Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Laura-Marinela AILIOAIE | -- | 6489 | 2023-07-05 14:53:09 | | | |
| 2 | Alfred Zheng | Meta information modification | 6489 | 2023-07-06 05:49:49 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Ailioaie, L.M.; Ailioaie, C.; Litscher, G. Infection and Inflammation Interplay in Pediatric Long COVID. Encyclopedia. Available online: https://encyclopedia.pub/entry/46463 (accessed on 27 July 2026).
Ailioaie LM, Ailioaie C, Litscher G. Infection and Inflammation Interplay in Pediatric Long COVID. Encyclopedia. Available at: https://encyclopedia.pub/entry/46463. Accessed July 27, 2026.
Ailioaie, Laura Marinela, Constantin Ailioaie, Gerhard Litscher. "Infection and Inflammation Interplay in Pediatric Long COVID" Encyclopedia, https://encyclopedia.pub/entry/46463 (accessed July 27, 2026).
Ailioaie, L.M., Ailioaie, C., & Litscher, G. (2023, July 05). Infection and Inflammation Interplay in Pediatric Long COVID. In Encyclopedia. https://encyclopedia.pub/entry/46463
Ailioaie, Laura Marinela, et al. "Infection and Inflammation Interplay in Pediatric Long COVID." Encyclopedia. Web. 05 July, 2023.
Copy Citation
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) in children and adolescents has generated repercussions, especially a few weeks after infection, for symptomatic patients who tested positive, for asymptomatic ones, or even just the contacts of an infected person, and evolved from severe forms such as multisystem inflammatory syndrome in children (MIS-C) to multifarious clinical manifestations in long COVID (LC). Referred to under the umbrella term LC, the onset of persistent and highly heterogeneous symptoms such as fatigue, post-exertion malaise, cognitive dysfunction, and others have a major impact on the child’s daily quality of life for months.
long COVID
infection
inflammation
children
1. Introduction
For the past four years, humanity has lived under the sign of uncertainty, infectivity, and mortality generated by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) that first appeared in Wuhan, China, in late 2019 and generated the coronavirus disease 2019 (COVID-19), responsible for the COVID-19 pandemic. Along with COVID-19 (2020 to present), there have been previous outbreaks that have been classified as public health emergencies of international concern by the World Health Organization (WHO), whose stimuli for the adoption of the 2005 International Health Regulations (IHR) as a treaty with 196 states were the following: the 1999–2000 influenza outbreak in Israel, the 2003 severe acute respiratory syndrome (SARS) pandemic, the 2009 H1N1 influenza pandemic, the 2014 Ebola outbreak, the 2015 Middle East respiratory syndrome (MERS) outbreak, poliomyelitis (2014 to present) [1][2][3].
Assumed as a relative of SARS and MERS, the SARS-CoV-2 betacoronavirus set the COVID-19 pandemic in motion by infecting primarily the lower respiratory tract and manifesting as pneumonia in humans. Also popularly known as the Wuhan seafood market pneumonia coronavirus, it is a positive-sense single-stranded ribonucleic acid (RNA) virus that infected the first symptomatic patients on 8 December 2019. WHO identified the first suspected cases three weeks later by genomic sequencing after RNA testing on samples taken from a patient with pneumonia during the initial outbreak in Wuhan. On 30 January 2020, the WHO declared this disease a Public Health Emergency of International Concern and characterized the outbreak as a pandemic on 11 March 2020. In the last century, and especially in the last three decades, an extensive percentage of viral outbreaks have been caused by RNA viruses, such as influenza, dengue, Zika, Ebola, and so on. Historically, unimaginable features have been discovered about the core replication machinery put into action by coronaviruses as a paradox among positive-strand RNA viruses because they use both a processivity factor and possess proofreading activity that tends to resemble deoxyribonucleic acid (DNA) organisms, in addition to the constitutive proteins involved in prompt RNA synthesis, often exploited by negative-strand RNA viruses [4][5][6][7][8][9].
The impact of COVID-19 on public health was considered severe, with the potential for community and international spread, so to reduce transmission, countries have put in place “stay at home” orders, strict lockdowns, and restrictions on public liberties. Although in children the infection usually resulted in mild illness, there has also been the rare complication of multisystem inflammatory syndrome in children (MIS-C) requiring emergency services among pediatric patients to save their lives [10].
The 2005 IHR is a valuable instrument of international law that is legally binding on many countries to build the capacity to identify and report potential public health emergencies worldwide. Investing WHO with more official authority and a proper understanding of the potential of IHR will strengthen global health security—a huge investment in human and animal health—while mitigating the possible extensive economic effects of the next global health emergency. The actual extent and trend of emergency department (ED) utilization, particularly in pediatrics, depends on both the epidemic disease and the mandated public health measures and how these affect decision-making, so policy makers need to be aware of the fact that fear of the virus in the general population certainly affects the response to public health advice, particularly in children, where there is a great inequity in considering the impact of the epidemic, which needs to be addressed in the future [1][10][11].
Many driving factors such as globalization, increased trade, urbanization, changing human behavior, and increased prevalence of viral diseases among animals can generate such pandemics as COVID-19, demonstrating that under the above-mentioned circumstances, viral diseases could easily spread between nations, having an impact on their economic stability. The COVID-19 pandemic has had an important global mortality rate and has presented a truly new challenge for medical systems worldwide to control this event quickly and correctly with multiple unforeseen consequences and harmful repercussions. It led to a special socioeconomic break due to the strict protection measures adopted by the authorities. The latest novel pandemic virus, i.e., SARS-CoV-2, has been shown to differ from the other two previous human coronaviruses related to genome structure, site of infection, transmissibility, and other important characteristics, all of which are implicated in its different lethal pattern. Like other RNA viruses, SARS-CoV-2, while adapting to its new human hosts, was susceptible to genetic evolution with the development of mutations over time, resulting in mutant variants, some of which were considered variants of concern by WHO, with an important impact on global public health. Many elderly or chronically ill people lost their lives, but also young people and children through the so-called “cytokine storm” (CS), an abnormal response of the host’s immune system (IS), characterized by an excessive discharge of proinflammatory cytokines/chemokines, which is activated by the SARS-CoV-2 virus with all its multiple and unpredictable clinical manifestations, and which could not be effectively managed with the drugs available at the beginning. Molecular mimicry allows this virus to affect the self-tolerance of the host’s IS, and the efforts of IS to eliminate SARS-CoV-2 can trigger autoimmunity by hyperactivating both the innate and adaptive ISs. This IS hyperactivity and production of autoantibodies after COVID-19 can trigger autoimmune diseases [12][13][14][15][16][17][18].
2. The Interplay between Main Molecular and Cellular Pathogenic Mechanisms in Pediatric Long COVID
Currently, for both adults and children, people still lack clear evidence about the factors involved and the pathophysiological mechanisms of the onset of LC symptoms. There are various theories and many hypotheses, but these are still limited by the heterogeneity of the available publications, so the proposed underlying mechanisms remain ambiguous. In general, it is believed that the long-lasting clinical symptoms are the result of viral aggression in the acute phase of the disease because of damage to the involved systems and organs, followed by the persistence of the SARS-CoV-2 or viral components, which, due to immune dysregulation, cannot be eliminated and which trigger autoimmunity phenomena, endothelial dysfunction, microcirculation disorders, and activation of the coagulation system [19][20].
2.1. Persistence of Virus or Viral Components and Prolonged Immune Activation with Inflammation
The initiation of acute SARS-CoV-2 infection is favored by the presence of the ACE2 receptors as a gateway for SARS-CoV-2. Through this gate, the virus enters and attacks the cells of the nasal cavity, oral mucosa, lungs, heart, GI tract, pancreas, liver, spleen, kidneys, brain, vascular endothelium, and others, causing damage manifested by acute symptoms, which may later become persistent or recurrent. The spike protein (S) in the structure of the coronavirus facilitates the penetration of the viral agent into the target cells. Attachment is achieved by the S1 subunit of the spike protein, which is responsible for recognizing the cell surface receptor ACE2 through a receptor binding domain (RBD), while the S2 subunit is required for membrane fusion. If the cell has fewer ACE2 receptors on its surface, the infection may be milder. Transcriptomic studies have shown that children have lower ACE2 protein expression than adults, although the extent of gene expression does not always correlate with protein levels. ACE2 receptors are encoded by a gene that can have several forms, that is, there is a polymorphism that varies from person to person. The most common polymorphisms of the angiotensin-converting receptor gene are the angiotensin-converting enzyme insertion (ACE I) and the angiotensin-converting enzyme deletion (ACE D) polymorphism [21][22][23].
For the virus to enter the cell, it is necessary to split the S protein by the proteases of the attacked cell into the S1/S2 and S2’ components. The process of fusion of virus and cell membranes is driven by the S2 subunit that binds ACE2 to be the entry receptor and uses the transmembrane protease serine 2 (TMPRSS2) to prime the S protein. A TMPRSS2 inhibitor approved for clinical use was shown to be able to prevent virus entry, an aspect that brings hope for treatment [24][25].
ACE2, in addition to being the cellular receptor for SARS-CoV-2 and other coronaviruses, plays an essential role in the regulation of inflammatory processes by decreasing the main proinflammatory peptides of the renin–angiotensin system (RAS) and the kinin–kallikrein system (KKS). These two interconnected signaling systems stimulate the inflammatory response through tumor necrosis factor alpha (TNF-α), which in turn is activated by tumor necrosis factor–alpha converting enzyme (TACE) also known as a disintegrin and metalloprotease 17 (ADAM17) protease that destroys ACE2 and recruits macrophages and neutrophils. ACE2 would have an important role in stopping this inflammatory process. If ACE2 is no longer activated or has undergone degradation, an inflammatory response will be triggered with disastrous consequences on organs, including the onset of pulmonary edema from COVID-19, but at the same time it can have long-term repercussions even after the SARS-CoV-2 infection has ceased, as observed in MIS-C. Regarding ACE2 expression in adults and children, there are conflicting opinions, but it is accepted that ACE2 expression does not change uniformly with age, starting from late fetal life, where ACE2 expression is low, and then reaching a maximum in early childhood, with large cellular and individual variations, and decline in old age. The reduction in ACE2 expression with age and the amplification of the proinflammatory components of the RAS may have a special role in supporting the inflammatory theory of the aging phenomenon and in the senescence processes. Since ACE2 is located in the X chromosome and its experimentally demonstrated expression is higher in females than in males, it may argue for the proposed negative correlation in the gender comparison. The inverse relationship between ACE2 expression and severity of COVID-19 is an exception in children, who manifest milder forms of COVID-19. This could be explained by the fact that children express less ACE2 than adults, but much more angiotensin II receptor type 2 (ATR2), which competes with angiotensin II receptor type 1 (ATR1) and blocks it, preventing its inflammatory effect. Therefore, it is believed that the high level of ATR2 receptors in children can counterbalance the low level of ACE2, and, as a result, clinical symptoms are moderate in children [26][27][28][29][30].
The extensive surface expression of the ACE2 receptors on lung alveolar epithelial cells, small intestine enterocytes, arterial and venous endothelial cells, and arterial smooth muscle cells in various organs could clarify the multitude of clinical manifestations of LC, after infection with SARS-CoV-2. Long-term effects of SARS-CoV-2 infection also occur in patients with moderate and mild forms of COVID-19, including brain fog, reduced gray matter thickness, and brain size. Research on the neuropathological characteristics of the brains of deceased patients revealed mild neuropathological changes and pronounced neuroinflammation in the brainstem. Activation of microglia and pronounced infiltration of brainstem tissue, cerebellum, and meningeal structures with cytotoxic T lymphocytes was observed in patients who died of COVID-19. Histoimmunochemical examination demonstrated the presence of SARS-CoV-2 RNA and proteins in both cranial nerves from the lower brainstem and in isolated brainstem cells from several cases who died of SARS-CoV-2 [31][32][33].
A growing body of evidence is being brought forward by researchers demonstrating that in some LC patients, the virus persists in hidden reservoirs in tissues or organs, including central nervous system structures, after acute infection [34][35].
Accumulation of SARS-CoV-2 spike protein in the skull–meninges–brain axis presents potential molecular mechanisms and therapeutic targets for neurological complications in LC patients, as a team of researchers very recently proved. Rong et al. investigated in mouse models and postmortem human tissues the presence and distribution of the SARS-CoV-2 spike protein in the skull–meninges–brain axis. The study revealed that the accumulation of spike protein in the marrow of the cranial bones and meninges for a long time after the onset of COVID-19 and then the penetration into the cerebral circulation and brain structures may participate in the emergence of the neurological manifestations of long COVID. These findings suggest that virus fragments accumulate and remain in the brain long after the virus is gone and trigger the inflammation that causes symptoms of long COVID [36].
In another study, although the presence of the virus in the brain parenchyma was not detected, the signs of an immune activation process translated by significant and persistent neuroinflammation in patients with acute COVID-19 were highlighted; it is believed that the highly immunogenic spike protein could be directly involved in the perpetuation and aggressiveness of the infection [37].
Recent publications have highlighted the presence of perivascular inflammation in the brains of patients who died of COVID-19, and others have demonstrated that spike protein could damage the vascular endothelium in an animal model and disrupt an in vitro blood–brain barrier (BBB) model, facilitating its penetration through the BBB and the occurrence of neuroinflammation. On the other hand, the spike protein seems to work together with human molecular chaperones, triggering a process of autoimmunity and the stimulation of toll-like receptors (TLR), with the release of proinflammatory cytokines. Some anti-protein antibodies cannot be neutralizing and change their conformation to bind to its receptor. Once the spike protein enters the brain or is expressed by brain cells, alone or together with inflammatory cytokines, it could stimulate microglia and initiate neuroinflammation in LC [38][39].
Prolonged or recurrent olfactory and gustatory dysfunctions are common in COVID-19, especially in patients with mild symptoms. An etiological, virological, molecular, and cellular study of the olfactory neuroepithelium in seven patients with COVID-19 who presented with acute loss of smell demonstrated the presence of viral transcripts and cells infected with SARS-CoV-2, along with phenomena of chronic local inflammation [40][41].
Neurological pathology as expressed by encephalitis, stroke, acute transverse myelitis, and cranial or peripheral nerve damage, such as Guillain-Barré syndrome, represents approximately 1% of cases reported in children. Some manifestations may occur during or after infection with SARS-CoV-2, through immune-mediated post-infectious mechanisms [42].
2.2. Processes Mediated by Autoantibodies in the Spectrum of Autoimmune Diseases
After the acute phase of the infection, complete removal of the SARS-CoV-2 and increased immunity will not necessarily lead to complete elimination of viral RNA, as virus- and host-related mechanisms intervene. The virus can protect itself by mutations of the genes that encode the junction or replication proteins, as well as by its ability to evade the action of the adaptive immune system. The host can defend itself by using non-cytolytic clearance mechanisms that protect against destruction of infected cells, which are specifically activated by innate immune responses in an attempt to abrogate virus replication in infected cells. Identifying the role of RNA persistence in protracted disease, how it is protected from degradation, its relative length and fragmented or defective viral genomes (DVGs), and how RNA is replicated will help us solve this mystery and possible therapeutic interventions [34].
Several months later, after infection with SARS-CoV-2, the persistence of viral messenger RNA (mRNA) and spike protein was evidenced in LC patients by positive PCR in the digestive and genitourinary tracts, by intestinal biopsy, and histopathological examination by microscopy electron and immunofluorescence. At the same time, it has been shown that the GI tract, especially in children, is a site of active viral replication of SARS-CoV-2 and can function as a long-term reservoir for this virus [43][44][45][46][47][48][49].
An immunohistochemistry and electron microscopy study of skin biopsies from seven children and adolescents with skin lesions and COVID-19 confirms vascular lesions and the presence of SARS-CoV-2 in endothelial cells. Vascular injury, inflammation, and chemotaxis persisted in patients with COVID-19 and were correlated with abnormal clinical features three months after discharge, particularly in severely recovered COVID-19 patients [50][51].
Other studies have shown that after acute infection with SARS-CoV-2, children continue to have the virus in their feces for a long time compared to adults, despite negative breath tests [52][53].
A very recent publication reported that spike fragments and/or viral RNA persist in post-COVID-19 recovered patients up to 1 year or more after acute SARS-CoV-2 infection and indicated that circulating spike protein is associated with extracellular vesicles in the absence of viral RNA inside these vesicles [54].
Extracellular vesicle (EV) is actually an umbrella name for a diversity of nano-sized particles (30–1000 nm) secreted by cells, which can consist of exosomes, cell membrane-derived microvesicles, and apoptotic bodies. EV has a lipid bilayer membrane, which gives it the ability to bind artificial ligands for various targets or incorporate microRNAs (miRNAs), messenger RNA (mRNA), DNA, lipids, biologically active compounds, or various proteins. Their value lies in the fact that they are biodegradable, biocompatible, allogeneic, and safe; therefore, they can be used as therapeutic platforms for antiviral and as anti-inflammatory and anticancer agents. The host cell infected with SARS-CoV-2 can escape the spike protein with the help of these EVs [55][56][57].
After the first 6 months of life, when the infant begins to lose the maternal protective antibodies that ensure passive immunity, he or she must face increasingly numerous external antigenic aggressions. Viral invasion of the upper airways with SARS-CoV-2 in children is counteracted by a strongly marked response through type I and type II interferon (IFN), as well as through inflammasome-dependent pathways, compared to the response mode in adults. IFN-I plays a crucial role by activating the genes that trigger the apoptosis of infected cells, the stimulation of which is earlier and more effective in children due to the multiple receptors in the cytosol, compared to the elderly, in whom this production is late, generating an insufficient tardy immune response that already allows the replication of the viral agents in the cascade. The appearance of anti-IFN autoantibodies is correlated with age; they will increase the severity of the disease by delaying the prompt reaction of the immune system against the virus, which explains why patients with severe form of COVID-19 have a high percentage of autoantibodies targeting IFN, complement, interleukins, chemokines, and other immunomodulatory proteins, including those on the cell surface. The key issues raised by the very different reactions and disease forms in COVID-19 and LC must be sought in correlation with the peculiarities of the children’s immune system, which can be a “successful immune response” compared to adults and the elderly. It was found that in small children the respiratory tract is stronger, and it defends itself much better, probably due to the training received at the frequency of viral infections and/or recent vaccinations and the changes in the local microbiome. More efficient local tissue responses, better thymic function, and reactive immunity have been proposed to explain mild clinical symptoms or even asymptomatic forms of SARS-CoV-2 infection in children [58][59][60][61].
After the COVID-19 pandemic, an increase in the number of cases of autoimmune disease is reported. The mechanism by which SARS-CoV-2 can induce these pathologies is represented by the overactivation of mature natural killer cells and CD8+ T lymphocytes, the dysregulation of B lymphocytes, and the participation of proinflammatory cytokines. It was shown that patients with COVID-19 had increased concentrations of TNF-α, interleukins IL-1 and IL-6, and leukocytes and neutrophils, whereas the number of lymphocytes, monocytes, eosinophils, and basophils was decreased [62][63].
Molecular mimicry would be one of the fundamental mechanisms by which peptide structures in the infectious agent SARS-CoV-2 similar to those of the host can activate T or B lymphocytes and trigger the phenomenon of autoimmunity, but molecular mimicry is not the only mechanism in the development of autoimmune responses. Many other factors can participate in this phenomenon, among which it mention the genetic structure of the host, the interaction with the microbiota, chemical adjuvants, disruption of immune tolerance, nonspecific activation with the persistence of some cryptic external antigens, the response of autoreactive T lymphocytes, the activation of autoimmune B cells, etc. [64][65][66].
The distorted immunological response is a key factor in the LC phenomenon in children and adolescents, as demonstrated in a recent study on 12 children with PASC (or LC) compared to 17 patients completely recovered after SARS-CoV-2 infection. In the studied group with PASC, three patients had pre-existing comorbidities (bronchial asthma, Schőlein-Henoch purpura, and Duchenne muscular dystrophy without oxygen demand)). The laboratory data objectified higher concentrations of plasmablasts, IgD-CD27+ memory, and switched IgM-IgD-B cells. Children from the control group presented significantly higher naïve and unswitched IgM+IgD+ and IgM+CD27-CD38dim B cell subsets. The serum concentrations of IL-6 and IL-1β were increased in both groups, but much higher in children with PASC, evoking the predominant participation of the innate immune system [67].
The study of IgM and IgG antibodies against the RBD of the spike protein of SARS-CoV-2 showed a significantly decreased concentration after 6 months of infection with SARS-CoV-2, whereas the number of RBD-specific memory B cells remained unchanged. This aspect can be explained by the persistence of a small amount of SARS-CoV-2 antigen or incomplete viral clearance. The hypothesis was confirmed by the presence of SARS-CoV-2 RNA and proteins in 7 of 14 intestinal biopsy samples from previously asymptomatic COVID-19 patients with a negative nasal swab PCR after a period of 4 months after acute infection [68].
Persistence of the SARS-CoV-2 or its components can induce LC symptoms by activating the host’s perverted immune response, which will trigger the generation of long-term autoantibodies that target interferon responses, type II cellular immunity, the acute phase response, leukocyte movement, and lymphocyte function or activation.
Autoantibodies directed against the endothelial adhesion molecule plasmalemma vesicle-associated protein (PLVAP), the regulator of angiogenesis r-spondin-3 (RSPO3), the metabotropic glutamate receptor, and the hypocretin receptor (orexin) receptor 2 gene responsible were detected of sleep disorder, in the central nervous system and against connective tissue, and of matrix metalloproteinases MMP7 and MMP9 [69][70].
Patients with symptoms after 3 months of acute illness more commonly show increased levels of proinflammatory cytokines (e.g., IL-1, IL-6, TNF-α), chemokines such as monocyte chemoattractant protein-1 (MCP-1) and interferon gamma-inducible protein-10 (IP-10), but also factors associated with vascular damage, such as intercellular adhesion molecule 1 (ICAM-1) and vascular cell adhesion molecule 1 (VCAM-1) [71].
Patterson et al. set out to research the presence of the SARS-CoV-2 S1 protein in a group of 144 people divided as follows: 29 normal individuals, 26 patients with mild-moderate COVID-19, 25 patients with severe COVID-19, and 64 of patients with chronic post-COVID-19 symptoms. Persistence of symptoms has been suggested to be secondary to immune stimulation by viral RNA and spike protein present in a subset of monocytes. From the initial batch, the authors investigated the presence of the SARS-CoV-2 S1 protein in 46 people. T, B lymphocyte subsets and monocytes from patients with severe COVID-19 and from those with persistent symptoms after acute infection with COVID-19 were studied. The number of intermediate (CD14+, CD16+) and nonclassical (CD14Lo, CD16+) monocytes was significantly increased in patients with PASC symptoms persisting 15 months after acute infection, compared to the control group of healthy subjects. At the same time, a significant percentage of nonclassical monocytes containing the SARS-CoV-2 S1 protein was detected in patients with severe COVID-19 but also in PASC patients 15 months after infection with SARS-CoV-2 [72].
As is known, circulating monocytes are classified into three subsets (classical, intermediate, and nonclassical) and have different functions in the immune system. Classical monocytes are involved in phagocytic activity, secrete the cytokines IL-6, IL-8, chemokine C-C motif chemokine ligand 2 (CCL2), CCL3, and CCL5, and produce large amounts of reactive oxygen species (ROS). Monocytic cells from the intermediate subset express C-C chemokine receptor type 5 (CCR5) on their surface, are antigen presenters, secrete cytokines TNF-a, IL-1β, IL-6, and chemokine CCL3 after stimulation of TLRs. The subtype of nonclassical monocytes presents on their surface high levels of CX3C motif chemokine receptor 1 (CX3CR1), participates in the phagocytosis process mediated by complement and crystallizable fragment (Fc) gamma, and is responsible for antiviral responses. After release from the bone marrow into the systemic circulation, 85% of human monocytes are of the classical subtype, and the remaining 15% are intermediate and nonclassical. The activation of nonclassical monocytes in patients with COVID-19 and the presence of S1 protein in patients with PASC indicate their role as a viral protein reservoir. In patients with PASC, high levels of the cytokine interferon-gamma (IFN-γ), which can stimulate the production of TNF-α, have been detected. High levels of TNF-α and IFN-γ induce the upregulation of the chemokine fractalkine (CX3CL1) by vascular endothelial cells, favoring the survival of nonclassical monocytes. Thus, nonclassical monocytes are perverted and acquire a proinflammatory function by activating the pathogenic nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) pathway and stimulating the production of proinflammatory cytokines IL-1α, TNF-α, and IL-8 [73][74][75][76].
SARS-CoV-2 infection worsened psychological status, especially among children and adolescents with long-term neuropsychiatric consequences. During the acute episode, there was worsening of obsessive disorders, anxiety, restlessness, mania, anger, and tics, with decreased cognitive performance and well-being. After COVID-19, more new cases of pediatric acute-onset neuropsychiatric syndrome (PANS) and even other symptoms have emerged. In explaining this pathology, silent viral agents such as Epstein–Barr virus have been hypothesized to be involved in pathogenic mechanisms through reactivation and immune responses with chronic neuroinflammation. Epstein–Barr virus is a pathogen that is part of the human lymphotropic herpesviruses (family Herpesviridae) that can affect approximately 95% of the global population. In total, 90% of adults have been shown to have anti-EBV antibodies, evidence of previous infection. Primary EBV infection is much more common in children and adolescents, usually expressed by atypical or asymptomatic manifestations, misdiagnosed or missed. Children under four years of age, immunosuppressed, or immunodeficient may not react with specific antibodies. In other age groups, the virus sometimes cannot stimulate the body strongly enough to produce enough antibodies to be detected. If EBV-specific viral antibodies cannot be demonstrated, then DNA testing is a technique used more and more often in the clinic. PANS is a model of expression of immune-mediated neuropsychiatric symptoms, as reactivation of a latent virus and a prototype of chronic post-acute manifestations of COVID-19. EBV reactivation can occur after infection with SARS-CoV-2 or concurrently with COVID-19, including initially asymptomatic patients. SARS-CoV-2 can train other infectious agents (e.g., cytomegalovirus) to participate in the long-term symptoms of COVID. More and more studies claim that long-term symptoms of COVID are a consequence of inflammation caused by SARS-CoV-2 or its components through reactivation of EBV. SARS-CoV-2 can stimulate subclinical chronic inflammation. Chronic recurrent/reactivated EBV infections are accompanied by the onset of several autoimmune diseases, including diabetes. If chronic or recurrent EBV infection affects epithelial cells, systemic lupus erythematosus and Sjögren’s syndrome can occur, and when B cells are infected, chronic arthritis, multiple sclerosis, and other diseases can occur. These secondary autoimmune diseases present as overlapping syndromes with clinical and biological manifestations expressed by specific autoantibodies (e.g., antinuclear antibodies and rheumatoid factors) that reflect epithelial and/or B-cell infection [77][78][79][80][81].
Other secondary manifestations of SARS-CoV-2 infection are those that affect the muscular system and the locomotor apparatus present in a percentage of 10–25% of patients with COVID-19. Suffering is manifested by asthenia, myalgia, joint pain, back pain, etc. Myalgia, protracted myositis, rhabdomyolysis, and “autoimmune necrotizing myositis” induced by SARS-CoV-2 have been reported by many publications in patients with COVID-19. During infection, release of proinflammatory cytokines with myotoxic potential, deposition of immune complexes, and damage caused by similarities between viral antigens and human muscle cells are considered immune-mediated mechanisms of muscle damage. Cachexia and sarcopenia may occur as muscle sequelae of long-term post-COVID-19 evolution. Reduction in muscle mass by destruction of fibers and infiltration with adipose tissue are detected by magnetic resonance imaging (MRI) in the case of muscle atrophy from cachexia and sarcopenia [82][83][84].
During the COVID-19 pandemic, there has been a sharp increase in new cases of juvenile dermatomyositis (JDM) in older female children. Some infectious agents such as parvovirus B19, coxsackie virus B, enterovirus, influenza, group A streptococcus, and toxoplasma gondii are known to trigger seasonal JDM. At the same time, infection with SARS-CoV-2 can be a strong trigger for JDM. It seems that dysregulation of the type I IFNs signaling pathway can lead to the expression of myxovirus-resistance protein A with its deposition in capillaries and muscle fibers and could be one of the hypotheses in the pathogenesis of JDM. Prolonged post-viral myositis documented by MRI occurs following SARS-CoV-2 infection approximately 3–7 days with diffuse or multifocal muscle pain and/or rhabdomyolysis and may last much longer, which is another hypothesis. This last entity is a syndrome of hyperinflammation such as MIS-C, KD or dermatomyositis, but it is not true dermatomyositis [85][86][87][88].
In addition to dermatomyositis, other entities such as juvenile idiopathic arthritis, reactive arthritis, spondyloarthritis, psoriatic arthritis, and systemic lupus erythematosus are part of the chronic rheumatic diseases that could start after SARS-CoV-2 infection [89][90][91][92][93][94][95][96].
In recently published studies, it is mentioned the particularly high risk for patients who have had COVID-19 of developing after a few months autoimmune diseases such as systemic sclerosis, mixed diseases of connective tissue disease, Behçet’s disease, polymyalgic vasculitis, inflammatory bowel disease, celiac disease, type 1 diabetes, etc. Attention is drawn to the link between COVID-19 and the risk of these autoimmune diseases, so that they are recognized in time and patients can receive early treatment [66][97][98].
2.3. Long COVID in the Era of the Microbiome and Connected Molecular Mechanisms
Research and scientific knowledge about SARS-CoV-2 infection and its consequences on the human body developed gradually as the pandemic unfolded. Although the primary site of infection is in the respiratory system, SARS-CoV-2 can be found in the GI tract and has been isolated from stool samples with evidence of virus activity in the GI. The virus was detected in the esophagus, stomach, duodenum, and rectum and in fecal samples from COVID-19 patients. Long-term GI manifestations, especially diarrhea, have been linked to decreased richness and diversity of gut microbiota, immune deficiencies, and delayed virus clearance. Operating in two directions, the interactions between the respiratory mucosa and the gut microbiota, on the gut–lung axis, are supposed to be involved in the immune responses to infection. As in other respiratory infections, gut dysbiosis is linked to heightened inflammation and diminished regulatory or anti-inflammatory processes in both the lung and gut, highlighting this significant connection between the two mucosal compartments. Investigations have pointed to the occurrence of intestinal dysbiosis in infected patients, with exacerbated inflammation and decreased regulatory or anti-inflammatory mechanisms, as a possible interrelationship in terms of disease severity. An open question that has been raised is whether dysbiosis is a consequence or a contributing cause of the severity of SARS-CoV-2 infection, such that treatments aimed at decreasing GI permeability may be salutary [97][99][100].
The damage outside the lungs triggered by COVID-19 includes the gut, because the virus can remain present where ACE2 receptors are strongly represented. GI manifestations involve diarrhea, vomiting, and abdominal pain existing in many patients due to the tropism of SARS-CoV-2 for ACE2. Gut homeostasis and the microenvironment play a vital part in ameliorating systemic inflammation. There is still insufficient information on residual GI symptoms of SARS-CoV-2 infection. This virus clearly and indirectly affects gut physiology in various ways; consequently, different functional disorders of the gut will take place after recovery, such as dysbiosis, disruption of the intestinal barrier, microinflammation of mucosa, post-infectious conditions, immune abnormalities in the regulation of metabolism, and physiological or even psychological stress due to possible pathophysiological changes [101].
Diet, environmental factors, and genetics all participate as important players in shaping the gut microbiota that influence immunity [102].
It is known that among environmental factors, the microbiome modulates immune responses to infection. The boundary that separates the host tissue from this microbial community is constituted by the intestinal epithelium and the mucus layers, where immune cells continuously pick up antigens from the lumen, as well as from the destroyed mucosal barrier triggered by infection, the physical damage or inflammation of disturbed tissues, which facilitates direct interplay between host immune cells and the microbial congregation. Immunocompromised patients have a “leaky” gut due to chemotherapy or opportunistic infections, leading to important microbial translocation that precipitates abnormal proinflammatory immune cell responses. The mechanisms of the evolutionary origin of variants of concern for SAR-CoV-2 are still a matter of debate. Chronic infections with SARS-CoV-2 in certain people who are immunosuppressed is one of the hypotheses. For immunocompromised children and adolescents, permanent surveillance of the possible impact of variant strains is necessary, and vaccination must be carefully considered. [101][103][104][105][106][107].
The role of immunomodulation in the treatment of LC is more and more obvious. Clinical symptoms in LC are modulated by proinflammatory and effector physical and biochemical characteristics, as determined by both genetic makeup and environmental influences induced by SARS-CoV-2 infection. Evaluation of cytokines and lymphocyte subsets at a long distance of at least 7 months from acute infection with SARS-CoV-2 shows that latent autoimmunity and not hidden autoimmunity continued firmly throughout this time. Proinflammatory phenomena were highlighted by the increase in interleukins IL-1β, IL-6, IL-13, and IL-17A, as well as the concentrations of IFN-α, TNF-α, and granulocyte colony-stimulating factor (G-CSF), simultaneously with the reduction in IP-10, induced by interferon-γ. Many effector T lymphocytes, CD8+, T effector CD4+ with memory, and naive B cells were recorded in LC. Total IgG S1 anti-SARS-CoV-2 antibodies levels remained high over time, so that this long-term remaining immune activation can concur to the progress of latent or expressed autoimmunity. Chronic inflammatory processes with increased numbers of B cells, as primary source of autoantibodies, support autoimmune mechanisms in LC [108][109][110].
There is less known regarding the degree of gut microbiome dysbiosis in children relating to disease severity during COVID-19 infection. Higher frequency of asymptomatic cases in children, as well as the symptomatic forms of COVID-19, represents a very interesting open direction of research regarding microbiome differences, by comparison with adults. LC-associated gut microbiome dysbiosis revealed higher levels of Ruminococcus gnavus and Bacteroides vulgatus as well as lower levels of Faecalibacterium prausnitzii (F. prausnitzii) in adults. Analysis of stool in asymptomatic infants with COVID-19 revealed a decrease in Bifidobacterium bifidum, Akkermansia muciniphila, Eubacterium limosum, Enterocloster clostridioformis, Blautia hominis, Veillonella dispar, and Enterobacter cloacae, of which the first two are of particular importance as they are known for anti-inflammatory bacterial taxa. Bifidobacterium, as one of the most investigated for its positive effects could be used to modulate the intestinal TJ barrier and to protect and treat gut inflammation [111].
There is an urgent need to search and develop innovative treatments to successfully interact with the human microbiota and immune system in the coronavirus crisis. New solutions, molecules, and probiotics should be designed and investigated to be innovatively applied to discover how bacteria can help us fight this pandemic, so that people can find the key to the hidden code of communication between RNA viruses, bacteria, and our body [112].
Intestinal inflammation and gut barrier dysfunction as well as microbial translocation trigger changes in the gut microbiome. SARS-CoV-2 in the GI tract results in the release of zonulin, an important coordinator of intercellular tight junctions (TJs) between epithelial cells; higher levels of zonulin increase intestinal permeability (the “leaky gut”), which in turn facilitates trafficking of SARS-CoV-2 antigens in the blood and lymph flow, leading to hyperinflammation and autoimmune phenomena [97][113][114], as shown in Figure 1.
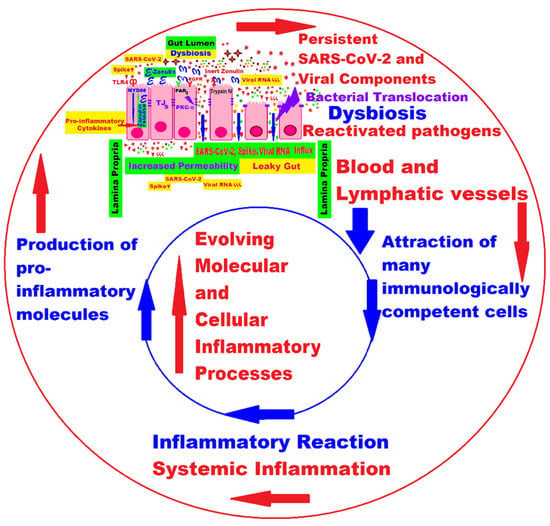
Figure 1. Persistence of SARS-CoV-2 and its remnants, intestinal dysbiosis, bacterial translocation, reactivation of pathogens, and their influence on the evolution of molecular and cellular processes in LC [97].
After acute infection with the SARS-CoV-2 associated with significant lung damage, there is a percentage of about 22% of patients who remain with GI symptoms, fatigue, sleep disturbances, mood disturbances, and cognitive dysfunction (“brain fog” and impaired memory) post-COVID-19. The mechanisms by which prolonged GI manifestations and neuropsychiatric symptoms occur in former patients with COVID-19 are unclear. From the data in the specialized literature, it can be observed that the modified structure of the intestinal microbiome, especially the reduction in butyrate and the increase in proinflammatory activities, are some of the causes of these disorders. In patients with chronic COVID-19, reduction in commensal bacteria with immunomodulatory power was observed, including F. prausnitzii, Eubacterium rectale, and Bifidobacteria strains; at the same time, a significant increase in proinflammatory taxa (for example, Prevotella and Veillonella) was detected in the oral microbiome [115][116][117][118].
Infection with the SARS-CoV-2 could alter certain microbial communities that disrupt the release of beneficial bacterial metabolites, subsequently inducing neurogenesis or neuroinflammation in central nervous system structures that facilitate the emergence of cognitive dysfunctions in LC. Persistent GI symptoms and chronic neuropsychiatric manifestations together with prolonged gut dysbiosis suggest impairment of the post-infectious microbiota–gut–brain (MGB) axis that could elucidate the cause of these symptoms in post-COVID-19 patients. Perturbation of the MGB axis of bidirectional communication between the intestinal environment and the brain can lead to various symptoms or diseases, including anxiety, cognitive deficits, and depression. Altered gut microbial colonization has been shown to be associated with behavioral disturbances in patients with irritable bowel syndrome, inflammatory bowel disease, metabolic disease (obesity, diabetes), autism in children, and neurodegenerative diseases (Parkinson, Alzheimer’s) in adults. Original, smart, and targeted multifunctional solutions and new nanodrugs may soon be developed to treat, for example, Alzheimer’s disease by targeting TJs not only at the brain level, but also at the gut level, with possible applications in long COVID in children, too [119][120][121].
Neuroinvasion of the brain by SARS-CoV-2 following intestinal infection and the zonulin hypothesis was highlighted. Zonulin, as a protein with an important role in the reversible regulation of epithelial permeability, released in large quantities in the intestinal lumen, can influence the integrity of TJs and the continuous efflux of antigens to the submucosa, generating inflammatory processes and severe immune reactions, with an effect on the severity of SARS-CoV-2 infection and triggering systemic inflammation. Among the possible pathways of neuroinvasion by SARS-CoV-2, an alternative pathway after gut infection, the zonulin hypothesis has been proposed, involving toll-like receptor 4 (TLR4), zonulin, protease-activated receptor 2 (PAR2), and the brain receptor zonulin [97][122][123].
Neurological and neuropsychological symptoms (fatigue, headaches, cognitive problems, and mood swings) post-COVID-19 are heterogeneous, but there is a similarity of brain patterns between children and adults, as can be seen from data published in the scientific literature. These manifestations could be explained by the hypometabolism of certain areas of the brain in long-term COVID-19. On the other hand, the dysfunction of the hypothalamic–pituitary–adrenal axis, modulated by the unbalanced gut microbiota, foreshadows that disturbed stress reactions may underlie the neuropsychiatric symptoms in LC. In pediatric pathology, neurological symptoms tend to decrease over time, and psychological symptoms remain present in large numbers for a long time in patients aged 6–17 years [124][125][126][127].
It is not yet known exactly whether the neuropsychiatric symptoms observed in children and adolescents with long COVID are the direct result of the infection or are due also to the stress caused by the restrictive measures during the pandemic and the illness itself. The neuroinflammation and the response molecules generated in the brain due to the persistence of the virus and its components could lead to an avalanche of phenomena that command the immune cells in the intestine, which should normally fight pathogens, but end up causing inflammation in the gut. Psychological assistance should be an important part in managing long COVID in children and adolescents [51][128][129].
References
- The World Health Organization. International Health Regulations (2005)–Third Edition. Available online: https://www.who.int/publications/i/item/9789241580496 (accessed on 10 April 2023).
- Gostin, L.O.; Katz, R. The International Health Regulations: The Governing Framework for Global Health Security. Milbank Q. 2016, 94, 264–313.
- Wilder-Smith, A.; Osman, S. Public health emergencies of international concern: A historic overview. J. Travel Med. 2020, 27, taaa227.
- Sohrabi, C.; Alsafi, Z.; O’Neill, N.; Khan, M.; Kerwan, A.; Al-Jabir, A.; Iosifidis, C.; Agha, R. World Health Organization declares Global Emergency: A review of the 2019 Novel Coronavirus (COVID-19). Int. J. Surg. 2020, 76, 71–76, Erratum in Int. J. Surg. 2020, 77, 217.
- Zhang, Y.; Yu, B.; Chen, X.; Rich, S.; Mo, Q.; Yan, H. Dynamics of the coronavirus disease 2019 (COVID-19) epidemic in Wuhan City, Hubei Province and China: A second derivative analysis of the cumulative daily diagnosed cases during the first 85 days. Glob. Health J. 2021, 5, 4–11.
- Worobey, M.; Levy, J.I.; Malpica Serrano, L.; Crits-Christoph, A.; Pekar, J.E.; Goldstein, S.A.; Rasmussen, A.L.; Kraemer, M.U.G.; Newman, C.; Koopmans, M.P.G.; et al. The Huanan Seafood Wholesale Market in Wuhan was the early epicenter of the COVID-19 pandemic. Science 2022, 377, 951–959.
- The World Health Organization. Coronavirus disease (COVID-19) Pandemic. Available online: https://www.who.int/europe/emergencies/situations/covid-19 (accessed on 12 April 2023).
- National Library of Medicine; National Center for Biology Information. Severe Acute Respiratory Syndrome Coronavirus 2 Isolate Wuhan-Hu-1, Complete Genome. GenBank: MN908947.3. Available online: https://www.ncbi.nlm.nih.gov/nuccore/MN908947 (accessed on 12 April 2023).
- Grellet, E.; L’Hôte, I.; Goulet, A.; Imbert, I. Replication of the coronavirus genome: A paradox among positive-strand RNA viruses. J. Biol. Chem. 2022, 298, 101923.
- Roland, D.; Gardiner, A.; Razzaq, D.; Rose, K.; Bressan, S.; Honeyford, K.; Buonsenso, D.; Da Dalt, L.; De, T.; Farrugia, R.; et al. Influence of epidemics and pandemics on paediatric ED use: A systematic review. Arch. Dis. Child. 2023, 108, 115–122.
- Pyone, T.; Aung, T.T.; Endericks, T.; Myint, N.W.; Inamdar, L.; Collins, S.; Pwint, K.H.; Hein, B.B.; Wilson, A. Health system governance in strengthening International Health Regulations (IHR) compliance in Myanmar. BMJ Glob. Health 2020, 5, e003566.
- Verikios, G. The dynamic effects of infectious disease outbreaks: The case of pandemic influenza and human coronavirus. Socio-Econ. Plan. Sci. 2020, 71, 100898.
- Castelli, V.; Cimini, A.; Ferri, C. Cytokine Storm in COVID-19: “When You Come Out of the Storm, You Won’t Be the Same Person Who Walked in”. Front. Immunol. 2020, 11, 2132.
- Araf, Y.; Faruqui, N.A.; Anwar, S.; Hosen, M.J. SARS-CoV-2: A new dimension to our understanding of coronaviruses. Int. Microbiol. 2021, 24, 19–24.
- Vahabi, M.; Ghazanfari, T.; Sepehrnia, S. Molecular mimicry, hyperactive immune system, and SARS-COV-2 are three prerequisites of the autoimmune disease triangle following COVID-19 infection. Int. Immunopharmacol. 2022, 112, 109183.
- Keshavarzi, A.; Horry, H.R. Bayesian estimation of a dynamic stochastic general equilibrium model with health disaster risk. Stoch. Environ. Res. Risk Assess. 2023, 37, 1199–1211.
- Cascella, M.; Rajnik, M.; Aleem, A.; Dulebohn, S.C.; Di Napoli, R. Features, Evaluation, and Treatment of Coronavirus (COVID-19). 9 January 2023. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://pubmed.ncbi.nlm.nih.gov/32150360 (accessed on 24 May 2023).
- Zebardast, A.; Hasanzadeh, A.; Ebrahimian Shiadeh, S.A.; Tourani, M.; Yahyapour, Y. COVID-19: A trigger of autoimmune diseases. Cell Biol. Int. 2023, 47, 848–858.
- Vilser, D. Long Covid/Post-COVID-19-Syndrom bei Kindern und Jugendlichen. Pädiatrie 2022, 34, 20–25.
- Castanares-Zapatero, D.; Chalon, P.; Kohn, L.; Dauvrin, M.; Detollenaere, J.; Maertens de Noordhout, C.; Primus-de Jong, C.; Cleemput, I.; Van den Heede, K. Pathophysiology and mechanism of long COVID: A comprehensive review. Ann. Med. 2022, 54, 1473–1487.
- de Carvalho, S.S.; Simões e Silva, A.C.; Sabino, A.D.P.; Evangelista, F.C.; Gomes, K.B.; Dusse, L.M.; Rios, D.R. Influence of ACE I/D Polymorphism on Circulating Levels of Plasminogen Activator Inhibitor 1, D-Dimer, Ultrasensitive C-Reactive Protein and Transforming Growth Factor β1 in Patients Undergoing Hemodialysis. PLoS ONE 2016, 11, e0150613.
- Lamers, M.M.; Haagmans, B.L. SARS-CoV-2 pathogenesis. Nat. Rev. Microbiol. 2022, 20, 270–284.
- Silva, M.G.; Falcoff, N.L.; Corradi, G.R.; Di Camillo, N.; Seguel, R.F.; Tabaj, G.C.; Guman, G.R.; de Matteo, E.; Nuñez, M.; Gironacci, M.M. Effect of age on human ACE2 and ACE2-expressing alveolar type II cells levels. Pediatr. Res. 2023, 93, 948–952.
- Matsuyama, S.; Nagata, N.; Shirato, K.; Kawase, M.; Takeda, M.; Taguchi, F. Efficient activation of the severe acute respiratory syndrome coronavirus spike protein by the transmembrane protease TMPRSS2. J. Virol. 2010, 84, 12658–12664.
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8.
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev. 2007, 128, 92–105.
- Hachim, I.Y.; Hachim, M.Y.; Talaat, I.M.; López-Ozuna, V.M.; Saheb Sharif-Askari, N.; Al Heialy, S.; Halwani, R.; Hamid, Q. The Molecular Basis of Gender Variations in Mortality Rates Associated with the Novel Coronavirus (COVID-19) Outbreak. Front. Mol. Biosci. 2021, 8, 728409.
- Guo, X.; Zhu, Y.; Hong, Y. Decreased Mortality of COVID-19 With Renin-Angiotensin-Aldosterone System Inhibitors Therapy in Patients with Hypertension: A Meta-Analysis. Hypertension 2020, 76, e13–e14.
- Papola, F.; Biancofiore, V.; Angeletti, C.; Grimaldi, A.; Carucci, A.C.; Cofini, V.; Necozione, S.; Rosciano, A.; Marinangeli, F.; Cervelli, C. Anti-AT1R autoantibodies and prediction of the severity of Covid-19. Hum. Immunol. 2022, 83, 130–133.
- Bastolla, U.; Chambers, P.; Abia, D.; Garcia-Bermejo, M.-L.; Fresno, M. Is Covid-19 Severity Associated with ACE2 Degradation? Front. Drug. Discov. 2022, 1, 789710.
- Matschke, J.; Lütgehetmann, M.; Hagel, C.; Sperhake, J.P.; Schröder, A.S.; Edler, C.; Mushumba, H.; Fitzek, A.; Allweiss, L.; Dandri, M.; et al. Neuropathology of patients with COVID-19 in Germany: A post-mortem case series. Lancet Neurol. 2020, 19, 919–929.
- Asadi-Pooya, A.A.; Akbari, A.; Emami, A.; Lotfi, M.; Rostamihosseinkhani, M.; Nemati, H.; Barzegar, Z.; Kabiri, M.; Zeraatpisheh, Z.; Farjoud-Kouhanjani, M.; et al. Long COVID syndrome-associated brain fog. J. Med. Virol. 2022, 94, 979–984.
- Stephenson, T.; Shafran, R.; Ladhani, S.N. Long COVID in children and adolescents. Curr. Opin. Infect. Dis. 2022, 35, 461–467.
- Griffin, D.E. Why does viral RNA sometimes persist after recovery from acute infections? PLoS Biol. 2022, 20, e3001687.
- Lopez-Leon, S.; Wegman-Ostrosky, T.; Ayuzo del Valle, N.C.; Perelman, C.; Sepulveda, R.; Rebolledo, P.A.; Cuapio, A.; Villapol, S. Long-COVID in children and adolescents: A systematic review and meta-analyses. Sci. Rep. Sci. 2022, 12, 9950.
- Rong, Z.; Mai, H.; Kapoor, S.; Puelles, G.V.; Czogalla, J.; Schädler, J.; Vering, J.; Delbridge, C.; Steinke, H.; Frenzel, H.; et al. SARS-CoV-2 Spike Protein Accumulation in the Skull-Meninges-Brain Axis: Potential Implications for Long-Term Neurological Complications in post-COVID-19. bioRxiv 2023.
- Fullard, J.F.; Lee, H.C.; Voloudakis, G.; Suo, S.; Javidfar, B.; Shao, Z.; Peter, C.; Zhang, W.; Jiang, S.; Corvelo, A.; et al. Single-nucleus transcriptome analysis of human brain immune response in patients with severe COVID-19. Genome Med. 2021, 13, 118.
- Theoharides, T.C. Could SARS-CoV-2 Spike Protein Be Responsible for Long-COVID Syndrome? Mol. Neurobiol. 2022, 59, 1850–1861.
- Theoharides, T.C.; Kempuraj, D. Role of SARS-CoV-2 Spike-Protein-Induced Activation of Microglia and Mast Cells in the Pathogenesis of Neuro-COVID. Cells 2023, 12, 688.
- de Melo, G.D.; Lazarini, F.; Levallois, S.; Hautefort, C.; Michel, V.; Larrous, F.; Verillaud, B.; Aparicio, C.; Wagner, S.; Gheusi, G.; et al. COVID-19-related anosmia is associated with viral persistence and inflammation in human olfactory epithelium and brain infection in hamsters. Sci. Transl. Med. 2021, 13, eabf8396.
- Jacobs, J.J.L. Persistent SARS-2 infections contribute to long COVID-19. Med. Hypotheses 2021, 149, 110538.
- Casabianca, M.; Caula, C.; Titomanlio, L.; Lenglart, L. Neurological consequences of SARS-CoV-2 infections in the pediatric population. Front. Pediatr. 2023, 11, 1123348.
- Tian, Y.; Rong, L.; Nian, W.; He, Y. Review article: Gastrointestinal features in COVID-19 and the possibility of faecal transmission. Aliment. Pharmacol. Ther. 2020, 51, 843–851.
- Ma, X.; Guan, C.; Chen, R.; Wang, Y.; Feng, S.; Wang, R.; Qu, G.; Zhao, S.; Wang, F.; Wang, X.; et al. Pathological and molecular examinations of postmortem testis biopsies reveal SARS-CoV-2 infection in the testis and spermatogenesis damage in COVID-19 patients. Cell. Mol. Immunol. 2021, 18, 487–489.
- Cherne, M.D.; Gentry, A.B.; Nemudraia, A.; Nemudryi, A.; Hedges, J.F.; Walk, H.; Blackwell, K.; Snyder, D.T.; Jerome, M.; Madden, W.; et al. Severe Acute Respiratory Syndrome Coronavirus 2 Is Detected in the Gastrointestinal Tract of Asymptomatic Endoscopy Patients but Is Unlikely to Pose a Significant Risk to Healthcare Personnel. Gastro Hep. Adv. 2022, 1, 844–852.
- Connelly, Z.M.; Whitaker, D.; Dullea, A.; Ramasamy, R. SARS-CoV-2 Effects on the Male Genitourinary System. Am. J. Clin. Exp. Urol. 2022, 10, 199–209.
- Zollner, A.; Koch, R.; Jukic, A.; Pfister, A.; Meyer, M.; Rössler, A.; Kimpel, J.; Adolph, T.E.; Tilg, H. Postacute COVID-19 is Characterized by Gut Viral Antigen Persistence in Inflammatory Bowel Diseases. Gastroenterology 2022, 163, 495–506.e8.
- Arostegui, D.; Castro, K.; Schwarz, S.; Vaidy, K.; Rabinowitz, S.; Wallach, T.D. Persistent SARS-CoV-2 Nucleocapsid Protein Presence in the Intestinal Epithelium of a Pediatric Patient 3 Months After Acute Infection. JPGN Rep. 2022, 3, e152.
- Tullie, L.; Ford, K.; Bisharat, M.; Watson, T.; Thakkar, H.; Mullassery, D.; Giuliani, S.; Blackburn, S.; Cross, K.; De Coppi, P.; et al. Gastrointestinal features in children with COVID-19: An observation of varied presentation in eight children. Lancet Child Adolesc. Health 2020, 4, e19–e20.
- Colmenero, I.; Santonja, C.; Alonso-Riaño, M.; Noguera-Morel, L.; Hernández-Martín, A.; Andina, D.; Wiesner, T.; Rodríguez-Peralto, J.L.; Requena, L.; Torrelo, A. SARS-CoV-2 endothelial infection causes COVID-19 chilblains: Histopathological, immunohistochemical and ultrastructural study of seven paediatric cases. Br. J. Dermatol. 2020, 183, 729–737.
- Fainardi, V.; Meoli, A.; Chiopris, G.; Motta, M.; Skenderaj, K.; Grandinetti, R.; Bergomi, A.; Antodaro, F.; Zona, S.; Esposito, S. Long COVID in Children and Adolescents. Life 2022, 12, 285.
- Li, W.T.; Zhang, Y.; Liu, M.; Liu, Y.Q.; Ma, X. Prolonged viral shedding in feces of children with COVID-19: A systematic review and synthesis of data. Eur. J. Pediatr. 2022, 181, 4011–4017.
- Santos, V.S.; Gurgel, R.Q.; Cuevas, L.E.; Martins-Filho, P.R. Prolonged Fecal Shedding of SARS-CoV-2 in Pediatric Patients: A Quantitative Evidence Synthesis. J. Pediatr. Gastroenterol. Nutr. 2020, 71, 150–152.
- Craddock, V.; Mahajan, A.; Spikes, L.; Krishnamachary, B.; Ram, A.K.; Kumar, A.; Chen, L.; Chalise, P.; Dhillon, N.K. Persistent circulation of soluble and extracellular vesicle-linked Spike protein in individuals with postacute sequelae of COVID-19. J. Med. Virol. 2023, 95, e28568.
- Dang, X.T.T.; Kavishka, J.M.; Zhang, D.X.; Pirisinu, M.; Le, M.T.N. Extracellular Vesicles as an Efficient and Versatile System for Drug Delivery. Cells 2020, 9, 2191.
- Flores-Alanis, A.; Sandner-Miranda, L.; Delgado, G.; Cravioto, A.; Morales-Espinosa, R. The receptor binding domain of SARS-CoV-2 spike protein is the result of an ancestral recombination between the bat-CoV RaTG13 and the pangolin-CoV MP789. BMC Res. Notes 2020, 13, 398.
- Scott, T.A.; Supramaniam, A.; Idris, A.; Cardoso, A.A.; Shrivastava, S.; Kelly, G.; Grepo, N.A.; Soemardy, C.; Ray, R.M.; McMillan, N.A.J.; et al. Engineered extracellular vesicles directed to the spike protein inhibit SARS-CoV-2. Mol. Ther. Methods Clin. Dev. 2022, 24, 355–366.
- Rotulo, G.A.; Palma, P. Understanding COVID-19 in children: Immune determinants and post-infection conditions. Pediatr. Res. 2023.
- Pou, C.; Nkulikiyimfura, D.; Henckel, E.; Olin, A.; Lakshmikanth, T.; Mikes, J.; Wang, J.; Chen, Y.; Bernhardsson, A.K.; Gustafsson, A.; et al. The repertoire of maternal anti-viral antibodies in human newborns. Nat. Med. 2019, 25, 591–596.
- Koch, C.M.; Prigge, A.D.; Anekalla, K.R.; Shukla, A.; Do Umehara, H.C.; Setar, L.; Chavez, J.; Abdala-Valencia, H.; Politanska, Y.; Markov, N.S.; et al. Age-related Differences in the Nasal Mucosal Immune Response to SARS-CoV-2. Am. J. Respir. Cell Mol. Biol. 2022, 66, 206–222.
- Brodin, P. SARS-CoV-2 infections in children: Understanding diverse outcomes. Immunity 2022, 55, 201–209.
- Varchetta, S.; Mele, D.; Oliviero, B.; Mantovani, S.; Ludovisi, S.; Cerino, A.; Bruno, R.; Castelli, A.; Mosconi, M.; Vecchia, M.; et al. Unique immunological profile in patients with COVID-19. Cell. Mol. Immunol. 2021, 18, 604–612.
- Abarca-Zabalía, J.; González-Jiménez, A.; Calle-Rubio, M.; López-Pastor, A.R.; Fariña, T.; Ramos-Acosta, C.; Anguita, E.; Urcelay, E.; Espino-Paisán, L. Alterations in the immune system persist after one year of convalescence in severe COVID-19 patients. Front. Immunol. 2023, 14, 1127352.
- Rojas, M.; Restrepo-Jiménez, P.; Monsalve, D.M.; Pacheco, Y.; Acosta-Ampudia, Y.; Ramírez-Santana, C.; Leung, P.S.C.; Ansari, A.A.; Gershwin, M.E.; Anaya, J.M. Molecular mimicry and autoimmunity. J. Autoimmun. 2018, 95, 100–123.
- Caso, F.; Costa, L.; Ruscitti, P.; Navarini, L.; Del Puente, A.; Giacomelli, R.; Scarpa, R. Could Sars-coronavirus-2 trigger autoimmune and/or autoinflammatory mechanisms in genetically predisposed subjects? Autoimmun. Rev. 2020, 19, 102524.
- Sundaresan, B.; Shirafkan, F.; Ripperger, K.; Rattay, K. The Role of Viral Infections in the Onset of Autoimmune Diseases. Viruses 2023, 15, 782.
- Di Sante, G.; Buonsenso, D.; De Rose, C.; Valentini, P.; Ria, F.; Sanguinetti, M.; Sali, M. Immune profile of children with post-acute sequelae of SARS-CoV-2 infection (Long COVID). medRxiv 2021.
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babajew, D.; Oliveira, T.Y.; et al. Evolution of antibody immunity to SARS-CoV-2. Nature 2021, 591, 639–644.
- Wang, E.Y.; Mao, T.; Klein, J.; Dai, Y.; Huck, J.D.; Jaycox, J.R.; Liu, F.; Zhou, T.; Israelow, B.; Wong, P.; et al. Diverse functional autoantibodies in patients with COVID-19. Nature 2021, 595, 283–288.
- Sun, J.; Xiao, J.; Sun, R.; Tang, X.; Liang, C.; Lin, H.; Zeng, L.; Hu, J.; Yuan, R.; Zhou, P.; et al. Prolonged Persistence of SARS-CoV-2 RNA in Body Fluids. Emerg. Infect. Dis. 2020, 26, 1834–1838.
- Zhou, M.; Yin, Z.; Xu, J.; Wang, S.; Liao, T.; Wang, K.; Li, Y.; Yang, F.; Wang, Z.; Yang, G.; et al. Inflammatory Profiles and Clinical Features of Coronavirus 2019 Survivors 3 Months After Discharge in Wuhan, China. J. Infect. Dis. 2021, 224, 1473–1488.
- Patterson, B.K.; Francisco, E.B.; Yogendra, R.; Long, E.; Pise, A.; Rodrigues, H.; Hall, E.; Herrera, M.; Parikh, P.; Guevara-Coto, J.; et al. Persistence of SARS CoV-2 S1 Protein in CD16+ Monocytes in Post-Acute Sequelae of COVID-19 (PASC) up to 15 Months Post-Infection. Front. Immunol. 2022, 12, 746021.
- Matsumiya, T.; Ota, K.; Imaizumi, T.; Yoshida, H.; Kimura, H.; Satoh, K. Characterization of synergistic induction of CX3CL1/fractalkine by TNF-alpha and IFN-gamma in vascular endothelial cells: An essential role for TNF-alpha in post-transcriptional regulation of CX3CL1. J. Immunol. 2010, 184, 4205–4214.
- Guilliams, M.; Mildner, A.; Yona, S. Developmental and Functional Heterogeneity of Monocytes. Immunity 2018, 49, 595–613.
- Kapellos, T.S.; Bonaguro, L.; Gemünd, I.; Reusch, N.; Saglam, A.; Hinkley, E.R.; Schultze, J.L. Human Monocyte Subsets and Phenotypes in Major Chronic Inflammatory Diseases. Front. Immunol. 2019, 10, 2035.
- Park, J.; Dean, L.S.; Jiyarom, B.; Gangcuangco, L.M.; Shah, P.; Awamura, T.; Ching, L.L.; Nerurkar, V.R.; Chow, D.C.; Igno, F.; et al. Elevated circulating monocytes and monocyte activation in COVID-19 convalescent individuals. Front. Immunol. 2023, 14, 1151780.
- Dowd, J.B.; Palermo, T.; Brite, J.; McDade, T.W.; Aiello, A. Seroprevalence of Epstein-Barr virus infection in U.S. children ages 6–19, 2003–2010. PLoS ONE 2013, 8, e64921.
- Neves, M.; Marinho-Dias, J.; Ribeiro, J.; Sousa, H. Epstein-Barr virus strains and variations: Geographic or disease-specific variants? J. Med. Virol. 2017, 89, 373–387.
- Gold, J.E.; Okyay, R.A.; Licht, W.E.; Hurley, D.J. Investigation of Long COVID Prevalence and Its Relationship to Epstein-Barr Virus Reactivation. Pathogens 2021, 10, 763.
- Houen, G.; Trier, N.H. Epstein-Barr Virus and Systemic Autoimmune Diseases. Front. Immunol. 2021, 11, 587380.
- Pallanti, S.; Di Ponzio, M. PANDAS/PANS in the COVID-19 Age: Autoimmunity and Epstein–Barr Virus Reactivation as Trigger Agents? Children 2023, 10, 648.
- Wiersinga, W.J.; Rhodes, A.; Cheng, A.C.; Peacock, S.J.; Prescott, H.C. Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19): A Review. JAMA 2020, 324, 782–793.
- Cipollaro, L.; Giordano, L.; Padulo, J.; Oliva, F.; Maffulli, N. Musculoskeletal symptoms in SARS-CoV-2 (COVID-19) patients. J. Orthop. Surg. Res. 2020, 15, 178.
- Beydon, M.; Chevalier, K.; Al Tabaa, O.; Hamroun, S.; Delettre, A.S.; Thomas, M.; Herrou, J.; Riviere, E.; Mariette, X. Myositis as a manifestation of SARS-CoV-2. Ann. Rheum. Dis. 2021, 80, e42.
- Manzano, G.S.; Woods, J.K.; Amato, A.A. Covid-19-Associated Myopathy Caused by Type I Interferonopathy. N. Engl. J. Med. 2020, 383, 2389–2390.
- Movahedi, N.; Ziaee, V. COVID-19 and myositis; true dermatomyositis or prolonged post viral myositis? Pediatr. Rheumatol. 2021, 19, 86.
- Tanboon, J.; Nishino, I. COVID-19-associated myositis may be dermatomyositis. Muscle Nerve 2021, 63, E9–E10.
- Qian, J.; Xu, H. COVID-19 Disease and Dermatomyositis: A Mini-Review. Front. Immunol. 2022, 12, 747116.
- Gokhale, Y.; Patankar, A.; Holla, U.; Shilke, M.; Kalekar, L.; Karnik, N.D.; Bidichandani, K.; Baveja, S.; Joshi, A. Dermatomyositis during COVID-19 Pandemic (A Case Series): Is there a Cause Effect Relationship? J. Assoc. Physicians India 2020, 68, 20–24.
- Crivelenti, L.R.M.P.; Frazão, M.M.N.; Maia, M.P.M.; Gomes, F.H.R.; de Carvalho, L.M. Chronic arthritis related to SARS-CoV-2 infection in a pediatric patient: A case report. Braz. J. Infect. Dis. 2021, 25, 101585.
- Sinaei, R.; Pezeshki, S.; Parvaresh, S.; Sinaei, R.; Shiari, R.; Hassas Yeganeh, M.; Bazargn, N.; Gharaei, N. Post SARS-CoV-2 infection reactive arthritis: A brief report of two pediatric cases. Pediatr. Rheumatol. Online J. 2021, 19, 89.
- Ailioaie, L.M.; Ailioaie, C.; Litscher, G. Implications of SARS-CoV-2 Infection in Systemic Juvenile Idiopathic Arthritis. Int. J. Mol. Sci. 2022, 23, 4268.
- Zitouni, J.; Bursztejn, A.C.; Belloni Fortina, A.; Beauchet, A.; Di Lernia, V.; Lesiak, A.; Thomas, J.; Topkarci, Z.; Murashkin, N.; Brzezinski, P.; et al. Children with psoriasis and COVID-19: Factors associated with an unfavourable COVID-19 course, and the impact of infection on disease progression (Chi-PsoCov registry). J. Eur. Acad. Dermatol. Venereol. 2022, 36, 2076–2086.
- Alexander, A.J.; Joshi, A.; Mehendale, A. The Musculoskeletal Manifestations of COVID-19: A Narrative Review Article. Cureus 2022, 14, e29076.
- Pal, A.; Roongta, R.; Mondal, S.; Sinha, D.; Sinhamahapatra, P.; Ghosh, A.; Chattopadhyay, A. Does post-COVID reactive arthritis exist? Experience of a tertiary care centre with a review of the literature. Reumatol. Clin. 2023, 19, 67–73.
- Kumar, P.; Jat, K.R. Post-COVID-19 Sequelae in Children. Indian J. Pediatr. 2023, 90, 605–611.
- Ailioaie, L.M.; Ailioaie, C.; Litscher, G.; Chiran, D.A. Celiac Disease and Targeting the Molecular Mechanisms of Autoimmunity in COVID Pandemic. Int. J. Mol. Sci. 2022, 23, 7719.
- Chen, B.; Julg, B.; Mohandas, S.; Bradfute, S.B. RECOVER Mechanistic Pathways Task Force (2023). Viral persistence, reactivation, and mechanisms of long COVID. Elife 2023, 12, e86015.
- de Oliveira, G.L.V.; Oliveira, C.N.S.; Pinzan, C.F.; de Salis, L.V.V.; Cardoso, C.R.B. Microbiota Modulation of the Gut-Lung Axis in COVID-19. Front. Immunol. 2021, 12, 635471.
- Troisi, J.; Venutolo, G.; Pujolassos Tanyà, M.; Delli Carri, M.; Landolfi, A.; Fasano, A. COVID-19 and the gastrointestinal tract: Source of infection or merely a target of the inflammatory process following SARS-CoV-2 infection? World J. Gastroenterol. 2021, 27, 1406–1418.
- Settanni, C.R.; Ianiro, G.; Ponziani, F.R.; Bibbò, S.; Segal, J.P.; Cammarota, G.; Gasbarrini, A. COVID-19 as a trigger of irritable bowel syndrome: A review of potential mechanisms. World J. Gastroenterol. 2021, 27, 7433–7445.
- Dhar, D.; Mohanty, A. Gut microbiota and Covid-19- possible link and implications. Virus Res. 2020, 285, 198018.
- Azer, S.A. COVID-19: Pathophysiology, diagnosis, complications and investigational therapeutics. New Microbes New Infect. 2020, 37, 100738.
- Chakaroun, R.M.; Massier, L.; Kovacs, P. Gut Microbiome, Intestinal Permeability, and Tissue Bacteria in Metabolic Disease: Perpetrators or Bystanders? Nutrients 2020, 12, 1082.
- Tsounis, E.P.; Triantos, C.; Konstantakis, C.; Marangos, M.; Assimakopoulos, S.F. Intestinal barrier dysfunction as a key driver of severe COVID-19. World J. Virol. 2023, 12, 68–90.
- Markov, P.V.; Ghafari, M.; Beer, M.; Lythgoe, K.; Simmonds, P.; Stilianakis, N.I.; Katzourakis, A. The evolution of SARS-CoV-2. Nat. Rev. Microbiol. 2023, 21, 361–379.
- Kwak, B.O.; Eun, B.W. COVID-19 in immunocompromised children and adolescents. Clin. Exp. Pediatr. 2023, 66, 182–189.
- Acosta-Ampudia, Y.; Monsalve, D.M.; Rojas, M.; Rodríguez, Y.; Zapata, E.; Ramírez-Santana, C.; Anaya, J.M. Persistent Autoimmune Activation and Proinflammatory State in Post-Coronavirus Disease 2019 Syndrome. J. Infect. Dis. 2022, 225, 2155–2162.
- Queiroz, M.A.F.; Neves, P.F.M.D.; Lima, S.S.; Lopes, J.D.C.; Torres, M.K.D.S.; Vallinoto, I.M.V.C.; Bichara, C.D.A.; Dos Santos, E.F.; de Brito, M.T.F.M.; da Silva, A.L.S.; et al. Cytokine Profiles Associated with Acute COVID-19 and Long COVID-19 Syndrome. Front. Cell. Infect. Microbiol. 2022, 12, 922422.
- Yin, J.X.; Agbana, Y.L.; Sun, Z.S.; Fei, S.W.; Zhao, H.Q.; Zhou, X.N.; Chen, J.H.; Kassegne, K. Increased interleukin-6 is associated with long COVID-19: A systematic review and meta-analysis. Infect. Dis. Poverty 2023, 12, 43.
- Al-Sadi, R.; Dharmaprakash, V.; Nighot, P.; Guo, S.; Nighot, M.; Do, T.; Ma, T.Y. Bifidobacterium bifidum Enhances the Intestinal Epithelial Tight Junction Barrier and Protects against Intestinal Inflammation by Targeting the Toll-like Receptor-2 Pathway in an NF-κB-Independent Manner. Int. J. Mol. Sci. 2021, 22, 8070.
- Ailioaie, L.M.; Litscher, G. Probiotics, Photobiomodulation, and Disease Management: Controversies and Challenges. Int. J. Mol. Sci. 2021, 22, 4942.
- Bacorn, M.; Romero-Soto, H.N.; Levy, S.; Chen, Q.; Hourigan, S.K. The Gut Microbiome of Children during the COVID-19 Pandemic. Microorganisms 2022, 10, 2460.
- Yonker, L.M.; Gilboa, T.; Ogata, A.F.; Senussi, Y.; Lazarovits, R.; Boribong, B.P.; Bartsch, Y.C.; Loiselle, M.; Rivas, M.N.; Porritt, R.A.; et al. Multisystem inflammatory syndrome in children is driven by zonulin-dependent loss of gut mucosal barrier. J. Clin. Investig. 2021, 131, e149633.
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S.; et al. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut 2021, 70, 698–706.
- Haran, J.P.; Bradley, E.; Zeamer, A.L.; Cincotta, L.; Salive, M.C.; Dutta, P.; Mutaawe, S.; Anya, O.; Meza-Segura, M.; Moormann, A.M.; et al. Inflammation-type dysbiosis of the oral microbiome associates with the duration of COVID-19 symptoms and long COVID. JCI Insight 2021, 6, e152346.
- Ancona, G.; Alagna, L.; Alteri, C.; Palomba, E.; Tonizzo, A.; Pastena, A.; Muscatello, A.; Gori, A.; Bandera, A. Gut and airway microbiota dysbiosis and their role in COVID-19 and long-COVID. Front. Immunol. 2023, 14, 1080043.
- Gareau, M.G.; Barrett, K.E. Role of the microbiota-gut-brain axis in postacute COVID syndrome. Am. J. Physiol. Gastrointest. Liver Physiol. 2023, 324, G322–G328.
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013.
- Bostick, J.W.; Schonhoff, A.M.; Mazmanian, S.K. Gut microbiome-mediated regulation of neuroinflammation. Curr. Opin. Immunol. 2022, 76, 102177.
- Ailioaie, L.M.; Ailioaie, C.; Litscher, G. Photobiomodulation in Alzheimer’s Disease—A Complementary Method to State-of-the-Art Pharmaceutical Formulations and Nanomedicine? Pharmaceutics 2023, 15, 916.
- Lorens, S.; Nava, E.; Muñoz-López, M.; Sánchez-Larsen, Á.; Segura, T. Neurological Symptoms of COVID-19: The Zonulin Hypothesis. Front. Immunol. 2021, 12, 665300.
- Okuyucu, M.; Yalcin Kehribar, D.; Çapraz, M.; Çapraz, A.; Arslan, M.; Çelik, Z.B.; Usta, B.; Birinci, A.; Ozgen, M. The Relationship Between COVID-19 Disease Severity and Zonulin Levels. Cureus 2022, 14, e28255.
- Al Bataineh, M.T.; Henschel, A.; Mousa, M.; Daou, M.; Waasia, F.; Kannout, H.; Khalili, M.; Kayasseh, M.A.; Alkhajeh, A.; Uddin, M.; et al. Gut Microbiota Interplay With COVID-19 Reveals Links to Host Lipid Metabolism Among Middle Eastern Populations. Front. Microbiol. 2021, 12, 761067.
- Reinold, J.; Farahpour, F.; Fehring, C.; Dolff, S.; Konik, M.; Korth, J.; van Baal, L.; Hoffmann, D.; Buer, J.; Witzke, O.; et al. A Pro-Inflammatory Gut Microbiome Characterizes SARS-CoV-2 Infected Patients and a Reduction in the Connectivity of an Anti-Inflammatory Bacterial Network Associates with Severe COVID-19. Front. Cell. Infect. Microbiol. 2021, 11, 747816.
- Cheng, X.; Zhang, Y.; Li, Y.; Wu, Q.; Wu, J.; Park, S.K.; Guo, C.; Lu, J. Meta-analysis of 16S rRNA microbial data identified alterations of the gut microbiota in COVID-19 patients during the acute and recovery phases. BMC Microbiol. 2022, 22, 274.
- Guido, C.A.; Lucidi, F.; Midulla, F.; Zicari, A.M.; Bove, E.; Avenoso, F.; Amedeo, I.; Mancino, E.; Nenna, R.; De Castro, G.; et al. Long-Covid Group of Department of Maternal Sciences. Neurological and psychological effects of long COVID in a young population: A cross-sectional study. Front. Neurol. 2022, 13, 925144.
- Schneider, K.M.; Blank, N.; Alvarez, Y.; Thum, K.; Lundgren, P.; Litichevskiy, L.; Sleeman, M.; Bahnsen, K.; Kim, J.; Kardo, S.; et al. The enteric nervous system relays psychological stress to intestinal inflammation. Cell 2023, 186, 2823–2838.e20.
- Sidik, S. Chronic stress can inflame the gut-now scientists know why. Nature 2023, 618, 221–222.
More
Information
Subjects:
Pediatrics
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
919
Entry Collection:
COVID-19
Revisions:
2 times
(View History)
Update Date:
06 Jul 2023
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No