Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Marcin Łukaszewicz | -- | 3980 | 2023-07-25 15:03:48 | | | |
| 2 | Sirius Huang | Meta information modification | 3980 | 2023-07-26 03:30:35 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Domżał-Kędzia, M.; Ostrowska, M.; Lewińska, A.; Łukaszewicz, M. Properties and Applications of Levan. Encyclopedia. Available online: https://encyclopedia.pub/entry/47259 (accessed on 23 July 2026).
Domżał-Kędzia M, Ostrowska M, Lewińska A, Łukaszewicz M. Properties and Applications of Levan. Encyclopedia. Available at: https://encyclopedia.pub/entry/47259. Accessed July 23, 2026.
Domżał-Kędzia, Marta, Monika Ostrowska, Agnieszka Lewińska, Marcin Łukaszewicz. "Properties and Applications of Levan" Encyclopedia, https://encyclopedia.pub/entry/47259 (accessed July 23, 2026).
Domżał-Kędzia, M., Ostrowska, M., Lewińska, A., & Łukaszewicz, M. (2023, July 25). Properties and Applications of Levan. In Encyclopedia. https://encyclopedia.pub/entry/47259
Domżał-Kędzia, Marta, et al. "Properties and Applications of Levan." Encyclopedia. Web. 25 July, 2023.
Copy Citation
Polysaccharides play a crucial role in medicine, pharmacy, and cosmetology, as well as in the production of biofuels and biomaterials. Among microbial biopolymers, microbial levan, a fructose polysaccharide, holds significant promise due to its high productivity and chemical diversity. Levan exhibits a wide range of properties, including film-forming ability, biodegradability, non-toxicity, self-aggregation, encapsulation, controlled release capacity, water retention, immunomodulatory and prebiotic activity, antimicrobial and anticancer activity, as well as high biocompatibility. These exceptional properties position levan as an attractive candidate for nature-based materials in food production, modern cosmetology, medicine, and pharmacy.
levan
biopolymers
polysaccharide
Bacillus subtilis
1. Biosynthesis of Levan
Levan is a polysaccharide made up of fructose molecules connected by β-2,6-glycosidic bonds in the main chain and β-2,1 in branching (Figure 1). Levan biosynthesis involves the hydrolysis of sucrose into glucose and fructose polymer through the action of the enzyme levansucrase. Levansucrase catalyzes the transfer of fructose units from sucrose to form β-2,6 glycosidic linkages, resulting in the formation of levan. Levan is primarily digested by the enzyme levanase, which breaks down the β-2,6 glycosidic linkages, releasing fructose as the main metabolite. Levan is synthesized from sucrose by various microorganisms, e.g., bacteria Zymomonas mobilis, Erwinia herbicola, B. subtilis or fungi such as Aspergillus sydowii, Aspergillus versicolor, and it is also found in plants [1][2][3]. Bacterial levans often have molecular weights over 500,000 Da, are commonly branched and form compact nanospheres offering a broad spectrum of applications [1][4].
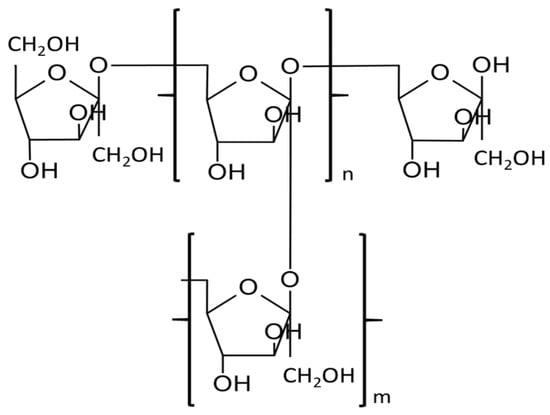
Figure 1. The structural formula of levan.
Levan can be produced by microbial fermentation by many microorganisms exhibiting levansucrase activity (EC 2.4.1.10) and can also be synthesized from sucrose by various levansucrase enzymes in raw, purified, recombinant or immobilized form. To date, levan biosynthesis has been reported using levansucrase from many microorganisms, especially strains of Bacillus, Geobacillus, Lactobacillus and Zymomonas species. Levansucrase is an enzyme responsible for the hydrolysis of sucrose molecules and the catalysis of transfructosylation reactions using either a sucrose molecule or fructooligosaccharide as an acceptor [5][6]. The enzyme is synthesized within the cytoplasm and subsequently accumulates in the periplasm, where it undergoes final conformational changes before secretion [7]. Secretion of levansucrase occurs via the SecA pathway, and its expression level directly impacts the production of levansucrase, indicating a close relationship between the biosynthesis pathway and the secretory pathway [8]. Genes important in the synthesis and hydrolysis of levan are located in two operons: levansucrase and levanase (Figure 2A,B). The gene encoding levansucrase (sacB) is located in a tricistronic operon containing, in addition to the sacB gene, the levB gene (yveB), encoding an enzyme with levanase activity, and the yveA gene, whose function has not yet been determined. yveA is suspected to encode an enzyme with permease activity, due to its similarity to other already-known proteins (Figure 3) [9][10]. The enzyme encoded by the levB gene has (according to the literature) endo-levanase activity, causing the hydrolysis of high molecular weight (HMW) levan molecules to oligosaccharides with a low degree of polymerization (1–10 kDa), down to levanotriosis, levanobiosis and to a small extent fructose molecules [11][12]. It hydrolyzes β-2,6-glycosidic bonds in the polymer, without interfering with β-2,1 bonds in branching [12]. Levanase encoded by levB remains bound to the cell membrane [11].
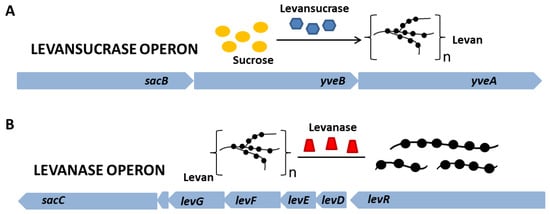
Figure 2. Organization of operons important in the production of levan by B. subtilis.
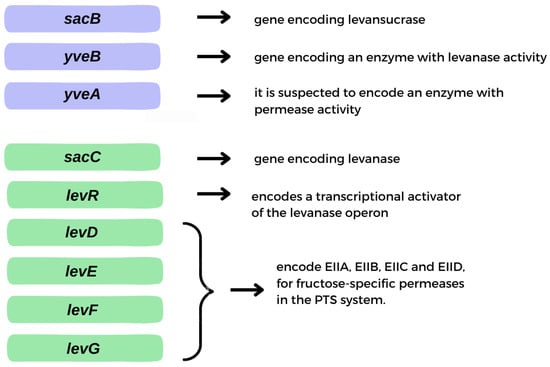
Figure 3. Genes essential in levan synthesis and hydrolysis.
It should be mentioned that levansucrase, in addition to the synthesis activity, also has levanase activity, disrupting β-2,6 bonds, stopping the hydrolysis after reaching β-2,1 bonds in branching [9]. This action targets LMW particles of levan, with a mass no higher than 106 Da [13]. The simultaneous action of enzymes encoded by the sacB and levB genes on levan results in an increased release of fructose molecules. This indicates their joint regulation—the action of levB as endo-levanase stimulates the activity of sacB exo-levanase. Working together, the two proteins mimic the sacC activity relative to levan [14].
The second important operon for levan is the levanase operon. Levanase is encoded by the sacC gene and is located in an operon consisting of five genes—sacC, levD, levE, levF and levG. It has the activity of exo-levanase, degrading levan molecules via hydrolysis of β-2,6 bonds in molecules that have at least three interconnected fructose molecules [9][15]. Its expression is induced by the presence of fructose [16]. The levR gene encodes the transcriptional activator of the levanase operon. Genes levD, levE, levF and levG encode EIIA, EIIB, EIIC and EIID, respectively, fructose-specific permeases in the PTS system (Figure 3). In addition, levE controls levR activity [16][17][18][19][20]. B. subtilis synthesizes levan with bimodal molecular weight distribution—HMW and low molecular weight (LMW) polymer fractions are formed [21]. Their synthesis occurs simultaneously, and the mechanism of this reaction is not yet well understood [22]. One of the hypotheses for the formation of two polymer fractions in one reaction system is the synthesis of HMW by enlarging the previously synthesized LMW levan [21]. Euzenat et al. considered this possibility, also noting the possible low affinity of levansucrase for LMW levan, leading to its accumulation. The slow transfer of several fructosyl residues to the LMW levan molecules eventually lengthens LMW to reach HMW levan [23]. The second hypothesis assumes that the synthesis of HMW and LMW occurs through two different elongation processes. The mechanism of elongation is partly determined by the concentration of levansucrase [21]. High sucrose concentrations favor the synthesis of LMW levan (up to ∼70 kDa), while HMW synthesis may occur at low enzyme concentrations [21]. It is worth noting that levan produced by different organisms has different average molecular weights with varied ranges, as well as the degree of branching, which affects its properties and possible applications [24].
A novel levansucrase-producing levan from Brenneria goodwinii was also characterized. Genomic DNA B. goodwinii, which was recently completed and deposited in the GenBank database, contains the putative gene encoding levansucrase, CPR14579.1. A full-length gene encoding levansucrase was commercially synthesized and cloned into the expression vector pET-22b(+). In this study, recombinant levansucrase B. goodwinii was purified and characterized as a levan polysaccharide. The high concentration of sucrose significantly promoted the polymerization reaction to produce the biopolymer. The purified recombinant enzyme produced 185 g/L levan from 50% (w/v) sucrose at pH 6.0 and 35 °C for 12 h. The molecular weight of the produced levan polysaccharide reached 1.3 × 108 Da, which was much higher than the ones produced by many reported levansucrases [25].
Research by Gao et al. describes the synthesis of levan by Clostridium acetobutylicum. The gene encoding levansucrase (Ca-sacB) consists of 1287 bp and encodes 428 amino acid residues. Interestingly, Ca-sacB was found to have a high product specificity, and fructooligosaccharide was not identified in the product, indicating that Ca-sacB may be valuable in industrial levan production. In addition, Ca-sacB is the first characterized levansucrase isolated from anaerobic bacteria, which should be valuable for the discovery of new enzymatic resources and advancing understanding of the catalytic mechanisms of this enzyme [26].
Synthesis of levan by Leuconostoc mesenteroides has also been demonstrated. Researchers from the Ishida team cloned the gene encoding levansucrase and confirmed that the product of the recombinant enzyme was levan, which is a type of secreted exopolysaccharide that contributes to IgA induction in mice [27].
Kirtle et al. points out that the halophilic bacteria Halomonas smyrnensis can synthesize this polysaccharide. The gene encoding levansucrase (LSC) was cloned into a vector with 6xHis Tag at the C terminus and then expressed in Escherichia coli. The purified recombinant levansucrase produced levan and a wide range of fructooligosaccharides from sucrose, but only in the presence of high salt concentrations (>1.5 M NaCl). Due to its unique biochemical properties, the enzyme from this bacterial strain is important for future research and potential industrial applications in a world facing drought and dwindling freshwater resources [28].
Reports by Lee et al. point to the discovery of levansucrase isolated from Sphingobium chunbukense DJ77. The entire coding sequence of the lsc gene was cloned downstream of the T7 promoter and subsequently purified using the Escherichia coli protein expression system. The results suggest that recombinant E. coli carrying levansucrase from S. chungbukense DJ77 can produce levan under normal growth conditions with less need for pH manipulation [29].
Gluconobacter japonicus LMG 1417, due to its adaptability, genetic tractability and GRAS status, is a promising platform for the industrial production of levan [30]. LevS1417 levansucrase, produced by G. japonicus LMG 1417, and secreted by a signaling peptide-independent pathway, showed exceptionally high activity (4726 ± 821 U/mg at 50 °C). Cell-free levan production based on the supernatant of the tested strain led to a final levan concentration of 157.9 ± 7.6 g/L. The amount of levansucrase secreted was more than doubled by homologous plasmid-mediated overproduction of LevS1417 in G. japonicus LMG 1417. Therefore, the efficiency of cell-free levan production was doubled using a mutant containing a plasmid [30].
Peanibacillus polymyxa can be an extremely cost-effective source for the industrial production of levan exopolysaccharide (EPS) and its functional biomaterials for a wide range of applications, including bioengineering. Liyaskina et al. isolated P. polymyxa from honeycombs and sequenced its genome. Bioinformatic analysis identified the putative levan operon. The sacC and sacB genes were cloned and their products were identified as glycosidic hydrolase and levansucrase, respectively. The infrared spectra obtained through Fourier transform infrared spectroscopy (FT-IR) and the nuclear magnetic resonance (NMR) analysis revealed that the EPS (Exopolysaccharide) is composed of a linear fructan with β-(2→6) glycosidic bonds, specifically known as levan [31].
New endolevanase (evB 2286) from Azotobacter chroococcum DSM 2286, combines extremely high specific activity with beneficial hydrolytic properties. In contrast with endolevanases, LevB2286 produced small amounts of fructose and levan even with significantly prolonged incubation. The combined activity of LevB2286 and levansucrase LevS1417 from Gluconobacter japonicus LMG 1417 led to a one-step synthesis of levan fructooligosaccharide from sucrose [32].
Interestingly, in addition to bacteria, the yeast Saccharomyces cerevisiae, through heterologous expression of levansucrase, also can synthesize levan. Heterologous expression of levansucrase was very difficult to achieve in S. cerevisiae. As a strategy, Franken et al. used null mutant invertase (Δsuc2) and two separate, modified strains of yeast accumulating sucrose as hosts to express M1FT levansucrase, previously cloned from Leuconostoc mesenteroides. Intracellular accumulation of sucrose was achieved either by expressing sucrose synthase (Susy) from potato or spinach sucrose transporter (SUT). The data indicate that in both Δsuc2 and sucrose-hoarding strains, M1FT was able to catalyze fructose polymerization. In the absence of the predicted M1FT secretion signal, intracellular levan accumulation was significantly increased for both strains of sucrose accumulation when they grew on minimal media [33].
Despite numerous years of research, levan synthesis and degradation continue to present intriguing avenues for investigation. The significance of the bimodal mass distribution of levan and the application of fractions with varying molecular weights remains unclear and requires further elucidation. The presence of branching within the levan molecule also poses many unresolved questions. It is yet to be determined whether the synthesis of branched polymers is genetically programmed within the microorganisms or if it is a consequence of enzyme activity and specific external conditions. There are indications that both the levansucrase and levanase operons mutually regulate each other, but the exact mechanisms are yet to be fully understood [22]. Additionally, it remains uncertain whether the presence of these operons in the genetic material of microorganisms guarantees their ability to synthesize levan, or if other factors are necessary for successful levan production. In many scientific articles, there is no exact characterization of the levan, and the structure and subsequent use may depend on the particular microorganism [12].
These challenges provide exciting directions for future research. Further investigations can focus on unraveling the molecular mechanisms underlying levan synthesis and degradation, including the regulatory interactions between relevant operons. Understanding the genetic control and environmental factors influencing levan branching and molecular weight distribution will contribute to a more comprehensive understanding of levan biosynthesis. Additionally, exploring the factors beyond operon presence that influences levan production will aid in optimizing levan synthesis strategies and potentially uncover novel pathways for enhanced levan production. Continued research in these areas will expand our knowledge of levan biology and pave the way for advancements in industrial applications and biotechnological utilization of this intriguing polysaccharide.
2. Microbial Preparation of Levan
Microbial levan is typically obtained through fermentation or enzymatic reactions using isolated enzymes. Sucrose serves as the substrate for the synthesis of this polysaccharide. The optimal sucrose concentration for achieving maximum synthesis efficiency varies not only among different species but also often among different strains of microorganisms, as evident from published data. Despite the various properties and potential uses of microbial polysaccharides, their industrial-scale utilization remains significantly lower compared to laboratory-scale applications. This is primarily due to the low efficiency of biomaterial synthesis, the multi-stage purification process and high production costs. Therefore, further research focusing on process optimization and product purification is crucial (Table 1). Furthermore, the fermentation duration undoubtedly affects the levan yield. Levansucrase, the enzyme involved in the synthesis, is hindered by the presence of residual post-reaction glucose. The accumulation of glucose in the production medium represents a bottleneck in large-scale levan production. Consequently, the microbiological production of this polymer remains limited at present. Commercially available levan is derived from E. herbicola, Z. mobilis or plants [34][35]. However, the limited quantities and high cost of commercial levan hinder its broader application, necessitating the ongoing search for alternatives in microbial production. An essential consideration in industrial levan production is the environmental impact associated with its manufacturing process. One stage of levan preparation involves precipitation using ethanol, methanol or acetone in varying proportions. The use of solvents and their subsequent recovery entail energy demands and have environmental implications. In certain cases, the pH of the solution is adjusted or other treatments are employed to cleanse it from the substrate before polymer precipitation [36][37][38][39]. Another strategy aimed at reducing costs and minimizing environmental impact involves the utilization of substitutes for raw materials required in polymer synthesis, often in the form of by-products derived from other industrial processes. The literature describes various approaches, including the use of sucrose substitutes like molasses or sugar syrups, alternative nitrogen sources or non-sterile production at high salt concentrations (Table 2).
Table 1. Obtaining levan from various microorganisms.
| Scale | Microorganism | Sucrose Concentration [g/L] | Cultivation Conditions | Levan Concentration [g/L] | Literature |
|---|---|---|---|---|---|
| Laboratory | Leuconostoc citreum BD1707 | 172.0 | 112 h, 26 °C, 200 rpm, pH 6.12 | 34.86 | [40] |
| Tanticharoenia sakaeratensis | 200.0 | 35 h, 37 °C, 250 rpm | 24.70 | [41] | |
| B. subtilis AF17 | 162.5 | 20 h, 30 °C, 150 rpm, pH 7.0 | 7.90 | [42] | |
| B.subtilis MT453867 | 80.0 | 54 h, 37 °C, 150 rpm, pH 5.0 | 33.00 | [43] | |
| Brenneria goodwinii | 500.0 | 12 h, 35 °C, 8000 rpm, pH 6.0 | 185.00 | [25] | |
| Clostridium acetobutylicum | 28.00 | 72 h, 20 °C, - *, pH 6.0 | - * | [26] | |
| Leuconostoc mesenteroides | 90.0 | 24 h, 30 °C, - *, pH 5.0 | - * | [27] | |
| Sphingobium chungbukense | 10.00 | 24 h, 37 °C, - *, pH 5–10 | - * | [29] | |
| Gluconobacter japonicus LMG 1417 | 720.00 | 24 h, 28 °C, 180 rpm, pH 6.8 | 157.90 | [30] | |
| Peanibacillus polymyxa | 124.00 | 96 h, 30 °C, 250 rpm, pH 7.2 | 68.00 | [31] | |
| Azotobacter chroococcum DSM 2286 + Gluconobacter japonicus LMG 1417 |
809.6 | 48 h, 45 °C, - *, pH 6.2 | 387.40 | [32] | |
| Saccharomyces cereviasie |
50.00 | 48 h, 30 °C, pH 6.5 | 15.00 | [33] | |
| Bioreactor | B. subtilis M | 100.0 | 150 L bioreactor, 24 h, 30 ℃, 200 rpm, pH 7.0, 0.5 vvm |
47.00 | [44] |
| B. subtlis B58 | 130.0 | 16 L bioreactor, 16 h, 37 ℃, 400 rpm, pH 7.0, 1 vvm |
26.65 | [45] | |
| Z. mobilis B-14023 | 299.1 | 42.3 h, 28 ℃, pH 6.0 | 40.20 | [34] | |
| E. herbicola | 50.0 | 5 L bioreactor, 48 h, 25 ℃, 200 rpm, 0.2 vvm |
4.50 | [35] | |
| B. subtilis AF17 | 162.5 | 30 °C, 72 h, 150 rpm, pH 7.0 | 7.90 | [42] | |
| Halomonas smyrnensis AAD6T | 20.00 | 10 L bioreactor 37 °C, 200 rpm, 0.1 vvm, |
18.06 | [45] | |
| Bacillus amyloliquefaciens |
20.00 | 5 L bioreactor, 37 °C, 48 h, 300 rpm, pH 6.0, 6 vvm |
102.00 | [46] | |
| Pichia pastoris | 160.00 | 1,5 L bioreactor, 59 h, 28 °C, 200–1200 rpm |
72.90 | [47] | |
| Saccharomyces cerevisiae |
191.00 | 50 L bioreactor, 48 h, 30 °C, 300 rpm, pH 5.5, 0.5 vvm |
76.00 | [48] |
* not indicated
One of the primary by-products commonly utilized in levan research is molasses, which is derived from sugar production. Gojgic-Cvijovic et al. used diluted molasses in a medium and obtained 53.2 g/L levan during 48 h of fermentation of B. licheniformis NS032 [49]. In recent years, other studies have explored alternative uses of molasses as a sucrose source for levan production, employing strains such as B. lentus V8 [50] or Brachybacterium phenoliresistens, where molasses and date syrup have been used [51]. Another solution to the challenge of searching for substitutes for culture medium substrates is the process described by Erkorkmaz et al., in which the growth and yielding of levan by Halomonas smyrnensis was compared in a medium containing industrial sucrose from sugar beet syrup, salts of various origins as well as three different industrial boron compounds [52]. The most optimal culture medium, consisting of industrial food sucrose from sugar beets, sea salt from Çamaltı, industrial boric acid and borax pentahydrate, yielded 18.60 g/L levan, which is the highest yield recorded for H. smyrnensis to date [52].
Table 2. Examples of the use of various by-products in obtaining levan.
| Substrate/By-Product used | Microorganism | Sucrose Concentration [%] | Levan Concentration [g/L] | Literature |
|---|---|---|---|---|
| Molasses | Halomonas sp. AAD6 | 48–51 | 12.40 | [36] |
| B. licheniformis NS032 | 49.40 | 53.20 | [53] | |
| B. lentus V8 | 54 | 45.34 | [50] | |
| Sugar beet juice and syrup | Z. mobilis 113 | 65 | 9.60 | [54] |
| The pulp after squeezing orange juice | B. atrophaeus | 12–27 | 24.20 | [38] |
| Distillers’ rye stock | Z. mobilis | * | 25.17 | [55] |
| Date syrup | Microbacterium laevaniformans | * | 10.48 | [56] |
| B. phenoliresistens | * | 8.12 | [51] | |
| Sugar cane syrup | Z. mobilis | * | 15.46 | [57] |
| Baklava syrup | Z. mobilis | * | 8.90 | [58] |
| Buckwheat sourdough bread | G. albidus TMW 2.1191; Kozakia baliensis NBRC 16680 | 8.86 ± 21.71; 50.26 ± 25.21 | 13.91; 12.65 | [39] |
* not indicated.
Levan is a multifunctional polymer of great industrial importance; hence, microorganisms characterized by its increased synthesis are still being sought. However, finding the right manufacturer is just the beginning of developing the process of receiving. Hence, it becomes important to improve strains using genetic engineering and optimization of the production process, to search for cheaper substitutes for expensive commercial microbiological substrates or to appropriate technology for obtaining the desired product for specific applications.
3. Properties and Application of Levan
Levan is a biodegradable, non-toxic and highly biocompatible compound, which makes it a very interesting polymer for the development of materials of natural origin. Levan exhibits a wide range of solubility, viscosity and stability depending on its source and production conditions. The properties depend on factors such as molecular weight concentration or branching and can range, for example, from highly soluble to partially soluble or insoluble.
Thanks to its properties, it is used in many fields, such as food production, modern cosmetology, medicine and pharmacy. The diversity in the structure of the levan molecule makes it a polymer that is still not fully understood.
In the food industry, levan is used as an emulsifying, stabilizing and thickening agent [21]. In addition, it can act as a sweetener and be a source of fructooligosaccharides with its prior hydrolysis [59][60]. Some levan fructooligosaccharides are more resistant to degradation in the gastrointestinal tract than their short-chain counterparts and have lower cholesterol levels [61][62]. Levan has prebiotic properties [63], promoting the growth of beneficial intestinal bacteria (Figure 4) [64]. It exhibits high thermal resistance, which makes it possible to process at high temperatures [65]. Recent research indicates the prebiotic properties of levan obtained via enzymatic synthesis. Xu et al. showed that levan increased the number of bacteria of the genus Firmicutes; mainly Megasphaera and Megamonas bacteria, and the proliferation of harmful genera was inhibited (Cedecea and Klebsiella) [66]. Levan also significantly increased total short-chain fatty acid levels, especially for propionic acid, butyric acid and valeric acid, which ensure the proper functioning of the intestines and accelerate the process of healing and regeneration of the intestinal epithelium. They also protect against pathogens (Salmonella spp., Escherichia coli, Campylobacter spp.) by lowering the pH of the environment in which they occur, inhibiting the multiplication of pathogenic microflora [67]. In their previous study, Xu et al. studied levan as a potential stabilizer of yoghurt. They proved that levan can increase the water-holding capacity (WHC) of fermented yogurt (over 77%) compared to fructoligosaccharides (FOS). In addition, levan showed better system stability (Zeta potential and WHC) than FOS. The addition of 0.2–0.5% obtained levan could increase the growth and sustainability of L. bulgaricus and S. thermophilus in fermented yoghurt [66].
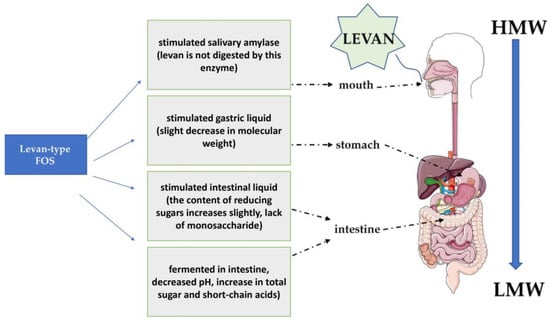
Figure 4. L-FOS and its specific prebiotic properties during digestion in humans based on [44].
Another interesting potential application of levan is the development of food packaging materials. Gan et al. tested its use in edible films with pullulan, chitosan and ε-polylysine that they coated strawberries with [68]. During their storage, they found that the weight loss of strawberries was lower and they retained their firmness and demonstrated antibacterial activity of films against Escherichia coli and Staphylococcus aureus, which are typical foodborne pathogens [68]. Koşarsoy-Ağçeli promotes an innovative approach to nanocomposite polymer-carbohydrate films using chia seed and levan mucus. In this case, for the first time, different degrees of mucus were obtained from chia seeds and the film-forming behavior of the levan biopolymer with these mucilages was studied. Films formed as nanocomposites of mucus, levan seeds and chia obtained at different temperatures exhibited different structural and mechanical properties. It was observed that films obtained with chia mucus and levan retained their antibacterial properties but lost their antifungal properties. These nanocomposites derived from chia seed mucilage and levan hold significant potential for applications in industrial and medical fields. Moreover, the environmentally friendly nature of these films plays a crucial role in promoting environmental protection [69].
Modern cosmetology continues to seek natural compounds that possess desirable properties in the beauty sector. In line with the current natural cosmetic trends, levan emerges as a suitable candidate due to its beneficial properties. Levan has been proven to be safe for human fibroblast cells and erythrocytes, making it a favorable ingredient in cosmetics [4][70][71]. Moreover, levan stimulates the proliferation of human keratinocytes and fibroblasts. It is non-irritating and can alleviate skin irritations caused by substances like sodium lauryl sulfate [72]. Levan, by inhibiting tyrosinase activity, and thus lowering melanin production, may act as a product regulating skin discoloration [73]. In vitro experiments by Kirtel et al. with microbiologically produced levan from Halomonas (HL) not only demonstrated increased proliferation of keratinocytes and fibroblasts, improved skin barrier function and increased type I collagen but also increased ability to rapidly heal wounds within 24 h on 2D wound models. This clearly shows that HL and its derivatives have a high potential for use as natural active ingredients in cosmeceutical and skin-regenerating preparations [28].
In the pharmaceutical industry, levan offers potential contributions to the stabilization and control of drug release in topical emulsion systems. Pantelić et al. have determined that levan is a compatible co-stabilizer with emulsifiers, cetearyl alcohol and sodium cetearyl sulfate, indicating its usefulness in preparations stabilized with anionic or non-ionic excipients [74]. Due to its ability to self-organize in water, it is an interesting component for the encapsulation of active compounds in the form of nanoparticles [75][76][77][78]. Furthermore, levan is also investigated for other “nano” forms, such as polymer micelles [79], nanocomposites [80] or nanoemulsions [78]. It could also be used to obtain hydrogels, e.g., for local skin antifungal therapy with amphotericin B [81]. Studies by Hamad et al. demonstrate the significant role of levan in the healing of burn wounds, accelerating them compared to base cream and saline [82]. Research by Osman et al. highlights the ease of dilution, washing, and loss of adhesion of levan in a humid environment. Researchers demonstrated a strategy for producing a levan-based self-adhesive hydrogel for hemostatic applications and wound healing by coupling catechol with levan. The prepared hydrogels showed significantly enhanced solubility in water and improved adhesion power to the hydrated skin of pigs. Hydrogels also promoted rapid blood clotting and much faster healing of rat skin incisions. Additionally, levan–catechol showed an immune response similar to a negative control response, attributed to significantly lower endotoxin levels compared to native levan [83].
It is worth mentioning that many studies indicate that the properties and subsequent use of levan are related to molecular weight and branching in the molecule. There are still few studies on the utilization of specific levan fractions [84][85] and they are often inconclusive. Previous studies have demonstrated that HMW levan exhibits anticancer activity [86][87] and lowers cholesterol [8], while the fraction of LMW levan has shown a better ability to encapsulate due to the possibility of slow and constant release of active substances resulting from its structure [77][88][89]. A more recent report shows that HMW levan can also be used to encapsulate xenobiotics [90]. Domżał-Kędzia et al. showed the cosmetic use of the LMW fraction of the biopolymer, indicating higher antioxidant properties than in the case of the HMW commercial levan from E. herbicola [4]. Charoenwongpaiboon et al. used ultrasound to reduce molecular weight and increase the bioavailability of levan. The bioactivity of both forms of levan (HMW and LMW) was tested using human osteosarcoma cells (Saos-2). The result clearly showed that the sonicated levan (LMW) had higher anti-proliferative activity in Saos-2 cells than the original levan (HMW) [91]. Despite studies investigating the properties of various levan fractions, the mechanisms of action specific to each fraction and the relationship between structure and properties have not been fully elucidated. Further research in this area is warranted.
References
- Greig-Smith, R.; Steel, T. Levan: A New Bacterial Gum from Sugar. J. Soc. Chem. Ind. Scottish Sect. 1902, 21, 1381.
- Gupta, S.K.; Das, P.; Singh, S.K.; Akhtar, M.S.; Meena, D.K.; Mandal, S.C. Microbial Levan, an Ideal Prebiotic and Immunonutrient in Aquaculture. World Aquac. 1990, 42, 61–64.
- Combie, J. Properties of Levan and Potential Medical Uses. Polysaccharides Drug Deliv. Pharm. Appl. 2006, 934, 13–263.
- Domżał-Kędzia, M.; Lewińska, A.; Jaromin, A.; Weselski, M.; Pluskota, R.; Łukaszewicz, M. Fermentation Parameters and Conditions Affecting Levan Production and Its Potential Applications in Cosmetics. Bioorg. Chem. 2019, 93, 102787.
- Goncalves, B.C.M.; Baldo, C.; Celligoi, M.A.P.C. Levan and Levansucrase—A Mini Review. Int. J. Sci. Technol. Res. 2015, 4, 100–104.
- Mardo, K.; Visnapuu, T.; Gromkova, M.; Aasamets, A.; Viigand, K.; Vija, H.; Alamäe, T. High-Throughput Assay of Levansucrase Variants in Search of Feasible Catalysts for the Synthesis of Fructooligosaccharides and Levan. Molecules 2014, 19, 8434–8455.
- González-Garcinuño, Á.; Tabernero, A.; Domínguez, Á.; Galán, M.A.; Martin del Valle, E.M. Levan and Levansucrases: Polymer, Enzyme, Micro-Organisms and Biomedical Applications. Biocatal. Biotransformation 2017, 36, 233–244.
- Freitas, F.; Reis, M.A.M.; De Lisboa, U.N.; Lisboa, U. De Polysaccharides; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–24.
- Leloup, L.; Driessen, A.J.M.; Freudl, R.; Chambert, R.; Petit-Glatron, M.F. Differential Dependence of Levansucrase and α-Amylase Secretion on SecA (Div) during the Exponential Phase of Growth of Bacillus Subtilis. J. Bacteriol. 1999, 181, 1820–1826.
- Daguer, J.P.; Geissmann, T.; Petit-Glatron, M.F.; Chambert, R. Autogenous Modulation of the Bacillus Subtilis SacB-LevB-YveA Levansucrase Operon by the LevB Transcript. Microbiology 2004, 150, 3669–3679.
- Rey, M.W.; Ramaiya, P.; Nelson, B.A.; Brody-Karpin, S.D.; Zaretsky, E.J.; Tang, M.; Lopez de Leon, A.; Xiang, H.; Gusti, V.; Clausen, I.G.; et al. Complete Genome Sequence of the Industrial Bacterium Bacillus Licheniformis and Comparisons with Closely Related Bacillus Species. Genome Biol. 2004, 5, r77.
- Pereira, Y.; Petit-Glatron, M.F.; Chambert, R. YveB, Encoding Endolevanase LevB, Is Part of the SacB-YveB-YveA Levansucrase Tricistronic Operon in Bacillus Subtilis. Microbiology 2001, 147, 3413–3419.
- Jensen, S.L.; Diemer, M.B.; Lundmark, M.; Larsen, F.H.; Blennow, A.; Mogensen, H.K.; Nielsen, T.H. Levanase from Bacillus Subtilis Hydrolyses β-2,6 Fructosyl Bonds in Bacterial Levans and in Grass Fructans. Int. J. Biol. Macromol. 2016, 85, 514–521.
- Méndez-Lorenzo, L.; Porras-Domínguez, J.R.; Raga-Carbajal, E.; Olvera, C.; Rodríguez-Alegría, M.E.; Carrillo-Nava, E.; Costas, M.; Munguía, A.L. Intrinsic Levanase Activity of Bacillus Subtilis 168 Levansucrase (SacB). PLoS ONE 2015, 10, e0143394.
- Shih, I.L.; Chen, L.D.; Wang, T.C.; Wu, J.Y.; Liaw, K.S. Tandem Production of Levan and Ethanol by Microbial Fermentation. Green Chem. 2010, 12, 1242–1247.
- Marvasi, M.; Visscher, P.T.; Casillas Martinez, L. Exopolymeric Substances (EPS) from Bacillus Subtilis: Polymers and Genes Encoding Their Synthesis. FEMS Microbiol. Lett. 2010, 313, 1–9.
- Martin, I.; Debarbouille, M.; Klier, A.; Rapoport, G. Induction and Metabolite Regulation of Levanase Synthesis in Bacillus Subtilis. J. Bacteriol. 1989, 171, 1885–1892.
- Martin, I.; Débarbouillé, M.; Ferrari, E.; Klier, A.; Rapoport, G. Characterization of the Levanase Gene of Bacillus Subtilis Which Shows Homology to Yeast Invertase. MGG Mol. Gen. Genet. 1987, 208, 177–184.
- Martin-Verstraete, I.; Débarbouillé, M.; Klier, A.; Rapoport, G. Levanase Operon of Bacillus Subtilis Includes a Fructose-Specific Phosphotransferase System Regulating the Expression of the Operon. J. Mol. Biol. 1990, 214, 657–671.
- Débarbouillé, M.; Martin-Verstraete, I.; Arnaud, M.; Klier, A.; Rapoport, G. Positive and Negative Regulation Controlling Expression of the Sac Genes in Bacillus Subtilis. Res. Microbiol. 1991, 142, 757–764.
- Wanker, E.; Huber, A.; Schwab, H. Purification and Characterization of the Bacillus Subtilis Levanase Produced in Escherichia coli. Appl. Environ. Microbiol. 1995, 61, 1953–1958.
- Raga-Carbajal, E.; Carrillo-Nava, E.; Costas, M.; Porras-Dominguez, J.; López-Munguía, A.; Olvera, C. Size Product Modulation by Enzyme Concentration Reveals Two Distinct Levan Elongation Mechanisms in Bacillus Subtilis Levansucrase. Glycobiology 2016, 26, 377–385.
- Shih, I.-L.; Yu, Y.-T.; Shieh, C.-J.; Hsieh, C.-Y. Selective Production and Characterization of Levan by Bacillus Subtilis (Natto) Takahashi. J. Agric. Food Chem. 2005, 53, 82118215.
- Euzenat, O.; Guibert, A.; Combes, D. Production of Fructo-Oligosaccharides by Levansucrase from Bacillus Subtilis C4. Process Biochem. 1997, 32, 237–243.
- Liu, Q.; Yu, S.; Zhang, T.; Jiang, B.; Mu, W. Efficient Biosynthesis of Levan from Sucrose by a Novel Levansucrase from Brenneria Goodwinii. Carbohydr. Polym. 2017, 157, 1732–1740.
- Gao, S.; Qi, X.; Hart, D.J.; Gao, H.; An, Y. Expression and Characterization of Levansucrase from Clostridium Acetobutylicum. J. Agric. Food Chem. 2017, 65, 867–871.
- Ishida, R.; Sakaguchi, K.; Matsuzaki, C.; Katoh, T.; Ishida, N.; Yamamoto, K.; Hisa, K. Levansucrase from Leuconostoc Mesenteroides NTM048 Produces a Levan Exopolysaccharide with Immunomodulating Activity. Biotechnol. Lett. 2016, 38, 681–687.
- Kirtel, O.; Menéndez, C.; Versluys, M.; Van den Ende, W.; Hernández, L.; Toksoy Öner, E. Levansucrase from Halomonas Smyrnensis AAD6T: First Halophilic GH-J Clan Enzyme Recombinantly Expressed, Purified, and Characterized. Appl. Microbiol. Biotechnol. 2018, 102, 9207–9220.
- Lee, S.Y.; Shin, W.-R.; Sekhon, S.S.; Lee, J.-P.; Kim, Y.-C.; Ahn, J.-Y.; Kim, Y.-H. Molecular Docking Analysis and Biochemical Evaluation of Levansucrase from Sphingobium Chungbukense DJ77. ACS Comb. Sci. 2018, 20, 414–422.
- Hövels, M.; Kosciow, K.; Kniewel, J.; Jakob, F.; Deppenmeier, U. High Yield Production of Levan-Type Fructans by Gluconobacter Japonicus LMG 1417. Int. J. Biol. Macromol. 2020, 164, 295–303.
- Liyaskina, E.V.; Rakova, N.A.; Kitykina, A.A.; Rusyaeva, V.V.; Toukach, P.V.; Fomenkov, A.; Vainauskas, S.; Roberts, R.J.; Revin, V.V. Production and Characterization of the Exopolysaccharide from Strain Paenibacillus Polymyxa 2020. PLoS ONE 2021, 16, e0253482.
- Hövels, M.; Kosciow, K.; Deppenmeier, U. Characterization of a Novel Endo-Levanase from Azotobacter Chroococcum DSM 2286 and Its Application for the Production of Prebiotic Fructooligosaccharides. Carbohydr. Polym. 2021, 255, 117384.
- Franken, J.; Brandt, B.A.; Tai, S.L.; Bauer, F.F. Biosynthesis of Levan, a Bacterial Extracellular Polysaccharide, in the Yeast Saccharomyces Cerevisiae. PLoS ONE 2013, 8, e77499.
- Silbir, S.; Dagbagli, S.; Yegin, S.; Baysal, T.; Goksungur, Y. Levan Production by Zymomonas Mobilis in Batch and Continuous Fermentation Systems. Carbohydr. Polym. 2014, 99, 454–461.
- Keith, J.; Wiley, B.; Ball, D.; Arcidiacono, S.; Zorfass, D.; Mayer, J.; Kaplan, D. Continuous Culture System for Production of Biopolymer Levan Using Erwinia Herbicola. Biotechnol. Bioeng. 1991, 38, 557–560.
- Küçükaşik, F.; Kazak, H.; Güney, D.; Finore, I.; Poli, A.; Yenigün, O.; Nicolaus, B.; Öner, E.T. Molasses as Fermentation Substrate for Levan Production by Halomonas sp. Appl. Microbiol. Biotechnol. 2011, 89, 1729–1740.
- Seibel, J.; Moraru, R.; Götze, S.; Buchholz, K.; Na’amnieh, S.; Pawlowski, A.; Hecht, H.J.; Secchi, M.; Castellani, V.; Collina, E.; et al. Selenium Enriched Green Tea Increase Stability of Lactobacillus Casei and Lactobacillus Plantarum in Chitosan Coated Alginate Microcapsules during Exposure to Simulated Gastrointestinal and Refrigerated Conditions. Int. J. Biol. Macromol. 2020, 21, 406–411.
- González-Garcinuño, Á.; Tabernero, A.; Sánchez-Álvarez, J.M.; Galán, M.A.; Martin del Valle, E.M. Effect of Bacteria Type and Sucrose Concentration on Levan Yield and Its Molecular Weight. Microb. Cell Fact. 2017, 16, 91.
- Ua-Arak, T.; Jakob, F.; Vogel, R.F. Influence of Levan-Producing Acetic Acid Bacteria on Buckwheat-Sourdough Breads. Food Microbiol. 2017, 65, 95–104.
- Han, J.; Feng, H.; Wang, X.; Liu, Z.; Wu, Z. Levan from Leuconostoc Citreum BD1707: Production Optimization and Changes in Molecular Weight Distribution during Cultivation. BMC Biotechnol. 2021, 21, 14.
- Aramsangtienchai, P.; Kongmon, T.; Pechroj, S.; Srisook, K. Enhanced Production and Immunomodulatory Activity of Levan from the Acetic Acid Bacterium, Tanticharoenia Sakaeratensis. Int. J. Biol. Macromol. 2020, 163, 574–581.
- Bouallegue, A.; Casillo, A.; Chaari, F.; Cimini, D.; Corsaro, M.M.; Bachoual, R.; Ellouz-Chaabouni, S. Statistical Optimization of Levan: Influence of the Parameter on Levan Structure and Angiotensin I-Converting Enzyme Inhibitory. Int. J. Biol. Macromol. 2020, 158, 945–952.
- Gamal, A.A.; Abbas, H.Y.; Abdelwahed, N.A.M.; Kashef, M.T.; Mahmoud, K.; Esawy, M.A.; Ramadan, M.A. Optimization Strategy of Bacillus Subtilis MT453867 Levansucrase and Evaluation of Levan Role in Pancreatic Cancer Treatment. Int. J. Biol. Macromol. 2021, 182, 1590–1601.
- Ragab, T.I.M.; Malek, R.A.; Elsehemy, I.A.; Farag, M.M.S.; Salama, B.M.; Abd EL-Baseer, M.A.; Gamal-Eldeen, A.M.; El Enshasy, H.A.; Esawy, M.A. Scaling up of Levan Yield in Bacillus Subtilis M and Cytotoxicity Study on Levan and Its Derivatives. J. Biosci. Bioeng. 2019, 127, 655–662.
- Hamid, K.R.A.; Elsayed, E.A.; Enshasy, H.A.E.; Esawy, M.; Malek, R.A. Bioprocess Optimization for Levan Production by Bacillus Subtilis B58. J. Sci. Ind. Res. (India) 2018, 77, 386–393.
- Gu, Y.; Zheng, J.; Feng, J.; Cao, M.; Gao, W.; Quan, Y.; Dang, Y.; Wang, Y.; Wang, S.; Song, C. Improvement of Levan Production in Bacillus Amyloliquefaciens through Metabolic Optimization of Regulatory Elements. Appl. Microbiol. Biotechnol. 2017, 101, 4163–4174.
- Ávila-Fernández, Á.; Montiel, S.; Rodríguez-Alegría, M.E.; Caspeta, L.; López Munguía, A. Simultaneous Enzyme Production, Levan-Type FOS Synthesis and Sugar by-Products Elimination Using a Recombinant Pichia Pastoris Strain Expressing a Levansucrase-Endolevanase Fusion Enzyme. Microb. Cell Fact. 2023, 22, 18.
- Ko, H.; Bae, J.H.; Sung, B.H.; Kim, M.J.; Kim, C.H.; Oh, B.R.; Sohn, J.H. Efficient Production of Levan Using a Recombinant Yeast Saccharomyces Cerevisiae Hypersecreting a Bacterial Levansucrase. J. Ind. Microbiol. Biotechnol. 2019, 46, 1611–1620.
- Sarilmiser, H.K.; Ates, O.; Ozdemir, G.; Arga, K.Y.; Oner, E.T. Effective Stimulating Factors for Microbial Levan Production by Halomonas Smyrnensis AAD6T. J. Biosci. Bioeng. 2015, 119, 455–463.
- Abou-taleb, K.A.; Abdel-Monem, M.O.; Yassin, M.H.; Draz, A.A.; Blaiotta, G. Production, Purification and Characterization of Levan Polymer from Bacillus Lentus V8 Strain. Br. Microbiol. Res. J. 2015, 5, 22–32.
- Moussa, T.A.A.; Al-Qaysi, S.A.S.; Thabit, Z.A.; Kadhem, S.B. Microbial Levan from Brachybacterium Phenoliresistens: Characterization and Enhancement of Production. Process Biochem. 2017, 57, 9–15.
- Erkorkmaz, B.A.; Kırtel, O.; Ateş Duru, Ö.; Toksoy Öner, E. Development of a Cost-Effective Production Process for Halomonas Levan. Bioprocess Biosyst. Eng. 2018, 41, 1247–1259.
- Gojgic-Cvijovic, G.D.; Jakovljevic, D.M.; Loncarevic, B.D.; Todorovic, N.M.; Pergal, M.V.; Ciric, J.; Loos, K.; Beskoski, V.P.; Vrvic, M.M. Production of Levan by Bacillus Licheniformis NS032 in Sugar Beet Molasses-Based Medium. Int. J. Biol. Macromol. 2019, 121, 142–151.
- Bekers, M.; Linde, R.; Danilevich, A.; Kaminska, E.; Upite, D.; Vigants, A.; Scherbaka, R. Sugar Beet Diffusion Juice and Syrup as Media for Ethanol and Levan Production by Zymomonas Mobilis. Food Biotechnol. 1999, 13, 107–119.
- Bekers, M.; Grube, M.; Vulfa, L.; Upite, D.; Kaminska, E.; Scherbaka, R.; Vigants, A.; Danilevich, A. Stillage as a Source of Growth Promoting Biofactors and a Stimulator of Levan and Extracellular Levansucrase Synthesis for Zymomonas Mobilis. Food Technol. Biotechnol. 2002, 40, 305–310.
- Bekers, M.; Grube, M.; Vulfa, L.; Upite, D.; Kaminska, E.E.; Scherbaka, R.; Vigants, A.; Danilevich, A.; de Oliveira, M.R.; da Silva, R.S.S.F.; et al. Microbial Production of Levan Using Date Syrup and Investigation of Its Properties. Int. J. Nutr. Food Eng. World Acad. Sci. Eng. Technol. 2007, 13, 107–119.
- de Oliveira, M.R.; da Silva, R.S.S.F.; Buzato, J.B.; Celligoi, M.A.P.C. Study of Levan Production by Zymomonas Mobilis Using Regional Low-Cost Carbohydrate Sources. Biochem. Eng. J. 2007, 37, 177–183.
- Omeroglu, M.A.; Gonul-Baltaci, N.; Arslan, N.P.; Adiguzel, A.; Taskin, M. Microbial Conversion of Waste Baklava Syrup to Biofuels and Bioproducts. Biocatal. Agric. Biotechnol. 2022, 42, 102364.
- Mummaleti, G.; Sarma, C.; Kalakandan, S.K.; Gazula, H.; Sivanandham, V.; Anandharaj, A. Characterization of Levan Produced from Coconut Inflorescence Sap Using Bacillus Subtilis and Its Application as a Sweetener. LWT 2022, 154, 112697.
- Bersaneti, G.T.; Pan, N.C.; Baldo, C.; Celligoi, M.A.P.C. Co-Production of Fructooligosaccharides and Levan by Levansucrase from Bacillus Subtilis Natto with Potential Application in the Food Industry. Appl. Biochem. Biotechnol. 2018, 184, 838–851.
- de Paula, V.C.; Pinheiro, I.O.; Lopes, C.E.; Calazans, G.M.T. Microwave-Assisted Hydrolysis of Zymomonas Mobilis Levan Envisaging Oligofructan Production. Bioresour. Technol. 2008, 99, 2466–2470.
- Zhang, W.; Xu, W.; Ni, D.; Dai, Q.; Guang, C.; Zhang, T.; Mu, W. An Overview of Levan-Degrading Enzyme from Microbes. Appl. Microbiol. Biotechnol. 2019, 103, 7891–7902.
- Cai, G.; Wu, D.; Li, X.; Lu, J. Levan from Bacillus Amyloliquefaciens JN4 Acts as a Prebiotic for Enhancing the Intestinal Adhesion Capacity of Lactobacillus Reuteri JN101. Int. J. Biol. Macromol. 2020, 146, 482–487.
- Cheng, R.; Cheng, L.; Zhao, Y.; Wang, L.; Wang, S.; Zhang, J. Biosynthesis and Prebiotic Activity of a Linear Levan from a New Paenibacillus Isolate. Appl. Microbiol. Biotechnol. 2021, 105, 769–787.
- Shimizu, N.; Abea, A.; Ushiyama, T.; Toksoy Öner, E. Effect of Temperature on the Hydrolysis of Levan Treated with Compressed Hot Water Fluids. Food Sci. Nutr. 2020, 8, 2004–2014.
- Xu, M.; Pan, L.; Zhou, Z.; Han, Y. Structural Characterization of Levan Synthesized by a Recombinant Levansucrase and Its Application as Yogurt Stabilizers. Carbohydr. Polym. 2022, 291, 119519.
- Kovanda, L.; Zhang, W.; Wei, X.; Luo, J.; Wu, X.; Atwill, E.R.; Vaessen, S.; Li, X.; Liu, Y. In Vitro Antimicrobial Activities of Organic Acids and Their Derivatives on Several Species of Gram-Negative and Gram-Positive Bacteria. Molecules 2019, 24, 3770.
- Gan, L.; Jiang, G.; Yang, Y.; Zheng, B.; Zhang, S.; Li, X.; Tian, Y.; Peng, B. Development and Characterization of Levan/Pullulan/Chitosan Edible Films Enriched with ε-Polylysine for Active Food Packaging. Food Chem. 2022, 388, 132989.
- Koşarsoy Ağçeli, G. A New Approach to Nanocomposite Carbohydrate Polymer Films: Levan and Chia Seed Mucilage. Int. J. Biol. Macromol. 2022, 218, 751–759.
- Kim, K.S.H.; Chung, C.B.; Kim, Y.H.; Kim, K.S.H.; Han, C.S.; Kim, C.H. Cosmeceutical Properties of Levan Produced by Zymomonas Mobilis. Int. J. Cosmet. Sci. 2006, 28, 231.
- Lewińska, A.; Domżał-Kędzia, M.; Kierul, K.; Bochynek, M.; Pannert, D.; Nowaczyk, P.; Łukaszewicz, M. Targeted Hybrid Nanocarriers as a System Enhancing the Skin Structure. Molecules 2021, 26, 1063.
- Kim, T.; Kim, K.; Ryo, K.; Lee, O.; Kim, T. Cosmetic Composition Containing Levan Having Cell Proliferation, Skin-Moisturizing and Irritation-Alleviating Effects. Patent JP2003277225A, 2 October 2003.
- Masayo, F.; Takayuki, T. Beautifully Whitening Agent 2006. Patent JP2006052146A, 23 February 2006.
- Pantelić, I.; Lukić, M.; Gojgić-Cvijović, G.; Jakovljević, D.; Nikolić, I.; Lunter, D.J.; Daniels, R.; Savić, S. Bacillus Licheniformis Levan as a Functional Biopolymer in Topical Drug Dosage Forms: From Basic Colloidal Considerations to Actual Pharmaceutical Application. Eur. J. Pharm. Sci. 2020, 142, 105109.
- Sezer, A.D.; Kazak Sarılmışer, H.; Rayaman, E.; Çevikbaş, A.; Öner, E.T.; Akbuğa, J. Development and Characterization of Vancomycin-Loaded Levan-Based Microparticular System for Drug Delivery. Pharm. Dev. Technol. 2017, 22, 627–634.
- Cinan, E.; Cesur, S.; Erginer Haskoylu, M.; Gunduz, O.; Toksoy Oner, E. Resveratrol-Loaded Levan Nanoparticles Produced by Electrohydrodynamic Atomization Technique. Nanomaterials 2021, 11, 2582.
- Kim, S.J.; Bae, P.K.; Chung, B.H. Self-Assembled Levan Nanoparticles for Targeted Breast Cancer Imaging. Chem. Commun. 2015, 51, 107–110.
- Richa, R.; Roy Choudhury, A. Exploration of Polysaccharide Based Nanoemulsions for Stabilization and Entrapment of Curcumin. Int. J. Biol. Macromol. 2020, 156, 1287–1296.
- Patel, P.; Agrawal, Y. Preparation and In-Vitro Evaluation of Levan Micelles: A Polyfructan Based Nano-Carrier for Breast Cancer Targeted Delivery. Drug Deliv. Lett. 2019, 9, 97–107.
- Xu, W.; Peng, J.; Ni, D.; Zhang, W.; Wu, H.; Mu, W. Preparation, Characterization and Application of Levan/Montmorillonite Biocomposite and Levan/BSA Nanoparticle. Carbohydr. Polym. 2020, 234, 115921.
- Demirci, T.; Hasköylü, M.E.; Eroğlu, M.S.; Hemberger, J.; Toksoy Öner, E. Levan-Based Hydrogels for Controlled Release of Amphotericin B for Dermal Local Antifungal Therapy of Candidiasis. Eur. J. Pharm. Sci. 2020, 145, 105255.
- Hamada, M.A.; Hassan, R.A.; Abdou, A.M.; Elsaba, Y.M.; Aloufi, A.S.; Sonbol, H.; Korany, S.M. Bio_Fabricated Levan Polymer from Bacillus Subtilis MZ292983.1 with Antibacterial, Antibiofilm, and Burn Healing Properties. Appl. Sci. 2022, 12, 6413.
- Osman, A.; Lin, E.; Hwang, D.S. A Sticky Carbohydrate Meets a Mussel Adhesive: Catechol-Conjugated Levan for Hemostatic and Wound Healing Applications. Carbohydr. Polym. 2023, 299, 120172.
- Hundschell, C.S.; Jakob, F.; Wagemans, A.M. Molecular Weight Dependent Structure and Polymer Density of the Exopolysaccharide Levan. arXiv 2019, arXiv:1909.07737.
- de Siqueira, E.C.; Rebouças, J.d.S.; Pinheiro, I.O.; Formiga, F.R. Levan-Based Nanostructured Systems: An Overview. Int. J. Pharm. 2020, 580, 119242.
- Makino Tadashi, I.; Masayuki Yamada, K.; Jun-ichi, K. Fast Dissolving Tablet and Production. Patent US005501861A, 5 March 1996.
- Kang, S.A.; Jang, K.-H.; Seo, J.-W.; Kim, K.H.; Kim, Y.H.; Rairakhwada, D.; Seo, M.Y.; Lee, J.O.; Do Ha, S.; Kim, C.-H.; et al. Levan: Applications and Perspectives. In Microbial Production of Biopolymers and Polymer Precursors; Caister Academic Press: Norwich, UK, 2009; pp. 145–161.
- Elvassore, N.; Bertucco, A.; Caliceti, P. Production of Insulin-Loaded Poly(Ethylene Glycol)/Poly(L-Lactide) (PEG/PLA) Nanoparticles by Gas Antisolvent Techniques. J. Pharm. Sci. 2001, 90, 1628–1636.
- Bahadori, F.; Eskandari, Z.; Ebrahimi, N.; Bostan, M.S.; Eroğlu, M.S.; Oner, E.T. Development and Optimization of a Novel PLGA-Levan Based Drug Delivery System for Curcumin, Using a Quality-by-Design Approach. Eur. J. Pharm. Sci. 2019, 138, 105037.
- González-Garcinuño, Á.; Tabernero, A.; Marcelo, G.; Sebastián, V.; Arruebo, M.; Santamaría, J.; Martín del Valle, E. Differences in Levan Nanoparticles Depending on Their Synthesis Route: Microbial vs Cell-Free Systems. Int. J. Biol. Macromol. 2019, 137, 62–68.
- Charoenwongpaiboon, T.; Wangpaiboon, K.; Septham, P.; Jiamvoraphong, N.; Issaragrisil, S.; Pichyangkura, R.; Lorthongpanich, C. Production and Bioactivities of Nanoparticulated and Ultrasonic-Degraded Levan Generated by Erwinia Tasmaniensis Levansucrase in Human Osteosarcoma Cells. Int. J. Biol. Macromol. 2022, 221, 1121–1129.
More
Information
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
3.4K
Revisions:
2 times
(View History)
Update Date:
26 Jul 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No