathways, which is indispensable for a wide range of developmental processes [32]. In addition, the endosomal trafficking is regulated by sequential recruitment of a variety of cytosolic and membrane-bound proteins [33]. Studies have elucidated many of the key components of the events that take place during the membrane trafficking. For example, small GTPases of Rab family, its effectors, Ca2+ levels, and phosphoinositides may be all important. Previous studies have demonstrated that Ca2+ influx triggers the final step in exocytotic membrane fusion events during neurotransmission [34].
Similarly, phosphoinositides have been shown to be important for the recruitment of Rab family proteins and the binding factors [35]. Plasma membrane channels are known to be regulated by the Rab proteins [36] and phosphoinositides, which are molecules that determine the vesicular identity and direction of membrane trafficking. In particular, PIP3 is essential for the membrane trafficking of early endosome [37]. The PIP3 has lots of effector proteins in mammalian cells, all of which contain PIP3-binding motifs such as FYVE and/or PH domains [38]. Selective recruitment of these effectors by PIP3 may provide a mechanism by which the directionality for incoming vesicles and endosomes may be established. The PIP3 is thus critical for the maturation of endosomes, and for fusion events with intracellular organelles [39]. Correspondingly, disruption of PIP3 synthesis by wortmannin, a PI3K inhibitor, affects the formation of internal vesicles and the maturation of endosomes [40]. While phosphoinositides can recruit phosphoinositide-binding proteins to regulate the activity of GTPases, the GTPases can, in turn, control the activity of PIP-metabolizing enzymes.
References
- Happé, H.; de Heer, E.; Peters, D.J. Polycystic kidney disease: The complexity of planar cell polarity and signaling during tissue regeneration and cyst formation. Biochim. Biophys. Acta 2011, 1812, 1249–1255.
- Santiago-Tirado, F.H.; Bretscher, A. Membrane-trafficking sorting hubs: Cooperation between PI4P and small GTPases at the trans-Golgi network. Trends Cell Biol 2011, 21, 515–525.
- Baum, B.; Georgiou, M. Dynamics of adherens junctions in epithelial establishment, maintenance, and remodeling. J. Cell Biol 2011, 192, 907–917.
- Wu, G.; Ge, J.; Huang, X.; Hua, Y.; Mu, D. Planar cell polarity signaling pathway in congenital heart diseases. J. Biomed. Biotechnol 2011, 2011, 589414.
- Traub, L.M. Tickets to ride: Selecting cargo for clathrin-regulated internalization. Nat. Rev. Mol. Cell Biol 2009, 10, 583–596.
- Martin-Belmonte, F.; Perez-Moreno, M. Epithelial cell polarity, stem cells and cancer. Nat. Rev. Cancer 2011, 12, 23–38.
- Layton, A.; Savage, N.S.; Howell, A.S.; Carroll, S.Y.; Drubin, D.G.; Lew, D.J. Modeling vesicle traffic reveals unexpected consequences for Cdc42p-mediated polarity establishment. Curr. Biol 2011, 21, 184–194.
- Golachowska, M.R.; Hoekstra, D.; van IJzendoorn, S.C. Recycling endosomes in apical plasma membrane domain formation and epithelial cell polarity. Trends Cell Biol 2010, 20, 618–626.
- Callebaut, I.; de Gunzburg, J.; Goud, B.; Mornon, J.P. RUN domains: A new family of domains involved in Ras-like GTPase signaling. Trends Biochem. Sci 2001, 26, 79–83.
- MacDonald, J.I.; Kubu, C.J.; Meakin, S.O. Nesca, a novel adapter, translocates to the nuclear envelope and regulates neurotrophin-induced neurite outgrowth. Cell Biol 2004, 164, 851–862.
- Yoshida, H.; Kitagishi, Y.; Okumura, N.; Murakami, M.; Nishimura, Y.; Matsuda, S. How do you RUN on? FEBS Lett 2011, 585, 1707–1710.
- Kukimoto-Niino, M.; Takagi, T.; Akasaka, R.; Murayama, K.; Uchikubo-Kamo, T.; Terada, T.; Inoue, M.; Watanabe, S.; Tanaka, A.; Hayashizaki, Y.; et al. Crystal structure of the RUN domain of the RAP2-interacting protein x. J. Biol. Chem 2006, 281, 31843–31853.
- Sun, Q.; Han, C.; Liu, L.; Wang, Y.; Deng, H.; Bai, L.; Jiang, T. Crystal structure and functional implication of the RUN domain of human NESCA. Protein Cell 2012, 3, 609–617.
- Pankiv, S.; Alemu, E.A.; Brech, A.; Bruun, J.A.; Lamark, T.; Overvatn, A.; Bjørkøy, G.; Johansen, T. FYCO1 is a Rab7 effector that binds to LC3 and PI3P to mediate microtubule plus end-directed vesicle transport. J. Cell Biol 2010, 188, 253–269.
- Sakamoto, R.; Byrd, D.T.; Brown, H.M.; Hisamoto, N.; Matsumoto, K.; Jin, Y. The Caenorhabditis elegans UNC-14 RUN domain protein binds to the kinesin-1 and UNC-16 complex and regulates synaptic vesicle localization. Mol. Biol. Cell 2005, 16, 483–496.
- Ogura, K.; Goshima, Y. The autophagy-related kinase UNC-51 and its binding partner UNC-14 regulate the subcellular localization of the Netrin receptor UNC-5 in Caenorhabditis elegans. Development 2006, 133, 3441–3450.
- Yang, J.; Kim, O.; Wu, J.; Qiu, Y. Interaction between tyrosine kinase Etk and a RUN domain- and FYVE domain-containing protein RUFY1. A possible role of ETK in regulation of vesicle trafficking. J. Biol. Chem 2002, 277, 30219–30226.
- Larance, M.; Ramm, G.; Stöckli, J.; van Dam, E.M.; Winata, S.; Wasinger, V.; Simpson, F.; Graham, M.; Junutula, J.R.; Guilhaus, M.; James, D.E. Characterization of the role of the Rab GTPase-activating protein AS160 in insulin-regulated GLUT4 trafficking. J. Biol. Chem 2005, 280, 37803–37813.
- Fouraux, M.A.; Deneka, M.; Ivan, V.; van der Heijden, A.; Raymackers, J.; van Suylekom, D.; van Venrooij, W.J.; van der Sluijs, P.; Pruijn, G.J. Rabip4′ is an effector of rab5 and rab4 and regulates transport through early endosomes. Mol. Biol. Cell 2004, 15, 611–624.
- Simonsen, A.; Wurmser, A.E.; Emr, S.D.; Stenmark, H. The role of phosphoinositides in membrane transport. Curr. Opin. Cell Biol 2001, 13, 485–492.
- Mari, M.; Macia, E.; le Marchand-Brustel, Y.; Cormont, M. Role of the FYVE finger and the RUN domain for the subcellular localization of Rabip4. J. Biol. Chem 2001, 276, 42501–42508.
- Yamamoto, H.; Koga, H.; Katoh, Y.; Takahashi, S.; Nakayama, K.; Shin, H.W. Functional cross-talk between Rab14 and Rab4 through a dual effector, RUFY1/Rabip4. Mol. Biol. Cell 2010, 21, 2746–2755.
- Mari, M.; Monzo, P.; Kaddai, V.; Keslair, F.; Gonzalez, T.; le Marchand-Brustel, Y.; Cormont, M. The Rab4 effector Rabip4 plays a role in the endocytotic trafficking of Glut 4 in 3T3-L1 adipocytes. J. Cell Sci 2006, 119, 1297–1306.
- Cormont, M.; Mari, M.; Galmiche, A.; Hofman, P.; le Marchand-Brustel, Y. A FYVE-finger-containing protein, Rabip4, is a Rab4 effector involved in early endosomal traffic. Proc. Natl. Acad. Sci. USA 2001, 98, 1637–1642.
- Barbe, L.; Lundberg, E.; Oksvold, P.; Stenius, A.; Lewin, E.; Björling, E.; Asplund, A.; Pontén, F.; Brismar, H.; Uhlén, M.; Andersson-Svahn, H. Toward a confocal subcellular atlas of the human proteome. Mol. Cell Proteomics 2008, 7, 499–508.
- Fukuda, M.; Kobayashi, H.; Ishibashi, K.; Ohbayashi, N. Genome-wide investigation of the Rab binding activity of RUN domains: Development of a novel tool that specifically traps GTP-Rab35. Cell Struct. Funct 2011, 36, 155–170.
- Mori, T.; Wada, T.; Suzuki, T.; Kubota, Y.; Inagaki, N. Singar1, a novel RUN domain-containing protein, suppresses formation of surplus axons for neuronal polarity. J. Biol. Chem 2007, 282, 19884–19893.
- Yoshida, H.; Okumura, N.; Kitagishi, Y.; Shirafuji, N.; Matsuda, S. Rab5(Q79L) interacts with the carboxyl terminus of RUFY3. Int. J. Biol. Sci 2010, 6, 187–189.
- Hammad, S.M.; Twal, W.O.; Barth, J.L.; Smith, K.J.; Saad, A.F.; Virella, G.; Argraves, W.S.; Lopes-Virella, M.F. Oxidized LDL immune complexes and oxidized LDL differentially affect the expression of genes involved with inflammation and survival in human U937 monocytic cells. Atherosclerosis 2009, 202, 394–404.
- Kimura, K.; Wakamatsu, A.; Suzuki, Y.; Ota, T.; Nishikawa, T.; Yamashita, R.; Yamamoto, J.; Sekine, M.; Tsuritani, K.; Wakaguri, H.; et al. Diversification of transcriptional modulation: Large-scale identification and characterization of putative alternative promoters of human genes. Genome Res 2006, 16, 55–65.
- Pfeffer, S.R. Multiple routes of protein transport from endosomes to the trans Golgi network. FEBS Lett 2009, 583, 3811–3816.
- González-Gaitán, M. Endocytic trafficking during Drosophila development. Mech. Dev 2003, 120, 1265–1282.
- Bakhru, S.H.; Altiok, E.; Highley, C.; Delubac, D.; Suhan, J.; Hitchens, T.K.; Ho, C.; Zappe, S. Enhanced cellular uptake and long-term retention of chitosan-modified iron-oxide nanoparticles for MRI-based cell tracking. Int. J. Nanomed 2012, 7, 4613–4623.
- Monck, J.R.; Fernandez, J.M. The exocytotic fusion pore and neurotransmitter release. Neuron 1994, 12, 707–716.
- Zaid, H.; Antonescu, C.N.; Randhawa, V.K.; Klip, A. Insulin action on glucose transporters through molecular switches, tracks and tethers. Biochem. J 2008, 413, 201–215.
- Saxena, S.K.; Kaur, S. Regulation of epithelial ion channels by Rab GTPases. Biochem. Biophys. Res. Commun 2006, 351, 582–587.
- Kanamarlapudi, V. Centaurin-alpha1 and KIF13B kinesin motor protein interaction in ARF6 signalling. Biochem. Soc. Trans 2005, 33, 1279–1281.
- Saito, K.; Tautz, L.; Mustelin, T. The lipid-binding SEC14 domain. Biochim. Biophys. Acta 2007, 1771, 719–726.
- Stenmark, H.; Gillooly, D.J. Intracellular trafficking and turnover of phosphatidylinositol 3-phosphate. Semin. Cell Dev. Biol 2001, 12, 193–199.
- Shisheva, A. Phosphoinositides in insulin action on GLUT4 dynamics: Not just PtdIns(3,4,5)P3. Am. J. Physiol. Endocrinol. Metab 2008, 295, E536–E544.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
571
Revisions:
2 times
(View History)
Update Date:
08 Sep 2023
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No