The life and proper functioning of the human body would not be possible without having extensive bacterial flora, which is present not only in the digestive and reproductive systems but also on the skin, which primarily has protective functions and is responsible for maintaining the proper homeostasis of the body. The skin microflora, also termed the skin microbiome, varies according to the sex and age of the host and is topographically diverse due to anatomical differences in the human body. The development of the microbiome on the skin in different bodily regions mainly depends on oxygen availability, the presence of sebaceous glands, humidity, and the temperature of the individual zones. It includes not only bacteria but also viruses, fungi, and bacteriophages [
1,
2]. The basic barrier, which is the skin, protects against mechanical and thermal factors, harmful radiation, and infections. The disruption of skin homeostasis, in combination with pathogen appearance, may lead to the development of various skin infections. Infection development is fostered by various factors, which include being immunocompromised, vascular insufficiency, poor lymphatic or venous drainage, sensory neuropathies, diabetes, obesity, poor hygiene, and surgical procedures [
3]. Infection occurrence is also influenced by the virulence of microorganisms and the degree of their presence. The simplest division of bacterial infections is acute and chronic infections. Pathogenic bacteria cause a wide variety of acute skin infections with varied clinical appearances depending on the aetiological factor, location, and immune status of the host. The most common acute infections are staphylococcal, streptococcal, and mixed staphylococcal–streptococcal infections [
4,
5]. In addition, other species also cause skin infections, including
Borrelia spp.,
Pseudomonas aeruginosa,
Corynebacterium spp.,
Actinomyces spp.,
Bacillus anthracis,
Listeria monocytogenes,
Nocardia spp.,
Erysipelothrix rhusiopathiae,
Bartonella spp.,
Klebsiella pneumoniae,
Klebsiella rhinoscleromatis,
Yersinia spp.,
Clostridium spp.,
Francisella tularensis,
Brucellae spp., and
Mycobacterium spp. [
6]. Moreover, bacteria that would not be pathogenic with an undamaged skin barrier can penetrate through damaged skin and become virulent. In general, most acute infections are superficial and relatively easy to treat with appropriate antibiotics [
7], whereas chronic infections, in particular chronic wound infections, often pose many difficulties in effective treatment, and in the era of multi-drug resistance, they are a major social and economic problem [
8]. The most common aetiological factors in chronic wounds are
Staphylococcus aureus,
P. aeruginosa,
Acinetobacter baumannii,
Proteus mirabilis,
Escherichia coli, and
Corynebacterium spp. [
9,
10,
11]. Among them, there are many antibiotic-resistant species included in the “ESKAPE” cluster composed of
Enterococcus faecium, S. aureus,
K. pneumoniae,
A. baumannii, P. aeruginosa, and
Enterobacter species [
12]. Due to the increasing drug resistance of bacteria responsible for skin infections, various studies have concentrated on the treatment of infections of wounds using various methods to reduce virulence factors [
13,
14,
15]. Some virulence factors include biofilm formation, exotoxins and endotoxins, proteases, quorum sensing, and motility [
14,
16].
The basic methods for treating chronic wounds are the use of antiseptics, a great variety of wound dressings, antibiotic therapy, compression therapy, and debridement. Other popular treatment options are hyperbaric oxygen therapy, negative pressure wound therapy, ultrasound, and electromagnetic therapy [
20]. Hyperbaric oxygen therapy is widely in use for more than 20 years in diabetic foot ulcers, and this therapy may improve wound healing by stimulating angiogenesis. The mechanism of action of hyperbaric oxygen therapy on tissues has not yet been fully elucidated [
21]. Another promising method for skin and wound treatment is negative pressure wound therapy, where the use of a vacuum reduces oedema and infection and increases local blood flow, which promotes healing [
22]. This method is widely used as an alternative to surgery for a wide range of wounds [
23]. Ultrasound therapy is the use of mechanical energy at high frequency (between 1 and 3 MHz) to induce thermal energy into musculoskeletal tissues and low frequency (between 22.5 and 40 kHz) to generate the cavitation effect, which, if delivered in waves, debride necrotic tissue from wound surface [
24]. Interestingly, skin wounds generate large and endogenous electric fields termed “current of injury” [
25]. This electrical skin activity is involved in the process of wound healing, so it has led to the hypothesis that applied electrical stimulation may support the healing of chronic wounds by imitating the natural electrical current [
26]. Several electrical stimulation methods are available, including pulsed current, direct current, and rhythmic electric frequency. However, the best strategy of electrical stimulation is still unachieved [
26]. Unfortunately, the above-mentioned methods frequently fail; thus, chronic wound treatment is difficult, long-lasting, and sometimes ineffective. As the problem of chronic wounds affects approximately 1–2% of the global population, innovative and more effective methods for treating hard-to-heal wounds are constantly being sought [
27,
28].
2. Types of Bacterial Movement
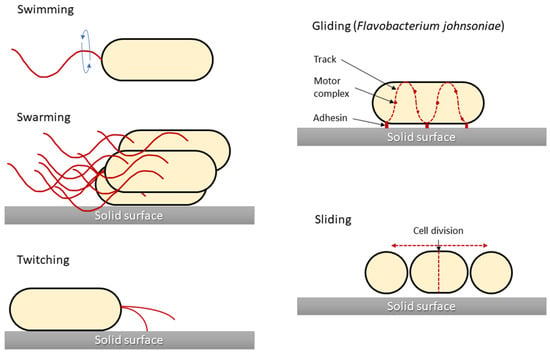
One of the main characteristics of microbes adapting to and surviving in the external environment is how they move and grow, whether in liquid, semi-liquid, or solid environments. Notably, not all bacteria are capable of active movement. In most cases, the flagella enable motion. Bacterial cells have flagella in variable quantities and locations, which is one of the features that enable their classification. However, different movement phenomena, such as the result of type IV pili (T4P), are still being investigated. In 1972, the different types of microbial translocation were characterised and named [
29]. Five of the original terms (swimming, swarming, twitching, gliding, and sliding) are still widely used [
30]. Differences in bacterial types of movement are shown in
Figure 1.
Figure 1. Graphical presentation of five bacterial movement types. Structures responsible for generating bacterial movement are marked in red. Sliding motility is presented for Flavobacterium johnsoniae as a model for this type of motility, although for other bacteria (i.e., Myxococcus xanthus) there are differences in the cell propulsion mechanism.
2.1. Swimming Motility
Swimming motility is one of the basic skills in the life cycle of bacteria, giving them an advantage in surviving unfavourable environmental conditions [
32]. Swimming motility is an individual cell movement that uses flagella rotation to move through aqueous environments [
33,
34]. Flagellated bacteria typically swim in a series of more or less straight runs interrupted by short reorientations [
35]. Each flagellum is rotated by a motor that is embedded in the cell membrane. Flagella are highly complex bacterial organelles that are unusually well conserved among diverse bacterial species. Over 60 structural and regulatory proteins are involved in flagellum synthesis and function [
36]. There are three elements in the structure of flagella found in prokaryotes: the fibre, the hook, and the basal body. A cell may have one, two, or more flagella on the front or back of the cell, over part of it, or covering its entire surface. The outer part of the cilia consists of the flagellin protein, of which there are approximately 20,000 subunits in a single cilium. The flagellum is anchored in the bacterial cell by a basic body consisting of the L, P, S, M, and C rings surrounding the cylindrical part. Gram-positive bacteria do not have the two outer L and P rings, which distinguishes them from Gram-negative bacteria with five rings [
37]. The flagella fibre is connected to the basic body with a flexible section called a hook, the length of which is approximately 55 nm [
38]. The direction and regulation of flagellar rotation enable bacteria to move in chemical gradients, termed chemotaxis [
35,
39]. This complex behaviour begins in cell membrane chemoreceptors, which detect chemical compounds and respond to them by changing their conformation.
E. coli has five such receptors (Tar, Tsr, Tap, Trg, and Aer), which are arranged in chemoreceptor clusters, together with two cytoplasmic proteins, the adaptor CheW and the histidine autokinase CheA [
40]. Motility regulation by the chemotaxis system is well investigated for a variety of bacterial species. Chemical gradients are sensed by chemoreceptors, which trigger the autophosphorylation of the cytoplasmic CheA, which forms a complex with the receptor through the coupling protein CheW [
33,
41]. In response to these changes, CheA transfers its phosphate group to CheY, a diffusible cytoplasmic response regulator that interacts with the flagellar motor and leads to a modulation of motility characteristics [
33,
41]. This signalling core is highly conserved among all chemotaxis pathways [
42].
Although only a fraction of bacteria associated with animal hosts are motile, flagellar motility and chemotaxis are important for the successful colonisation and virulence of many pathogens, for example, gastrointestinal
Campylobacter jejuni,
Salmonella enterica serovar Typhimurium,
Helicobacter pylori, and
Vibrio cholerae [
41]. In several cases, it has also been shown that the same flagellin, the protein subunit comprising the bacterial flagellum, can be a major driver of inflammation [
35].
Motility might have several functions in the host–microbe interactions. For example, in the early stages of biofilm formation, planktonic bacteria swim close to the surface by rotating their flagella and attaching to the surface using their pili [
43]. The most common opportunistic pathogen in wound infections is
P. aeruginosa. Many studies have demonstrated that
P. aeruginosa biofilms are the key factors for exacerbating the skin inflammatory response and its resistance to antimicrobial agents [
13,
36].
Bacteria are inextricably associated with wound healing. However, the latest research indicates that they are not always associated with infection and worse wound healing. It has been reported that the skin’s natural microbiome can contribute to more rapid wound healing. Several studies were performed with mice infected with staphylococci. Interestingly, S. aureus was found to be the best healing inducer.
Sometimes, swimming bacteria are used by non-flagellated bacteria that do not have the capacity to independently translocate with this mechanism. The genus
Staphylococcus, for example, is classically considered non-motile in fluid environments due to a lack of flagella. Despite their motility limitations, staphylococcal species effectively reach and thrive in their preferred ecological niches. Data suggested that
S. aureus has acquired, through
P. aeruginosa, an increased capacity to travel longer distances, allowing it to colonise niches that are relatively inaccessible in the absence of swimming carrier bacteria, which may be due to the hitchhiking of
S. aureus on
P. aeruginosa. It was also observed that
P. aeruginosa can carry another staphylococcal cargo,
Staphylococcus epidermidis, which may also be important in wound infection [
44].
2.2. Swarming Motility
Swarming motility is mainly based on the differentiation of vegetative cells into swarming cells with a large number of flagella, which can migrate rapidly in a coordinated manner over solid surfaces. Bacteria display different phenotypes during swarming, but not all of them are equal: swarming
P. aeruginosa do not develop typical swarmer cell phenotypes (elongated, hyper-flagellated cells) similar to other swarming bacteria, and individual cells swarming in rafts also reveal great variance and the lack of a highly differentiated swarmer cell phenotype [
45]. Swarming phenotypes vary not only within a species but also within a strain, whereby the different
P. aeruginosa strains grown on identical medium conditions will display different swarming patterns [
46]. A swarm of migrating bacteria moves forward and traps a water reservoir, and in this moist region, individual cell speed is comparable to swimming speeds in bulk liquid, typically in the order of 20 μm/sec [
47]. Bacterial swarming plays a crucial role in many pathogen–host interactions, and is considered an important virulence factor. The differentiation during swarming into the hyper-flagellated elongated cell of
P. mirabilis is coupled to the ability of this bacterium to enter host cells, with the manifestation of upregulated virulence protein expression (haemolysin, urease, and protease) [
48,
49].
The stimuli that are necessary to control this movement are responsive to cell density, surface contact, and physiological signals. The swarming motility needs signals from the quorum sensing system and the cyclic di-GMP network that regulates the transitions between motile and biofilm modes across many bacterial species [
50]. These signals activate flagella biogenesis via the main flagellar operon
flhDC, which is the main point of the regulatory network of cell differentiation and migration and is crucial for swarming motility [
48,
51]. Surface sensing occurs in two proposed ways, through the inhibition of flagellar rotation and/or through the detection of the O antigen contact of lipopolysaccharide (LPS) with a solid surface [
51] in Gram-negative bacteria. In addition, LPS is suggested to act as an osmolarity agent and facilitate swarming [
52].
The swarming phenomenon is characterised by bacterial growth in the form of zones with clearly delimited darker peripheral circles on an agar medium. In a typical swarming, after contact with a surface, microbial cells are able to extend up to 40 times and increase the number of flagella. In the following stage, cells divide into short cells that are unable to move. The cycles of cell elongation, movement, and division are repeated constantly, resulting in a characteristic “pattern” of their growth on the substrate, in which the darker rings reflect the periods of bacterial division. Changes in cell length and shape result from changes in the hardness of the surface on which the cells grow, and the presence of LPS, which participates in the regulation of osmolarity, helps them overcome the environmental barriers that inhibit bacterial mobility. Among others,
E. coli,
Salmonella spp.,
P. mirabilis,
Bacillus subtilis, and
P. aeruginosa are able to migrate in this way. Compared with swimming, chemotaxis is unnecessary here, and the oscillating motion is mainly based on the flagellar drive and the mechanical interactions taking place [
53]. The swarming motility is also closely associated with the resistance of bacterial cells, which are antibiotic-sensitive, providing cells with greater availability of nutrients and competitive advantage due to secreted surfactants [
48]. For example, the multi-resistant phenotype of
P. aeruginosa is closely associated with swarming growth and has a transient form [
54]. Currently, research on this motility is focused on obtaining more detailed information about their lipids, proteins, and enzymes, as well as their relationships, the metabolic pathways involved, and methods of regulation at the molecular level [
55].