Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Satoru Matsuda | -- | 2228 | 2023-08-21 18:24:25 | | | |
| 2 | Dean Liu | Meta information modification | 2228 | 2023-08-22 02:35:48 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Suga, N.; Ikeda, Y.; Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Matsuda, S. N6-Methyladenosine in Epitranscriptome, Autophagy and Neurodegenerative Diseases. Encyclopedia. Available online: https://encyclopedia.pub/entry/48289 (accessed on 29 July 2026).
Suga N, Ikeda Y, Yoshikawa S, Taniguchi K, Sawamura H, Matsuda S. N6-Methyladenosine in Epitranscriptome, Autophagy and Neurodegenerative Diseases. Encyclopedia. Available at: https://encyclopedia.pub/entry/48289. Accessed July 29, 2026.
Suga, Naoko, Yuka Ikeda, Sayuri Yoshikawa, Kurumi Taniguchi, Haruka Sawamura, Satoru Matsuda. "N6-Methyladenosine in Epitranscriptome, Autophagy and Neurodegenerative Diseases" Encyclopedia, https://encyclopedia.pub/entry/48289 (accessed July 29, 2026).
Suga, N., Ikeda, Y., Yoshikawa, S., Taniguchi, K., Sawamura, H., & Matsuda, S. (2023, August 21). N6-Methyladenosine in Epitranscriptome, Autophagy and Neurodegenerative Diseases. In Encyclopedia. https://encyclopedia.pub/entry/48289
Suga, Naoko, et al. "N6-Methyladenosine in Epitranscriptome, Autophagy and Neurodegenerative Diseases." Encyclopedia. Web. 21 August, 2023.
Copy Citation
Changes in epitranscriptome with N6-methyladenine (m6A) modification could be involved in the development of multiple diseases, which might be a prevalent modification of messenger RNAs (mRNAs) in eukaryotes. The m6A modification might be performed through the action of methyltransferases, demethylases, and methylation-binding proteins. Importantly, the m6A methylation may be associated with various neurological disorders including Alzheimer’s disease (AD), Parkinson’s disease (PD), depression, aging-related diseases, and/or aging itself. In addition, the m6A methylation might functionally regulate the eukaryotic transcriptome by influencing the splicing, export, subcellular localization, translation, stability, and decay of mRNAs.
N6-methyladenine
reactive oxygen species
RNA-binding protein
1. Introduction
Neurodegenerative diseases are interlaced with the disorders of multicellular function and/or interaction in the central nervous system (CNS) [1]. “Neurodegeneration” represents a condition where neuronal cells gradually lose their potential functions, and eventually perish. Alzheimer’s disease (AD) is the most common neurodegenerative disease. Neurodegenerative diseases have a huge impact on patients themselves, adding to the social economic burden [2], which is regrettably expected to increase worldwide [3]. Now, progress in resolving the mechanisms behind several neurodegenerative diseases has been considerably recognized. For example, epigenetic modifications and genetic risk factors may be implicated in their appearance, and their mixture could predict or support the identification of individuals at significant risk [4]. In general, neurodegenerative diseases could result in motor dysfunctions and behavioral manifestations such as ataxia, and/or dementia [5]. Clarification of pathogenic mechanisms and new targeted drugs have been urgently needed.
The most important advances in RNA-modification-mediated regulation of gene expression might be an evolving field of epitranscriptomics. The epitranscriptome may contain all the biochemical modifications of the RNA within cells, which is dynamically regulated by specific enzymatic reactions. These RNA modifications could regulate a variety of physiological RNA functions. More than hundred chemical alterations of RNAs have been found in all types of RNAs [6]. The most notable RNA methylation may account for more than 60% of all RNA alterations. Among them, the N6-methyladenosine (m6A) might be a noticeable post-transcriptional RNA modification in eukaryotes, which could regulate the expression of various genes [7][8]. It is noteworthy that the m6A is highly enriched in adult brain tissue [9], which might indicate that it plays a critical role in neurogenesis, neurodevelopment, and/or neurodegenerative disorders [10]. In addition, the profusion of m6A in the brain seems to gradually increase with age from newborns, and reaches a peak in later life [11]. Within the structure of total RNAs, the m6A is mostly dispersed in the 3’ untranslated region (UTR), coding sequence of message RNA (mRNA), and/or regions nearby the stop codon [12]. The roles of the m6A modification in the control of gene expression may be strictly connected to various normal and/or pathological routes containing DNA damage response, cellular differentiation, and/or the occurrence of neurodegenerative diseases. In addition to the regulation of the transcriptional mRNA, m6A modification could also regulate the transcription of a variety of non-coding RNAs (ncRNAs) including microRNAs (miRNAs), circular RNAs (circRNAs), and/or long non-coding RNAs (lncRNAs) [13]. Therefore, m6A modification could take part in the regulation of various processes in physiological cellular activities including cell development, embryonic development and/or stress responses, as well as in a regulatory role of several diseases [14][15]. The motif of m6A during the development may diverge across tissues and/or developmental courses [16], in which the m6A modification may be inclined to the coding sequence and/or 3′UTR [16]. Remarkably, even a single modification could have a vast effect on the function of RNAs. Interestingly, ncRNAs such as lncRNAs could also regulate the expression of m6A-related proteins [17].
Studies have suggested that m6A modification on mRNAs could influence the proliferation and/or differentiation of neural progenitor cells in a comprehensive understanding of RNA methylation-based diagnosis and/or therapies for neurodegenerative diseases [18][19]. In addition, many studies have also demonstrated the effects of m6A modification in the autophagy mechanisms [20], suggesting that the m6A change might play a critical role in controlling the development of neurodegenerative diseases via the control of autophagy. For example, the m6A modification could instruct direct inhibitory effects on autophagy [21]. Moreover, the m6A modification could influence the construction of autophagosomes to dysregulate autophagy [22]. Occasionally, the m6A modification could even initiate and/or promote autophagy [23]. Interestingly, the properties of the m6A modification for autophagy may be reliant on disease situation and/or stages. Accordingly, abnormal m6A methylation could modify biological processes and/or regulate various human disorders [24]. Therefore, the roles of m6A modifying enzymes might be involved in the pathological process of neurodegenerative diseases, which might be of huge importance to the development of a novel tactic for specific therapy of neurodegenerative diseases. In this report, researchers will go over the main points of the latest studies describing the impact on m6A modification in neurodegenerative diseases via the regulation of autophagy, and discuss the role of the m6A modification–autophagy relationship for the promising treatment of neurodegenerative diseases (Figure 1).
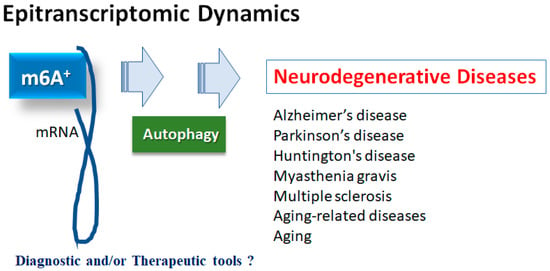
Figure 1. Illustration of the relation of m6A RNAs methylation to various neurodegenerative diseases. Roles of m6A RNAs methylation with autophagy have been proposed in several neurodegenerative diseases including Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, myasthenia gravis and multiple sclerosis, as well as in aging-related diseases and aging. Consequently, m6A RNAs methylation could be diagnostic and/or therapeutic tools for these diseases.
2. m6A Methylation of mRNAs in Neurodegenerative Diseases
2.1. Alzheimer’s Disease
The m6A abnormality is directly related to AD [25], which may be one of the leading reasons of dementia and an increasing health problem worldwide. There is still no cure for AD. AD may be characterized by synaptic loss, hyper-phosphorylation of tau, extracellular plaques of amyloid-beta accumulation, and various levels of neuro-inflammation, as in neurodegenerative diseases [26]. Several biological processes may be exaggerated by epitranscriptomic modifications, which could control the expression of specific genes via the metabolism of mRNAs. In response to modifications of certain mRNAs by m6A methylation, the nervous system could become dysfunctional, which might play an important role in the development of AD. In general, the N6-methyladenosine levels are high in the brain [27]. Correspondingly, the expression of multiple m6A methylation regulators could change when AD occurs. In particular, m6A methylation levels are significantly elevated in the cerebral cortex and/or hippocampus of AD model mice, possibly due to the elevated expression of the m6A methyltransferase and/or the decreased expression of the m6A demethylase, which might be associated with the membrane synaptic growth [25][28]. It has been reported that the methyltransferase-like 3 (METTL3) may be downregulated at the hippocampus in human AD samples, and upregulation of METTL3 could promote autophagic p-Tau clearance and ameliorate AD both in vitro and in vivo [29]. In addition, the m6A could control protein levels of key genes involved in AD-associated pathways [27][30].
2.2. Parkinson’s Disease
Parkinson’s disease (PD) might be the second most-frequent neurodegenerative disease after AD. It has been shown that multiple m6A methylated proteins are associated with PD. The m6A modifications could also regulate dopaminergic signaling. For example, the deletion of METTL14 in the substantia nigra could reduce the level of m6A mRNAs, and might impair the autonomic activity in mice [31]. Overexpression of the fat mass and obesity-associated (FTO) gene and m6A inhibitor cycloleucine could reduce m6A levels in PD models [32]. It has been shown that manganese exposure is a major environmental cause of PD. Remarkably, FTO overexpression could also diminish the manganese-induced cytotoxicity, and thereby could improve the symptoms of PD [33]. Accordingly, it is suggested that some inhibitors of FTO might be utilized for the treatment of PD. For example, entacapone is a catechol-O-methyltransferase inhibitor approved as an adjunctive therapy in combination with levodopa for PD treatment which could promote the modification of the target gene forkhead box protein O1 (FOXO1)-m6A by decreasing FTO expression [34].
2.3. Neurodegenerative Disorders and Aging
Huntington’s disease is an autosomal dominant neurodegenerative disorder, which is characterized by choreiform movements, cognitive deficits, and/or psychiatric symptoms [35]. These impairments may be attributed to hippocampal dysfunction and/or corticostriatal dysfunction [36]. Altered m6A RNA methylation as a unique hallmark of Huntington’s disease might contribute to hippocampal memory deficits in model mice of Huntington’s disease [37]. Huntington’s disease is an autoimmune disease of the nervous system mainly caused by the autoantibodies against the acetylcholine receptor (AChR) in the postsynaptic membranes of the neuromuscular junction [38]. It has been suggested that myasthenia gravis may be a complex multifactorial disease involving environmental, genetic, and/or immunological factors. Among them, the m6A regulators might play imperative roles in the pathogenesis of myasthenia gravis [39]. Multiple sclerosis is a neuro-inflammatory disease that affects the brain, spinal cord, and optic nerves [40]. Unfortunately, there is also no permanent cure for this neurological disorder. Optimal treatment regimens should be agreed upon for a good quality of life (QOL) [41]. The m6A RNA modifications might be closely associated with the development of multiple sclerosis [42]. It is noteworthy that cellular senescence is an important component of the aging process. It has been suggested that aging could definitely affect m6A RNAs methylation in the hippocampal regions [43]. In addition, there are aging-related differences in the modification level of the epigenome m6A [44]. Additionally, the reversibility of the m6A modification in aging may indicate its possibility for delaying aging.
3. ROS, Inflammation and m6A mRNAs
The m6A modification is the most dynamic and reversible epigenetic modification of eukaryotic mRNAs [45], which could be regulated by methyltransferases and/or demethylases [46]. Methyltransferases, also called “writers”, may be composed of METTL3, methyltransferase-like 14 (METTL14), and/or Wilms tumor 1-associated protein (WTAP), which could catalyze the methylation of N6-adenosine with the function of methyl connection [47]. The METTL3 might be the catalytic subunit, while the METTL14 may be involved in the stability of the complex, as well as the RNA recruitment. The WTAP may also be required for the recruitment of mRNA. Among them, the METTL3 has been described as playing imperative roles in many pathological processes, predominantly in inflammatory and/or autoimmune responses [47]. m6A-RNAs binding proteins, also called “readers”, may generally contain heterogeneous nuclear ribonucleoprotein C (HNRNPC), YTH domain-containing protein 1/2 (YTHDC1/2), insulin-like growth factor 2 mRNA-binding protein 1/2/3 (IGF2BP1/2/3), and/or YTH domain-containing family protein 1/2/3 (YTHDF1/2/3). Reader molecules might be prepared for distinguishing the m6A motif, thereby accomplishing the modification of function on the m6A-RNA [48]. Additionally, the m6A modification could be detached by demethylases, also known as “erasers”, including alkB homolog 5 (ALKBH5) and/or FTO (Figure 2). For example, the FTO could favorably demethylate the m6A located nearer to the mRNA cap.
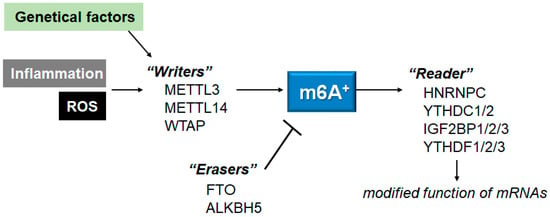
Figure 2. Schematic representation of the players involved in the m6A RNAs methylation. The m6A modification is regulated by methyltransferases “writers” and demethylases “erasers”. The m6A-RNAs binding proteins are called “readers”. Example molecules are also shown for each player. Genetic factors and/or inflammation with ROS may affect the function of these players. The arrowhead means stimulation, and the hammerhead represents inhibition. Note that some critical pathways have been omitted, for clarity.
For example, the half-life of myosin heavy chain 3 (Myh3) mRNA could be significantly decreased after METTL3 deletion, in which Myh3 might have several potential m6A modification sites. Downregulation of METTL3 could reverse the lipopolysaccharide (LPS)-induced myocardial cell damage, largely by increasing Myh3 mRNA stability [49]. In addition, the deletion of METTL3 could also increase the expression of myeloid differentiation factor 88 (MyD88), which may prevent the activation of NF-κB signaling in the LPS-treated cells [50]. Similarly, the silencing of FTO could suppress the proliferation and/or invasion of cervical cancer cells via the m6A modification of the myelocytomatosis oncogene (Myc) and the zinc finger E-box-binding homeobox 1 (ZEB1) [51]. The silencing of FTO could also inhibit the NLRP3-mediated IL-1β expression through the modification of the NF-κB signaling [52], which may be related to the protection of cells.
Methylation put in by the “writers” could be reversed by “erasers” [53], which might be controlled for healthy cellular homeostasis. Therefore, dysregulations of m6A might be linked to the perturbations of cell death and/or proliferation in diverse diseases [54]. Accordingly, m6A modifications are potential novel diagnostic and/or therapeutic targets for several diseases, including neurodegenerative disease [55]. As mentioned above, the m6A modification may be vigorously controlled by RNA methyltransferases (also known as “writers”), RNA demethylases (“erasers”) and RNA-binding proteins (“readers”) [56]. Interestingly, the m6A level of RNAs may be significantly increased with the treatment of LPS [57]. In addition, METTL3 expression might also be upregulated via the increased ROS production [58]. For example, the ROS content might be elevated with the concomitant increase in m6A methylation in the liver after LPS treatment [59]. Therefore, the m6A methylation of RNAs could be modulated under oxidative stresses [59]. Furthermore, ROS could significantly increase the expression of YTHDF2, with the associated elevation of m6A methylation. Gathering this evidence has suggested that the alteration of the m6A modification may be a prevalent phenomenon under oxidative stress conditions. Therefore, immunological and/or inflammatory stresses could affect the m6A level of mRNAs [60]. In general, ROS could regulate major epigenetic processes in various cells [61][62]. In particular, the change in ROS levels might contribute to the m6A RNA methylation [63]. Consequently, the m6A methylation may be related to the increased levels of ROS content.
References
- Wang, M.; Pan, W.; Xu, Y.; Zhang, J.; Wan, J.; Jiang, H. Microglia-mediated neuroinflammation: A potential target for the treatment of cardiovascular diseases. J. Inflamm. Res. 2022, 15, 3083–3094.
- Martinez-Martin, P.; Rodriguez-Blazquez, C.; Forjaz, M.J. Quality of life and burden in caregivers for patients with Parkinson’s disease: Concepts, assessment and related factors. Expert Rev. Pharmacoecon. Outcomes Res. 2012, 12, 221–230.
- Brodaty, H.; Breteler, M.M.; DeKosky, S.T.; Dorenlot, P.; Fratiglioni, L.; Hock, C.; Kenigsberg, P.A.; Scheltens, P.; De Strooper, B. The world of dementia beyond 2020. J. Am. Geriatr. Soc. 2011, 59, 923–927.
- Do, M.D.; Tran, T.N.; Luong, A.B.; Le, L.H.G.; Van Le, T.; Le, K.T.; Van Vo, N.T.; Le, T.N.N.; Vu, H.A.; Mai, T.P.; et al. Clinical and genetic analysis of Vietnamese patients diagnosed with early-onset Parkinson’s disease. Brain Behav. 2023, 13, e2950.
- Naseri, N.; Sharma, M.; Velinov, M. Autosomal dominant neuronal ceroid lipofuscinosis: Clinical features and molecular basis. Clin. Genet. 2021, 99, 111–118.
- Cantara, W.A.; Crain, P.F.; Rozenski, J.; McCloskey, J.A.; Harris, K.A.; Zhang, X.; Vendeix, F.A.; Fabris, D.; Agris, P.F. The RNA modification database, RNAMDB: 2011 update. Nucleic Acids Res. 2010, 39 (Suppl. S1), D195–D201.
- Li, Y.; Xiao, J.; Bai, J.; Tian, Y.; Qu, Y.; Chen, X.; Wang, Q.; Li, X.; Zhang, Y.; Xu, J. Molecular characterization and clinical relevance of m6A regulators across 33 cancer types. J. Mol. Cancer 2019, 18, 137.
- Ren, J.; Li, Y.; Wuermanbieke, S.; Hu, S.; Huang, G. N6-methyladenosine (m6A) methyltransferase METTL3-mediated LINC00680 accelerates osteoarthritis through m6A/SIRT1 manner. Cell Death Discov. 2022, 8, 240.
- Weng, Y.-L.; Wang, X.; An, R.; Cassin, J.; Vissers, C.; Liu, Y.; Liu, Y.; Xu, T.; Wang, X.; Wong, S.Z.H.; et al. Epitranscriptomic m6A regulation of axon regeneration in the adult mammalian nervous system. Neuron 2018, 97, 313–325.e316.
- Widagdo, J.; Anggono, V. The m6A-epitranscriptomic signature in neurobiology: From neurodevelopment to brain plasticity. J. Neurochem. 2018, 147, 137–152.
- Livneh, I.; Moshitch-Moshkovitz, S.; Amariglio, N.; Rechavi, G.; Dominissini, D. The m6A epitranscriptome: Transcriptome plasticity in brain development and function. Nat. Rev. Neurosci. 2020, 21, 36–51.
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206.
- Lu, S.; Ding, X.; Wang, Y.; Hu, X.; Sun, T.; Wei, M.; Wang, X.; Wu, H. The relationship between the network of non-coding RNAs-molecular targets and N6-methyladenosine modification in colorectal cancer. Front. Cell Dev. Biol. 2021, 9, 772542.
- Ma, S.; Chen, C.; Ji, X.; Liu, J.; Zhou, Q.; Wang, G.; Yuan, W.; Kan, Q.; Sun, Z. The interplay between m6A RNA methylation and noncoding RNA in cancer. J. Hematol. Oncol. 2019, 12, 121.
- Zhao, B.S.; Roundtree, I.A.; He, C. Post-transcriptional gene regulation by mRNA modifications. Nat. Rev. Mol. Cell Biol. 2017, 18, 31–42.
- Zhang, H.; Shi, X.; Huang, T.; Zhao, X.; Chen, W.; Gu, N.; Zhang, R. Dynamic landscape and evolution of m6A methylation in human. Nucleic Acids Res. 2020, 48, 6251–6264.
- Wang, W.; Dong, D.; Yu, P.; Chen, T.; Gao, R.; Wei, J.; Mo, Z.; Zhou, H.; Yang, Q.; Yue, C.; et al. Prognostic model based on m6A -associated lncRNAs in esophageal cancer. Front. Endocrinol. 2022, 13, 947708.
- Zhou, H.; Wang, B.; Sun, H.; Xu, X.; Wang, Y. Epigenetic regulations in neural stem cells and neurological diseases. Stem Cells Int. 2018, 2018, 6087143.
- Li, J.; Yang, X.; Qi, Z.; Sang, Y.; Liu, Y.; Xu, B.; Liu, W.; Xu, Z.; Deng, Y. The role of mRNA m6A methylation in the nervous system. Cell Biosci. 2019, 9, 66.
- Liu, S.; Li, Q.; Chen, K.; Zhang, Q.; Li, G.; Zhuo, L.; Zhai, B.; Sui, X.; Hu, X.; Xie, T. The emerging molecular mechanism of m6A modulators in tumorigenesis and cancer progression. Biomed. Pharmacother. 2020, 127, 110098.
- Song, H.; Feng, X.; Zhang, H.; Luo, Y.; Huang, J.; Lin, M.; Jin, J.; Ding, X.; Wu, S.; Huang, H.; et al. METTL3 and ALKBH5 oppositely regulate m6A modification of TFEB mRNA, which dictates the fate of hypoxia/reoxygenation-treated cardiomyocytes. Autophagy 2019, 15, 1419–1437.
- Wang, X.; Wu, R.; Liu, Y.; Zhao, Y.; Bi, Z.; Yao, Y.; Liu, Q.; Shi, H.; Wang, F.; Wang, Y. m6A mRNA methylation controls autophagy and adipogenesis by targeting Atg5 and Atg7. Autophagy 2020, 16, 1221–1235.
- Liu, S.; Li, Q.; Li, G.; Zhang, Q.; Zhuo, L.; Han, X.; Zhang, M.; Chen, X.; Pan, T.; Yan, L.; et al. The mechanism of m6A methyltransferase METTL3-mediated autophagy in reversing gefitinib resistance in NSCLC cells by β-elemene. Cell Death Dis. 2020, 11, 969.
- Li, G.; Luo, R.; Zhang, W.; He, S.; Wang, B.; Liang, H.; Song, Y.; Ke, W.; Shi, Y.; Feng, X.; et al. m6A hypomethylation of DNMT3B regulated by ALKBH5 promotes intervertebral disc degeneration via E4F1 deficiency. Clin. Transl. Med. 2022, 12, e765.
- Han, M.; Liu, Z.; Xu, Y.; Liu, X.; Wang, D.; Li, F.; Wang, Y.; Bi, J. Abnormality of m6A mRNA methylation is involved in Alzheimer’s disease. J. Front. Neurosci. 2020, 14, 98.
- Pinheiro, L.; Faustino, C. Therapeutic strategies targeting amyloid-β in Alzheimer’s disease. Curr. Alzheimer Res. 2019, 16, 418–452.
- Shafik, A.M.; Zhang, F.; Guo, Z.; Dai, Q.; Pajdzik, K.; Li, Y.; Kang, Y.; Yao, B.; Wu, H.; He, C.; et al. N6-methyladenosine dynamics in neurodevelopment and aging, and its potential role in Alzheimer’s disease. Genome Biol. 2021, 22, 17.
- Shao, N.; Ye, T.; Xuan, W.; Zhang, M.; Chen, Q.; Liu, J.; Zhou, P.; Song, H.; Cai, B. The effects of N 6-methyladenosine RNA methylation on the nervous system. Mol. Cell Biochem. 2023, 1–13.
- Tang, Z.; Cao, J.; Yao, J.; Fan, X.; Zhao, J.; Zhao, M.; Duan, Q.; Han, B.; Duan, S. KDM1A-mediated upregulation of METTL3 ameliorates Alzheimer’s disease via enhancing autophagic clearance of p-Tau through m6A-dependent regulation of STUB1. Free Radic. Biol. Med. 2023, 195, 343–358.
- Hao, X.; Li, Y.; Huang, G.; Zeng, Y. Role of the N6-methyladenosine regulatory factor in reducing the risk of cardiovascular disease: Subtype diagnosis following aerobic exercise-assisted weight loss. Am. J. Transl. Res. 2022, 14, 5363.
- Teng, Y.; Liu, Z.; Chen, X.; Liu, Y.; Geng, F.; Le, W.; Jiang, H.; Yang, L. Conditional deficiency of m6A methyltransferase Mettl14 in substantia nigra alters dopaminergic neuron function. J. Cell. Mol. Med. 2021, 25, 8567–8572.
- Chen, X.; Yu, C.; Guo, M.; Zheng, X.; Ali, S.; Huang, H.; Zhang, L.; Wang, S.; Huang, Y.; Qie, S.; et al. Down-regulation of m6A mRNA methylation is involved in dopaminergic neuronal death. ACS Chem. Neurosci. 2019, 10, 2355–2363.
- Qi, Z.; Wang, S.; Li, J.; Wen, Y.; Cui, R.; Zhang, K.; Liu, Y.; Yang, X.; Zhang, L.; Xu, B.; et al. Protective role of mRNA demethylase FTO on axon guidance molecules of nigro-striatal projection system in manganese-induced parkinsonism. J. Hazard. Mater. 2022, 426, 128099.
- Ondo, K.; Isono, M.; Nakano, M.; Hashiba, S.; Fukami, T.; Nakajima, M. The N6-methyladenosine modification posttranscriptionally regulates hepatic UGT2B7 expression. Biochem. Pharmacol. 2021, 189, 114402.
- Foroud, T.; Siemers, E.; Kleindorfer, D.; Bill, D.J.; Hodes, M.; Norton, J.A.; Conneally, P.M.; Christian, J.C. Cognitive scores in carriers of Huntington’s disease gene compared to noncarriers. Ann. Neurol. 1995, 37, 657–664.
- Harris, K.L.; Armstrong, M.; Swain, R.; Erzinclioglu, S.; Das, T.; Burgess, N.; Barker, R.A.; Mason, S.L. Huntington’s disease patients display progressive deficits in hippocampal-dependent cognition during a task of spatial memory. Cortex 2019, 119, 417–427.
- Pupak, A.; Singh, A.; Sancho-Balsells, A.; Alcalá-Vida, R.; Espina, M.; Giralt, A.; Martí, E.; Ørom, U.A.V.; Ginés, S.; Brito, V. Altered m6A RNA methylation contributes to hippocampal memory deficits in Huntington’s disease mice. Cell. Mol. Life Sci. 2022, 79, 416.
- Gomez, A.M.; Van Den Broeck, J.; Vrolix, K.; Janssen, S.P.; Lemmens, M.A.; Van Der Esch, E.; Duimel, H.; Frederik, P.; Molenaar, P.C.; Martínez-Martínez, P.; et al. Antibody effector mechanisms in myasthenia gravis—Pathogenesis at the neuromuscular junction. Autoimmunity 2010, 43, 353–370.
- Li, S.; Liu, H.; Ruan, Z.; Guo, R.; Sun, C.; Tang, Y.; Huang, X.; Gao, T.; Hao, S.; Li, H.; et al. Landscape analysis of m6A modification regulators related biological functions and immune characteristics in myasthenia gravis. J. Transl. Med. 2023, 21, 166.
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173.
- Dalmau, J. Diagnostic and treatment challenges, a new section in N2. Neurol. Neuroimmunol. Neuroinflamm. 2018, 5, e511.
- Mo, X.-B.; Lei, S.-F.; Qian, Q.-Y.; Guo, Y.-F.; Zhang, Y.-H.; Zhang, H. Integrative analysis revealed potential causal genetic and epigenetic factors for multiple sclerosis. J. Neurol. 2019, 266, 2699–2709.
- Castro-Hernández, R.; Berulava, T.; Metelova, M.; Epple, R.; Peña Centeno, T.; Richter, J.; Kaurani, L.; Pradhan, R.; Sakib, M.S.; Burkhardt, S.; et al. Conserved reduction of m6A RNA modifications during aging and neurodegeneration is linked to changes in synaptic transcripts. Proc. Natl. Acad. Sci. USA 2023, 120, e2204933120.
- Su, X.; Shen, Y.; Jin, Y.; Kim, I.-M.; Weintraub, N.L.; Tang, Y. Aging-associated differences in epitranscriptomic m6A regulation in response to acute cardiac ischemia/reperfusion injury in female mice. Front. Pharmacol. 2021, 12, 654316.
- Chen, Y.-S.; Ouyang, X.-P.; Yu, X.-H.; Novák, P.; Zhou, L.; He, P.-P.; Yin, K. N6-Adenosine methylation (m6A) RNA modification: An emerging role in cardiovascular diseases. J. Cardiovasc. Transl. Res. 2021, 14, 857–872.
- Shi, H.; Wei, J.; He, C. Where, when, and how: Context-dependent functions of RNA methylation writers, readers, and erasers. Mol. Cell 2019, 74, 640–650.
- Luo, J.; Xu, T.; Sun, K. N6-methyladenosine RNA modification in inflammation: Roles, mechanisms, and applications. Front. Cell Dev. Biol. 2021, 9, 670711.
- Mapperley, C.; van de Lagemaat, L.N.; Lawson, H.; Tavosanis, A.; Paris, J.; Campos, J.; Wotherspoon, D.; Durko, J.; Sarapuu, A.; Choe, J.; et al. The mRNA m6A reader YTHDF2 suppresses proinflammatory pathways and sustains hematopoietic stem cell function. J. Exp. Med. 2020, 218, e20200829.
- Gong, C.; Wu, J.; Li, H.; Luo, C.; Ji, G.; Guan, X.; Liu, J.; Wang, M. METTL3 achieves lipopolysaccharide-induced myocardial injury via m6A -dependent stabilization of Myh3 mRNA. Biochim. Biophys. Acta Mol. Cell Res. 2023, 1870, 119503.
- Feng, Z.; Li, Q.; Meng, R.; Yi, B.; Xu, Q. METTL 3 regulates alternative splicing of MyD88 upon the lipopolysaccharide-induced inflammatory response in human dental pulp cells. J. Cell. Mol. Med. 2018, 22, 2558–2568.
- Wang, A.; Jin, C.; Wang, Y.; Yu, J.; Wang, R.; Tian, X. FTO promotes the progression of cervical cancer by regulating the N6-methyladenosine modification of ZEB1 and Myc. Mol. Carcinog. 2023, 62, 1228–1237.
- Luo, J.; Wang, F.; Sun, F.; Yue, T.; Zhou, Q.; Yang, C.; Rong, S.; Yang, P.; Xiong, F.; Yu, Q.; et al. Targeted inhibition of FTO demethylase protects mice against LPS-induced septic shock by suppressing NLRP3 inflammasome. Front. Immunol. 2021, 12, 663295.
- Yue, Y.; Liu, J.; He, C. RNA N6-methyladenosine methylation in post-transcriptional gene expression regulation. Genes Dev. 2015, 29, 1343–1355.
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA modifications in gene expression regulation. Cell 2017, 169, 1187–1200.
- Deng, Y.; Zhu, H.; Xiao, L.; Liu, C.; Liu, Y.-L.; Gao, W. Identification of the function and mechanism of m6A reader IGF2BP2 in Alzheimer’s disease. Aging 2021, 13, 24086.
- Yang, Y.; Hsu, P.J.; Chen, Y.-S.; Yang, Y.-G. Dynamic transcriptomic m6A decoration: Writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018, 28, 616–624.
- Guo, F.; Zhang, Y.; Ma, J.; Yu, Y.; Wang, Q.; Gao, P.; Wang, L.; Xu, Z.; Wei, X.; Jing, M. m6A mRNA Methylation Was Associated With Gene Expression and Lipid Metabolism in Liver of Broilers Under Lipopolysaccharide Stimulation. Front. Genet. 2022, 13, 818357.
- Qi, L.; Wang, Y.; Hu, H.; Li, P.; Hu, H.; Li, Y.; Wang, K.; Zhao, Y.; Feng, M.; Lyu, H.; et al. m6A methyltransferase METTL3 participated in sympathetic neural remodeling post-MI via the TRAF6/NF-κB pathway and ROS production. J. Mol. Cell. Cardiol. 2022, 170, 87–99.
- Zhong, X.; Yu, J.; Frazier, K.; Weng, X.; Li, Y.; Cham, C.M.; Dolan, K.; Zhu, X.; Hubert, N.; Tao, Y.; et al. Circadian clock regulation of hepatic lipid metabolism by modulation of m6A mRNA methylation. Cell Rep. 2018, 25, 1816–1828.e4.
- Han, Y.-C.; Xie, H.-Z.; Lu, B.; Xiang, R.-L.; Zhang, H.-P.; Li, J.-Y.; Zhang, S.-Y. Lipopolysaccharide alters the m6A epitranscriptomic tagging of RNAs in cardiac tissue. Front. Mol. Biosci. 2021, 8, 670160.
- Xu, Q.; Wang, Y.; Chen, Z.; Yue, Y.; Huang, H.; Wu, B.; Liu, Y.; Zhou, D.-X.; Zhao, Y. ROS-stimulated protein lysine acetylation is required for crown root development in rice. J. Adv. Res. 2023, 48, 33–46.
- Wu, Q.; Ni, X. ROS-mediated DNA methylation pattern alterations in carcinogenesis. Curr. Drug Targets 2015, 16, 13–19.
- Xu, M.; Zhuo, R.; Tao, S.; Liang, Y.; Liu, C.; Liu, Q.; Wang, T.; Zhong, X. m6A RNA Methylation Mediates NOD1/NF-kB Signaling Activation in the Liver of Piglets Challenged with Lipopolysaccharide. Antioxidants 2022, 11, 1954.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
421
Revisions:
2 times
(View History)
Update Date:
22 Aug 2023
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No