 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Grzegorz Czerwonka | -- | 2433 | 2023-07-06 11:33:34 | | | |
| 2 | Rita Xu | -43 word(s) | 2390 | 2023-07-06 11:41:27 | | |
Video Upload Options
Skin and wound infections are serious medical problems, and the diversity of bacteria makes such infections difficult to treat. Bacteria possess many virulence factors, among which motility plays a key role in skin infections. This feature allows for movement over the skin surface and relocation into the wound. The aim is to review the type of bacterial movement and to indicate the underlying mechanisms than can serve as a target for developing or modifying antibacterial therapies applied in wound infection treatment. Five types of bacterial movement are distinguished: appendage-dependent (swimming, swarming, and twitching) and appendage-independent (gliding and sliding). All of them allow bacteria to relocate and aid bacteria during infection. Swimming motility allows bacteria to spread from ‘persister cells’ in biofilm microcolonies and colonise other tissues. Twitching motility enables bacteria to press through the tissues during infection, whereas sliding motility allows cocci (defined as non-motile) to migrate over surfaces. Bacteria during swarming display greater resistance to antimicrobials. Molecular motors generating the focal adhesion complexes in the bacterial cell leaflet generate a ‘wave’, which pushes bacterial cells lacking appendages, thereby enabling movement.
1. Introduction
2. Types of Bacterial Movement
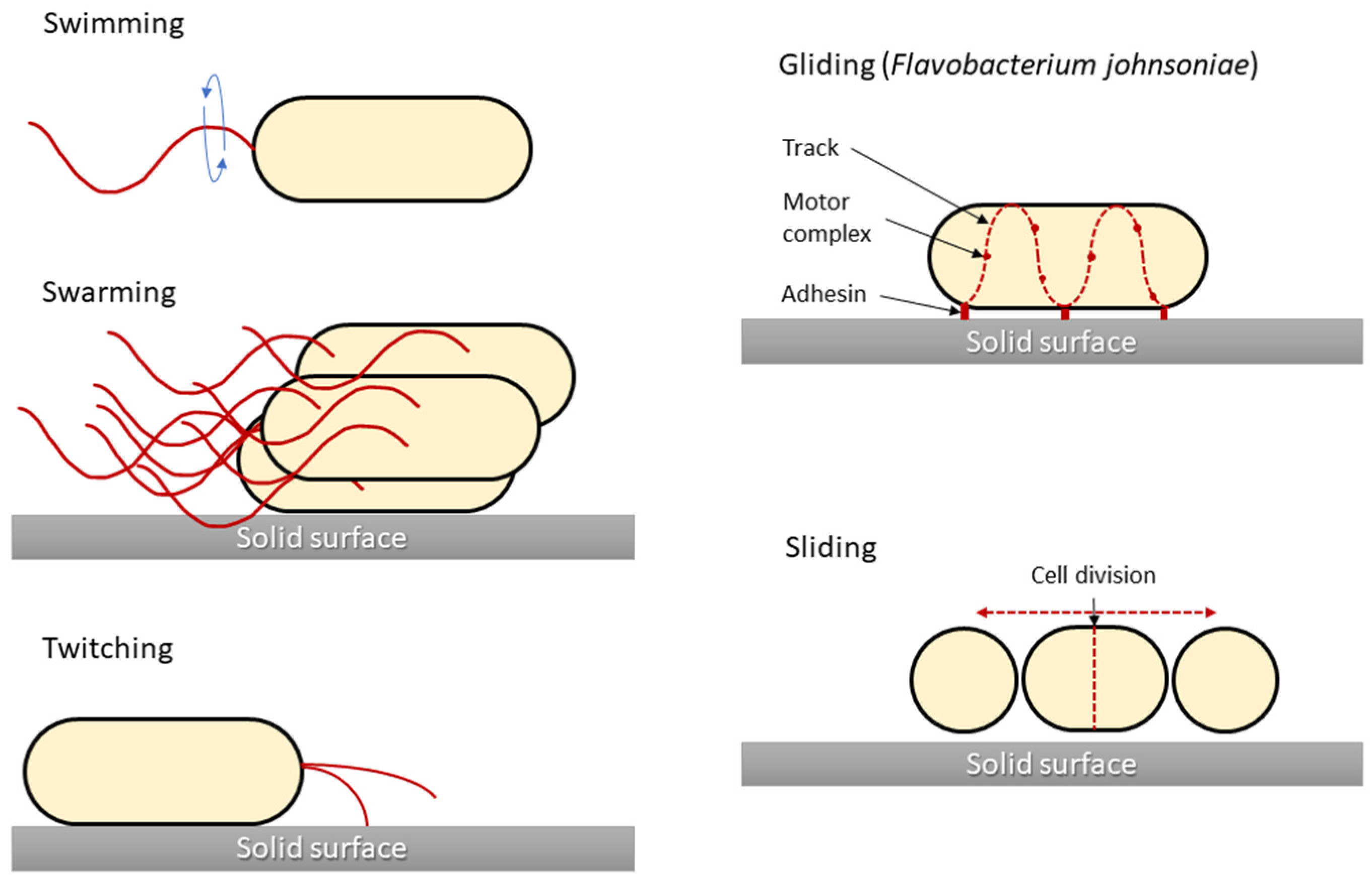
2.1. Swimming Motility
2.2. Swarming Motility
References
- Schommer, N.N.; Gallo, R.L. Structure and Function of the Human Skin Microbiome. Trends Microbiol. 2013, 21, 660–668.
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The Human Skin Microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155.
- DiNubile, M.J.; Lipsky, B.A. Complicated Infections of Skin and Skin Structures: When the Infection Is More than Skip Deep. J. Antimicrob. Chemother. 2004, 53, ii37–ii50.
- Kaye, K.S.; Petty, L.A.; Shorr, A.F.; Zilberberg, M.D. Current Epidemiology, Etiology, and Burden of Acute Skin Infections in the United States. Clin. Infect. Dis. 2019, 68, S193–S199.
- Tognetti, L.; Martinelli, C.; Berti, S.; Hercogova, J.; Lotti, T.; Leoncini, F.; Moretti, S. Bacterial Skin and Soft Tissue Infections: Review of the Epidemiology, Microbiology, Aetiopathogenesis and Treatment: A Collaboration between Dermatologists and Infectivologists. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 931–941.
- Plewig, G.; French, L.; Ruzicka, T.; Kaufmann, R.; Hertl, M. Braun-Falco’s Dermatology, 2020th ed.; Springer: Berlin/Heidelberg, Germany, 2020.
- Cates, J.E.; Mitrani-Gold, F.S.; Li, G.; Mundy, L.M. Systematic Review and Meta-Analysis to Estimate Antibacterial Treatment Effect in Acute Bacterial Skin and Skin Structure Infection. Antimicrob. Agents Chemother. 2015, 59, 4510–4520.
- Olsson, M.; Järbrink, K.; Divakar, U.; Bajpai, R.; Upton, Z.; Schmidtchen, A.; Car, J. The Humanistic and Economic Burden of Chronic Wounds: A Systematic Review: The Humanistic and Economic Burden of Chronic Wounds. Wound Repair Regen. 2019, 27, 114–125.
- Bessa, L.J.; Fazii, P.; di Giulio, M.; Cellini, L. Bacterial Isolates from Infected Wounds and Their Antibiotic Susceptibility Pattern: Some Remarks about Wound Infection. Int. Wound J. 2015, 12, 47–52.
- Puca, V.; Marulli, R.Z.; Grande, R.; Vitale, I.; Niro, A.; Molinaro, G.; Prezioso, S.; Muraro, R.; di Giovanni, P. Microbial Species Isolated from Infected Wounds and Antimicrobial Resistance Analysis: Data Emerging from a Three-Years Retrospective Study. Antibiotics 2021, 10, 1162.
- Wolcott, R.D.; Hanson, J.D.; Rees, E.J.; Koenig, L.D.; Phillips, C.D.; Wolcott, R.A.; Cox, S.B.; White, J.S. Analysis of the Chronic Wound Microbiota of 2,963 Patients by 16S RDNA Pyrosequencing. Wound Repair Regen. 2016, 24, 163–174.
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539.
- Guoqi, W.; Zhirui, L.; Song, W.; Tongtong, L.; Lihai, Z.; Licheng, Z.; Peifu, T. Negative Pressure Wound Therapy Reduces the Motility of Pseudomonas Aeruginosa and Enhances Wound Healing in a Rabbit Ear Biofilm Infection Model. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2018, 111, 1557–1570.
- Mousavi, M.; Behrouz, B.; Irajian, G.; Mahdavi, M.; Korpi, F.; Motamedifar, M. Passive Immunization against Pseudomonas Aeruginosa Recombinant PilA in a Murine Burn Wound Model. Microb. Pathog. 2016, 101, 83–88.
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281.
- Nakagami, G.; Morohoshi, T.; Ikeda, T.; Ohta, Y.; Sagara, H.; Huang, L.; Nagase, T.; Sugama, J.; Sanada, H. Contribution of Quorum Sensing to the Virulence of Pseudomonas Aeruginosa in Pressure Ulcer Infection in Rats. Wound Repair Regen. 2011, 19, 214–222.
- Han, G.; Ceilley, R. Chronic Wound Healing: A Review of Current Management and Treatments. Adv. Ther. 2017, 34, 599–610.
- Huang, X.; Liang, P.; Jiang, B.; Zhang, P.; Yu, W.; Duan, M.; Guo, L.; Cui, X.; Huang, M.; Huang, X. Hyperbaric Oxygen Potentiates Diabetic Wound Healing by Promoting Fibroblast Cell Proliferation and Endothelial Cell Angiogenesis. Life Sci. 2020, 259, 118246.
- Agarwal, P.; Kukrele, R.; Sharma, D. Vacuum Assisted Closure (VAC)/Negative Pressure Wound Therapy (NPWT) for Difficult Wounds: A Review. J. Clin. Orthop. Trauma 2019, 10, 845–848.
- Gupta, S.; Gabriel, A.; Lantis, J.; Téot, L. Clinical Recommendations and Practical Guide for Negative Pressure Wound Therapy with Instillation. Int. Wound J. 2016, 13, 159–174.
- Kloth, L.C. Discussion: Advanced Technologies to Improve Wound Healing: Electrical Stimulation, Vibration Therapy, and Ultrasound-What Is the Evidence? Plast. Reconstr. Surg. 2016, 138, 105–106.
- Balakatounis, K.C.; Angoules, A.G. Low-Intensity Electrical Stimulation in Wound Healing: Review of the Efficacy of Externally Applied Currents Resembling the Current of Injury. Eplasty 2008, 8, e28.
- Ashrafi, M.; Alonso-Rasgado, T.; Baguneid, M.; Bayat, A. The Efficacy of Electrical Stimulation in Lower Extremity Cutaneous Wound Healing: A Systematic Review. Exp. Dermatol. 2017, 26, 171–178.
- Singh, M.R.; Saraf, S.; Vyas, A.; Jain, V.; Singh, D. Innovative Approaches in Wound Healing: Trajectory and Advances. Artif. Cells Nanomed. Biotechnol. 2013, 41, 202–212.
- Bekara, F.; Vitse, J.; Fluieraru, S.; Masson, R.; de Runz, A.; Georgescu, V.; Bressy, G.; Labbé, J.L.; Chaput, B.; Herlin, C. New Techniques for Wound Management: A Systematic Review of Their Role in the Management of Chronic Wounds. Arch. Plast. Surg. 2018, 45, 102–110.
- Henrichsen, J. Bacterial Surface Translocation: A Survey and a Classification. Bacteriol. Rev. 1972, 36, 478–503.
- Shrout, J.D. A Fantastic Voyage for Sliding Bacteria. Trends Microbiol. 2015, 23, 244–246.
- Minnullina, L.; Kostennikova, Z.; Evtugin, V.; Akosah, Y.; Sharipova, M.; Mardanova, A. Diversity in the Swimming Motility and Flagellar Regulon Structure of Uropathogenic Morganella Morganii Strains. Int. Microbiol. 2022, 25, 111–122.
- Corral, J.; Sebastià, P.; Coll, N.S.; Barbé, J.; Aranda, J.; Valls, M. Twitching and Swimming Motility Play a Role in Ralstonia Solanacearum Pathogenicity. Msphere 2020, 5, e00740-19.
- Miyata, M.; Robinson, R.C.; Uyeda, T.Q.P.; Fukumori, Y.; Fukushima, S.I.; Haruta, S.; Homma, M.; Inaba, K.; Ito, M.; Kaito, C.; et al. Tree of Motility—A Proposed History of Motility Systems in the Tree of Life. Genes Cells 2020, 25, 6–21.
- Colin, R.; Ni, B.; Laganenka, L.; Sourjik, V. Multiple Functions of Flagellar Motility and Chemotaxis in Bacterial Physiology. FEMS Microbiol. Rev. 2021, 45, fuab038.
- Li, Y.; Xia, H.; Bai, F.; Song, X.; Zhuang, L.; Xu, H.; Zhang, X.; Qiao, M. PA5001 Gene Involves in Swimming Motility and Biofilm Formation in Pseudomonas Aeruginosa. Microb. Pathog. 2020, 144, 103982.
- Terashima, H.; Kawamoto, A.; Morimoto, Y.V.; Imada, K.; Minamino, T. Structural Differences in the Bacterial Flagellar Motor among Bacterial Species. Biophys. Physicobiol. 2017, 14, 191–198.
- Bardy, S.L.; Ng, S.Y.M.; Jarrell, K.F. Prokaryotic Motility Structures. Microbiology 2003, 149, 295–304.
- Sun, E.; Liu, S.; Hancock, R.E.W. Surfing Motility: A Conserved yet Diverse Adaptation among Motile Bacteria. J. Bacteriol. 2018, 191, 5569–5576.
- Schauer, O.; Mostaghaci, B.; Colin, R.; Hürtgen, D.; Kraus, D.; Sitti, M.; Sourjik, V. Motility and Chemotaxis of Bacteria-Driven Microswimmers Fabricated Using Antigen 43-Mediated Biotin Display. Sci. Rep. 2018, 8, 9801.
- Schwanbeck, J.; Oehmig, I.; Groß, U.; Zautner, A.E.; Bohne, W. Clostridioides Difficile Single Cell Swimming Strategy: A Novel Motility Pattern Regulated by Viscoelastic Properties of the Environment. Front. Microbiol. 2021, 12, 715220.
- Wang, X.; Vu, A.; Lee, K.; Dahlquist, F.W. CheA-Receptor Interaction Sites in Bacterial Chemotaxis. J. Mol. Biol. 2012, 422, 282–290.
- Khong, N.Z.J.; Zeng, Y.; Lai, S.K.; Koh, C.G.; Liang, Z.X.; Chiam, K.H.; Li, H.Y. Dynamic Swimming Pattern of Pseudomonas Aeruginosa near a Vertical Wall during Initial Attachment Stages of Biofilm Formation. Sci. Rep. 2021, 11, 1952.
- Samad, T.; Billings, N.; Birjiniuk, A.; Crouzier, T.; Doyle, P.S.; Ribbeck, K. Swimming Bacteria Promote Dispersal of Non-Motile Staphylococcal Species. ISME J. 2017, 11, 1933–1937.
- Madukoma, C.S.; Liang, P.; Dimkovikj, A.; Chen, J.; Lee, S.W.; Chen, D.Z.; Shrout, J.D. Single Cells Exhibit Differing Behavioral Phases during Early Stages of Pseudomonas Aeruginosa Swarming. J. Bacteriol. 2019, 201, e00184-19.
- Morris, J.D.; Hewitt, J.L.; Wolfe, L.G.; Kamatkar, N.G.; Chapman, S.M.; Diener, J.M.; Courtney, A.J.; Leevy, W.M.; Shrout, J.D. Imaging and Analysis of Pseudomonas Aeruginosa Swarming and Rhamnolipid Production. Appl. Environ. Microbiol. 2011, 77, 8310–8317.
- Be’er, A.; Ariel, G. A Statistical Physics View of Swarming Bacteria. Mov. Ecol. 2019, 7, 9.
- Kearns, D.B. A Field Guide to Bacterial Swarming Motility. Nat. Rev. Microbiol. 2010, 8, 634–644.
- Walker, K.E.; Moghaddame-Jafari, S.; Lockatell, C.V.; Johnson, D.; Belas, R. ZapA, the IgA-Degrading Metalloprotease of Proteus Mirabilis, Is a Virulence Factor Expressed Specifically in Swarmer Cells. Mol. Microbiol. 1999, 32, 825–836.
- Yan, J.; Monaco, H.; Xavier, J.B. The Ultimate Guide to Bacterial Swarming: An Experimental Model to Study the Evolution of Cooperative Behavior. Annu. Rev. Microbiol. 2019, 73, 293–312.
- Morgenstein, R.M.; Szostek, B.; Rather, P.N. Regulation of Gene Expression during Swarmer Cell Differentiation in Proteus Mirabilis. FEMS Microbiol. Rev. 2010, 34, 753–763.
- Toguchi, A.; Siano, M.; Burkart, M.; Harshey, R.M. Genetics of Swarming Motility in Salmonella Enterica Serovar Typhimurium: Critical Role for Lipopolysaccharide. J. Bacteriol. 2000, 182, 6308–6321.
- Damton, N.C.; Turner, L.; Rojevsky, S.; Berg, H.C. Dynamics of Bacterial Swarming. Biophys. J. 2010, 98, 2082–2090.
- Lai, S.; Tremblay, J.; Déziel, E. Swarming Motility: A Multicellular Behaviour Conferring Antimicrobial Resistance. Environ. Microbiol. 2009, 11, 126–136.
- Poudel, S.; Giannone, R.J.; Farmer, A.T.; Campagna, S.R.; Bible, A.N.; Morrell-Falvey, J.L.; Elkins, J.G.; Hettich, R.L. Integrated Proteomics and Lipidomics Reveal That the Swarming Motility of Paenibacillus Polymyxa Is Characterized by Phospholipid Modification, Surfactant Deployment, and Flagellar Specialization Relative to Swimming Motility. Front. Microbiol. 2019, 10, 2594.

