Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Elisa Pinto | -- | 2420 | 2023-06-22 09:19:59 | | | |
| 2 | Rita Xu | -16 word(s) | 2404 | 2023-06-26 03:48:47 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Pinto, E.; Pelizzaro, F.; Farinati, F.; Russo, F.P. Angiogenesis in Hepatocellular Carcinoma. Encyclopedia. Available online: https://encyclopedia.pub/entry/45963 (accessed on 25 July 2026).
Pinto E, Pelizzaro F, Farinati F, Russo FP. Angiogenesis in Hepatocellular Carcinoma. Encyclopedia. Available at: https://encyclopedia.pub/entry/45963. Accessed July 25, 2026.
Pinto, Elisa, Filippo Pelizzaro, Fabio Farinati, Francesco Paolo Russo. "Angiogenesis in Hepatocellular Carcinoma" Encyclopedia, https://encyclopedia.pub/entry/45963 (accessed July 25, 2026).
Pinto, E., Pelizzaro, F., Farinati, F., & Russo, F.P. (2023, June 22). Angiogenesis in Hepatocellular Carcinoma. In Encyclopedia. https://encyclopedia.pub/entry/45963
Pinto, Elisa, et al. "Angiogenesis in Hepatocellular Carcinoma." Encyclopedia. Web. 22 June, 2023.
Copy Citation
Hepatocellular carcinoma (HCC) is the most common primary liver malignancy. The hypervascular nature of the majority of HCCs and the peculiar vascular derangement occurring during liver carcinogenesis underscore the importance of angiogenesis in the development and progression of these tumors. Indeed, several angiogenic molecular pathways have been identified as deregulated in HCC. The hypervascular nature and the peculiar vascularization of HCC, as well as deregulated angiogenic pathways, represent major therapeutic targets.
hepatocellular carcinoma
angiogenesis
tyrosine kinase inhibitors
1. Introduction
Hepatocellular carcinoma (HCC) represents the sixth most commonly diagnosed cancer and the third leading cause of cancer-related death globally [1]. Approximately half of HCC patients are diagnosed at advanced tumor stages, precluding potentially curative treatments such as surgical resection or liver transplantation [2]. As a consequence, the prognosis of patients with HCC is very poor with 5-year survival of 20% [3].
Angiogenesis, one of the fundamental hallmarks of cancer [4], plays a pivotal role in the development and progression of HCC, which is typically a hypervascular tumor [5][6]. Since, the growth of liver tumor requires the formation of new blood vessels, HCC displays intense neoangiogenic activity during its development. Moreover, a peculiar vascular derangement occurs during liver carcinogenesis, since the tumor tends to be almost entirely fed by arterial inflow, unlike the surrounding parenchyma that receives the majority of blood supply through the portal system [7]. However, in liver tumors, newly formed blood vessels display marked vascular abnormalities, which may further activate angiogenic pathways, leading to a vicious cycle. It has been demonstrated that the overactivation of angiogenesis in HCC is associated with worse prognosis. A transcriptomic signature of five genes involved in the angiogenetic process (ANGPT2, NETO2, ESM1, NR4A1, and DLL4) was found to accurately identify rapidly growing tumors and was associated with shorter survival [8]. In addition, several studies suggest that overexpression of vascular endothelial growth factor (VEGF) and its transcription factor hypoxia-inducible factor (HIF)-1α, the two key mediators of angiogenesis, is a negative prognostic factor, particularly in patients treated with surgery and systemic therapies [9][10][11][12][13][14][15][16][17][18][19][20].
The very important role of angiogenesis in the development and progression of HCC provides a strong rationale for antiangiogenic strategies as therapy. Angiogenesis has always been considered an important therapeutic target in these patients. Intra-arterial locoregional treatments (IATs) (i.e., transarterial embolization (TAE) and transarterial chemoembolization (TACE)) are commonly applied treatments for HCC worldwide [21]. Their activity is completely or in part reliant on the embolization of tumor feeding arteries with the aim of achieving tumor ischemic necrosis. Considering systemic therapies, over the last decades, multiple antiangiogenic therapies have been developed. In fact, most currently approved treatments for advanced HCC in the first- and second-line settings target angiogenic pathways [22]. More recently, the combination of an immune checkpoint inhibitor (ICI) anti-programmed death ligand 1 (PD-L1) (atezolizumab) and a monoclonal antibody targeting VEGF (bevacizumab) demonstrated a clear survival benefit over sorafenib [23]. Considering that previous trials with anti-PD1 immune checkpoint inhibitors alone (nivolumab and pembrolizumab) failed to show efficacy in first- and second-line treatment [24][25], these results seem to further confirm the importance of angiogenic pathways in the progression of HCC.
2. Angiogenesis in Hepatocellular Carcinoma
Normal liver receives approximately 26% of cardiac output (~1.1 mL O2/g/min) and consumes approximately 20% of the total O2 used by the body at rest (~0.06 mL O2/g/min) [26]. About 75% of the hepatic blood supply is received by the portal vein, while the rest is received by the hepatic artery. Lobules are the fundamental unit of normal liver: they are segregated by interlobular connective tissue and contain “cords” of hepatic parenchymal cells (hepatocytes), separated by vascular sinusoids. Sinusoidal endothelium is fenestrated and lacks a basement membrane, therefore permitting blood plasma to surround hepatocytes through the space of Disse. Other cells involved in liver physiology are hepatic stellate cells, also known as pericytes, and Kupffer cells, which are resident liver macrophages. Stellate cells are closely linked to sinusoids in the space of Disse and play a crucial role in liver fibrosis after liver damage. In the hepatic sinusoids, arterial (from the hepatic artery) and venous (from the portal vein) blood mix together, and after being “filtered” by hepatocytes, this blood flows out of the lobule through the central hepatic vein.
In liver tumors, newly formed blood vessels display marked vascular abnormalities [27][28], leading to hypovascular areas and severe hypoxia and/or necrosis and causing further stimulation of angiogenesis. Although HCC is a highly angiogenic cancer, it seems to be characterized by hypoxia [5], which has been associated with HCC growth, progression and resistance to therapies [29]. Nevertheless, while some characteristics of HCC (hypervascularity, areas of necrosis and primary resistance to therapy) suggest the presence of severe hypoxia, direct evidence of hypoxia in human HCC is missing [26]. In fact, pO2 in human HCC have not yet been accurately measured directly, and thus, the relevance of hypoxia in determining hypervascularity and arterialization of HCC is still unproved [26]. Moreover, it is important to keep in mind that the activation of angiogenic pathways (HIF target genes) can be achieved by various hypoxia-independent mechanisms [30].
The destabilization of the microvasculature, leading to vascular hyperpermeability, remodeling of the extracellular matrix and endothelial cell activation, is a fundamental step for the initiation of angiogenesis [5]. Activated endothelial cells form new blood vessels by proliferating, migrating and undergoing cord formation. Subsequently, recruited and activated pericytes stabilize the newly formed blood vessels [31][32][33]. During physiological angiogenesis, the release of antiangiogenic molecules balances the expression of proangiogenic factors [34]. By contrast, as shown in Figure 1, tumor-induced angiogenesis results from an imbalance between proangiogenic factors (VEGF-A, -B, -C and -D, angiopoietins, fibroblast growth factor (FGF), hepatocyte growth factor, endoglin (CD105), platelet-derived growth factor (PDGF), and others) and anti-angiogenic molecules (angiostatin, thrombospondin-1, endostatin, and others) [22].
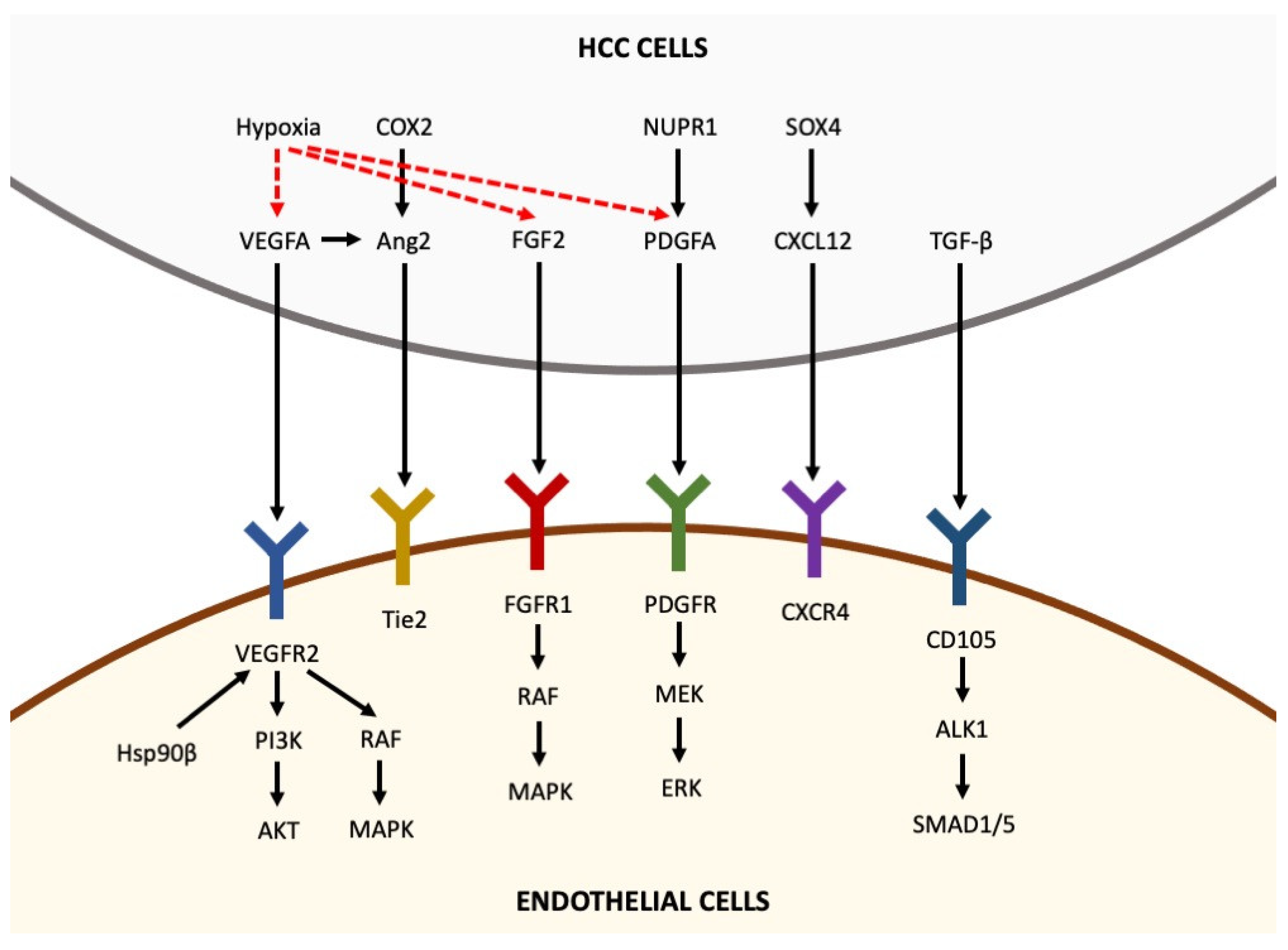
Figure 1. Pro-angiogenic factors inducing angiogenesis in hepatocellular carcinoma. Pro-angiogenic factors, including VEGFA, Ang2, FGF2, PDGFA, CXCL12 and TGF-β, are secreted by HCC cells and bind to their receptors expressed in endothelial cells, thus activating intracellular pathways that promote angiogenesis. Hypoxia is able to upregulate the expression of VEGFA, FGF2 and PDGFA in HCC cells (the arrows indicate the sequentiality of molecular pathways).
2.1. Hypoxia-Inducible Factor 1 (HIF-1)
HIF-1 is a heterodimer composed of two subunits, HIF-1α and HIF-1β. The former is an oxygen-sensitive subunit whose expression is induced under hypoxic conditions, while HIF-1β is constitutively expressed [35]. Regardless of O2 levels, HIF-1α is constitutively transcribed and synthesized through a series of events involving different growth factors and signal molecules. Under normoxic conditions, HIF-1α undergoes rapid degradation by proteasome and ubiquitination within a pathway involving the Von Hippel Lindau protein (pVHL), a tumor suppressor protein that is part of ubiquitin-ligase E3, which recognizes HIF-1α following its prolyl-hydroxylation by proteins containing protohydroxylases (PHD1, PHD2 and PHD3). In addition, asparagine hydroxylation of HIF1α blocks its interaction with the transcriptional co-activators, CREB-binding protein (CBP) and p300. By contrast, under hypoxic conditions, several pathways control the stability and transcriptional activity of this subunit through post-transcriptional modifications, including hydroxylation, acetylation, ubiquitination and phosphorylation reactions [36][37]. Being oxygen-requiring processes, neither hydroxylation nor acetylation of proline and lysine residues can occur under hypoxic conditions, resulting in greater stability of HIF-1α. There is also a pVHL-independent negative regulatory system that acts at the level of transactivation. In addition, the synthesis, degradation, and activity of HIF-1α are also regulated by an O2-independent system involving several cytokines and other signaling molecules [30]. Examples of this regulatory mechanism are the pathway including PI3K-AKT-mTOR, RAS/RAF/MEK/ERK kinase cascade, with phosphorylation of the CBP/p300 coactivator resulting in increased formation of the HIF-1α/p300 complex, the Hsp90 pathway and the Mdm2-p53 system, which is often altered in certain types of hypoxic tumors where low p53 levels are found, resulting in increased levels of HIF-1α [38][39].
HIF-1 acts as a transcription factor binding to 5′(A/G) CGTG- 3′ consensus sequences called hypoxia-responsive elements (HREs), which allows the activation of target genes [36] involved in the processes of tumor metastasis, angiogenesis, metabolic energy, cell differentiation and apoptosis [40][41]. Tumor cells often experience hypoxia due to a decrease in oxygen transport and diffusion and to an increase in O2 consumption. In fact, the intense proliferation of tumor cells causes an increased oxygen demand. Moreover, the distance between cells and the existing vascular system increases, making oxygen diffusion difficult and thus creating hypoxia. Hypoxia and a general imbalance in the distribution of oxygen within a solid tumor are typical features of neoplastic tissue and lead to more aggressive growth of the neoplasm. Oxygen deficiency leads to an increased expression of HIF-1α, which then activates angiogenesis, glucose metabolism, cell proliferation, invasion, and metastasis. Considering angiogenesis, the main HIF-1α target genes include VEGF, angiopoietin 1 and 2 and metalloproteases, leading to the formation of new, albeit unstructured, vessels within the tumor [42].
Beyond its roles in angiogenesis, HIF-1α also has several different roles in cancer progression. It triggers tumor cells invasion, acting in the epithelial–mesenchymal transition (EMT) process, increases cells proliferation and decreases apoptosis [42]. Moreover, HIF-1α is an important regulator of tumor cell metabolism, acting mainly on glucose catabolism, and favoring the development of an acidosic environment, stimulating fatty acid synthesis and glycogen synthesis (Warburg effect) [43].
The role of HIF-1α in HCC is certainly multifaceted as it is involved in several processes that influence carcinogenesis, such as vascularization, inflammation, infection by hepatotropic viruses, and changes in the microenvironment. The importance of HIF in these mechanisms has certainly stimulated research into the role that this marker might have in the treatment of liver cancer. Several in vivo studies highlight the importance of targeting the HIF pathway to inhibit tumor progression and to improve the efficacy of anti-VEGF therapies by controlling the hypoxic tumor microenvironment [44][45]. Indeed, long-term success of sorafenib treatment in HCC is limited due to the development of resistance caused by various mechanisms, including antiangiogenic effects and HIF-mediated cellular responses. Overexpression of HIF-1α and HIF-2α in HCC patients indicates a poor prognosis, prompting exploration of combined therapies targeting HIFs to overcome sorafenib resistance. Targeting both HIF-1α and HIF-2α shows potential as a more effective strategy than selective therapies as there is a strong correlation between the hypoxic microenvironment and sorafenib resistance [44]. Targeting HIF may limit the side effects caused by hypoxia induced by radiation or anti-angiogenic factor therapies, leading to clinically significant treatment improvements [46].
2.2. VEGF/VEGFR
The most well-known regulators of angiogenesis are the VEGF and VEGF receptors (VEGFRs) [47], which are fundamental for HCC development and progression. The ligands VEGF-A, VEGF-B, VEGF-D and VEGF-E belong to a family of structurally related dimeric proteins [48]. These growth factors (VEGF-A, VEGF-C, or VEGF-D) bind and stimulate VEGFR-2, which is expressed in nearly all endothelial cells [48]. VEGF-A is the most important isoform, responsible for angiogenesis and vascular remodeling. The ligand–receptor binding triggers downstream cellular pathways, involving many signal molecules (Y1213, Y1333, Sck, PLC-γ, VRAPAKT, FAK, p38 MAPK, eNOS, Src and PI3K), ultimately leading to formation of new tumor blood vessels within tumors which are essential for facilitating tumor development and progression [49][50]. VEGF expression, with its transcription regulated by the binding between HIF and hypoxia-responsive elements (HREs) [51], is modulated by tissue oxygen levels [52][53].
Increased levels of circulating VEGF have been observed in HCC [54] and have been demonstrated to be associated with accelerated disease progression and poorer prognosis [55][56]. In addition, VEGF seems to play a role in chemoresistance by acting on autophagy through NRP2 and mTOR [48]. These observations [55][56] provide support for the assessment of VEGF-pathway-directed therapies as a useful approach for treating HCC. Moreover, the groundbreaking survival results obtained with the combination atezolizumab + bevacizumab [23] confirm the importance of angiogenesis and support targeting the VEGF axis in HCC.
2.3. PDGF/PDGFR
The PDGF family consists of several ligands (PDGF-A, PDGF-B, PDGF-C, PDGF-D and PDGF-AB [57]) which bind to the tyrosine kinase PDGF receptor (PDGFR)-α and -β expressed on mesenchymal cells (fibroblast, smooth muscle cells, and pericytes). This interaction activates similar pathways to those stimulated by VEGF [57][58]. Binding of PDGF with their corresponding receptors leads to the activation of a signaling cascade which results in upregulation of VEGF and recruitment of perivascular cells. The relevance of PDGF/PDGFR pathways in human HCC is demonstrated by the fact that overexpression of PDGFR-α is associated with vessel density and worse prognosis [22]. Moreover, a shorter survival was demonstrated in HCC patients expressing PDGFR-α, PDGFR-β and VEGF [22]. Nevertheless, the inhibition of the PDGFR pathway as a target for anti-angiogenic therapy in HCC remains of uncertain clinical relevance. Although TKIs (sorafenib and others) also target PDGFR, these drugs also inhibit other pathways, so it is currently unclear what impact the inhibition of the PDGF pathway has on the overall clinical benefit.
2.4. FGF/FGFR
The FGF family includes several ligands that interact with four tyrosine kinase receptors (FGFR-1, -2, -3 and -4) [59]. FGFs and FGFRs are ubiquitously expressed, and among their various functions, they regulate cell growth and maintain VEGF-induced neovascularization [60]. During the initial phases of tumor growth, the cross-talk between FGF-2 and VEGF-A is able to induce neovascularization and boost tumor progression [61]. FGFs and VEGF-A are linked to enhanced capillarization of sinusoids [62], while FGF-induced integrin expression interacts with endothelial cells in the microenvironment, thereby modifying the essential cellular parameters required for angiogenesis. The resistance of advanced HCC to the VEGFR inhibitor sorafenib may be partially explained by this synergism between the FGF and VEGF pathways [63][64].
2.5. Angiopoietin/Tie Pathway
Angiopoietin 1 (Ang1) and 2 (Ang2) are ligands for the Tie2 receptor expressed on endothelial cells [65]. While vascular support cells widely express Ang1, Ang2 is only present at sites of vascular remodeling [66]. Indeed, Ang1 and Ang2 compete for their binding to Tie2, and these interactions modulate the pathway. Ang1 stabilizes the blood vessels, while Ang2 expression in areas of vascular remodeling competes with Ang1 for the interaction with Tie2, destabilizing blood vessel support cells. This is a necessary step to facilitate vessel proliferation induced by VEGF [66].
Patients with HCC showed high levels of Ang2, suggesting a central role in carcinogenesis, potentially together with VEGF [65]. Considering the importance of this pathway in the progression of HCC, some agents targeting Angiopoietins/Tie2 interaction alone or in combination with sorafenib have been tested in clinical practice [67], but any potential clinical benefit remains to be determined.
2.6. Endoglin (CD105)
Endoglin (CD105) expression is increased in actively dividing endothelial cells, including those found in liver cancers [68][69]. It functions as an accessory coreceptor of transforming growth factor-β (TGF-β), antagonizing its inhibitory effects, but it also modulates the transition of endothelial progenitor cells to mature epithelial cells [70].
Endoglin expression is associated with the HCC stage differentiation and aggressiveness, promoting invasion and metastatic spread by increasing VEGF expression [71]. Although intriguing, targeting this pathway for the treatment of HCC has an unclear clinical relevance.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: Globocan Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249.
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Prim. 2021, 7, 6.
- Pelizzaro, F.; Vitale, A.; Sartori, A.; Vieno, A.; Penzo, B.; Russo, F.P.; Frigo, A.C.; Giannini, E.G.; Piccinnu, M.; Rapaccini, G.L.; et al. Surveillance as Determinant of Long-Term Survival in Non-Transplanted Hepatocellular Carcinoma Patients. Cancers 2021, 13, 897.
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70.
- Zhu, A.X.; Duda, D.G.; Sahani, D.V.; Jain, R.K. HCC and angiogenesis: Possible targets and future directions. Nat. Rev. Clin. Oncol. 2011, 8, 292–301.
- Muto, J.; Shirabe, K.; Sugimachi, K.; Maehara, Y. Review of angiogenesis in hepatocellular carcinoma. Hepatol. Res. 2015, 45, 12310.
- Matsui, O.; Kobayashi, S.; Sanada, J.; Kouda, W.; Ryu, Y.; Kozaka, K.; Kitao, A.; Nakamura, K.; Gabata, T. Hepatocelluar nodules in liver cirrhosis: Hemodynamic evaluation (angiography-assisted CT) with special reference to multi-step hepatocarcinogenesis. Abdom. Imaging 2011, 36, 264–272.
- Villa, E.; Critelli, R.; Lei, B.; Marzocchi, G.; Cammà, C.; Giannelli, G.; Pontisso, P.; Cabibbo, G.; Enea, M.; Colopi, S.; et al. Neoangiogenesis-related genes are hallmarks of fast-growing hepatocellular carcinomas and worst survival. Results from a prospective study. Gut 2016, 65, 861–869.
- Liu, L.; Zhu, X.-D.; Wang, W.-Q.; Shen, Y.; Qin, Y.; Ren, Z.-G.; Sun, H.-C.; Tang, Z.-Y. Activation of β-Catenin by Hypoxia in Hepatocellular Carcinoma Contributes to Enhanced Metastatic Potential and Poor Prognosis. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 2740–2750.
- Huang, G.-W.; Yang, L.-Y.; Lu, W.-Q. Expression of hypoxia-inducible factor 1α and vascular endothelial growth factor in hepatocellular carcinoma: Impact on neovascularization and survival. World J. Gastroenterol. 2005, 11, 1705–1708.
- Zhan, P.; Qian, Q.; Yu, L.-K. Serum VEGF level is associated with the outcome of patients with hepatocellular carcinoma: A meta-analysis. HepatoBiliary Surg. Nutr. 2013, 2, 209–215.
- Llovet, J.M.; Peña, C.E.; Lathia, C.D.; Shan, M.; Meinhardt, G.; Bruix, J. Plasma Biomarkers as Predictors of Outcome in Patients with Advanced Hepatocellular Carcinoma. Clin. Cancer Res. 2012, 18, 2290–2300.
- Xia, L.; Mo, P.; Huang, W.; Zhang, L.; Wang, Y.; Zhu, H.; Tian, D.; Liu, J.; Chen, Z.; Zhang, Y.; et al. The TNF-α/ROS/HIF-1-induced Upregulation of FoxMI Expression Promotes HCC Proliferation and Resistance to Apoptosis. Carcinogenesis 2012, 33, 2250–2259.
- Wada, H.; Nagano, H.; Yamamoto, H.; Yang, Y.; Kondo, M.; Ota, H.; Nakamura, M.; Yoshioka, S.; Kato, H.; Damdinsuren, B.; et al. Expression pattern of angiogenic factors and prognosis after hepatic resection in hepatocellular carcinoma: Importance of angiopoietin-2 and hypoxia-induced factor-1a. Liver Int. 2006, 26, 414–423.
- Dai, C.-X.; Gao, Q.; Qiu, S.-J.; Ju, M.-J.; Cai, M.-Y.; Xu, Y.-F.; Zhou, J.; Zhang, B.-H.; Fan, J. Hypoxia-inducible factor-1 alpha, in association with inflammation, angiogenesis and MYC, is a critical prognostic factor in patients with HCC after surgery. BMC Cancer 2009, 9, 418.
- Wang, D.; Zhang, X.; Lu, Y.; Wang, X.; Zhu, L. Hypoxia inducible factor 1α in hepatocellular carcinoma with cirrhosis: Association with prognosis. Pathol.-Res. Pract. 2018, 214, 1987–1992.
- Yang, S.-L.; Liu, L.-P.; Jiang, J.-X.; Xiong, Z.-F.; He, Q.-J.; Wu, C. The Correlation of Expression Levels of HIF-1 and HIF-2 in Hepatocellular Carcinoma with Capsular Invasion, Portal Vein Tumor Thrombi and Patients’ Clinical Outcome. Jpn. J. Clin. Oncol. 2013, 44, 159–167.
- Liu, L.-P.; Hu, B.; Ye, C.; Ho, R.L.K.; Chen, G.G.; Lai, P.B.-S. HBx mutants differentially affect the activation of hypoxia-inducible factor-1α in hepatocellular carcinoma. Br. J. Cancer 2013, 110, 1066–1073.
- Zheng, S.-S.; Chen, X.-H.; Yin, X.; Zhang, B.-H. Prognostic Significance of HIF-1α Expression in Hepatocellular Carcinoma: A Meta-Analysis. PLoS ONE 2013, 8, e65753.
- Deli, G.; Jin, C.-H.; Mu, R.; Yang, S.; Liang, Y.; Chen, D.; Makuuchi, M. Immunohistochemical assessment of angiogenesis in hepatocellular carcinoma and surrounding cirrhotic liver tissues. World J. Gastroenterol. 2005, 11, 960–963.
- Pelizzaro, F.; Haxhi, S.; Penzo, B.; Vitale, A.; Giannini, E.G.; Sansone, V.; Rapaccini, G.L.; Di Marco, M.; Caturelli, E.; Magalotti, D.; et al. Transarterial Chemoembolization for Hepatocellular Carcinoma in Clinical Practice: Temporal Trends and Survival Outcomes of an Iterative Treatment. Front. Oncol. 2022, 12, 822507.
- Morse, M.A.; Sun, W.; Kim, R.; He, A.R.; Abada, P.B.; Mynderse, M.; Finn, R.S. The Role of Angiogenesis in Hepatocellular Carcinoma. Clin. Cancer Res. 2019, 25, 912–920.
- Finn, R.S.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.-Y.; Kudo, M.; Breder, V.; Merle, P.; Kaseb, A.O.; et al. Atezolizumab plus Bevacizumab in Unresectable Hepatocellular Carcinoma. N. Engl. J. Med. 2020, 382, 1894–1905.
- Yau, T.; Park, J.-W.; Finn, R.S.; Cheng, A.-L.; Mathurin, P.; Edeline, J.; Kudo, M.; Harding, J.J.; Merle, P.; Rosmorduc, O.; et al. Nivolumab versus sorafenib in advanced hepatocellular carcinoma (CheckMate 459): A randomised, multicentre, open-label, phase 3 trial. Lancet Oncol. 2022, 23, 77–90.
- Finn, R.S.; Ryoo, B.-Y.; Merle, P.; Kudo, M.; Bouattour, M.; Lim, H.Y.; Breder, V.; Edeline, J.; Chao, Y.; Ogasawara, S.; et al. Pembrolizumab As Second-Line Therapy in Patients With Advanced Hepatocellular Carcinoma in KEYNOTE-240: A Randomized, Double-Blind, Phase III Trial. J. Clin. Oncol. 2020, 38, 193–202.
- Cramer, T.; Vaupel, P. Severe Hypoxia Is a Typical Characteristic of Human Hepatocellular Carcinoma: Scientific Fact or Fallacy? J. Hepatol. 2022, 76, 975–980.
- Fukumura, D.; Yuan, F.; Monsky, W.L.; Chen, Y.; Jain, R.K. Effect of host microenvironment on the microcirculation of human colon adenocarcinoma. Am. J. Pathol. 1997, 151, 679–688.
- Yang, Z.F.; Poon, R.T. Vascular Changes in Hepatocellular Carcinoma. Anat. Rec. 2008, 291, 721–734.
- Wu, X.-Z.; Xie, G.-R.; Chen, D. Hypoxia and hepatocellular carcinoma: The therapeutic target for hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2007, 22, 1178–1182.
- Iommarini, L.; Porcelli, A.M.; Gasparre, G.; Kurelac, I. Non-Canonical Mechanisms Regulating Hypoxia-Inducible Factor 1 Alpha in Cancer. Front. Oncol. 2017, 7, 286.
- Dvorak, H.F. Vascular Permeability Factor/Vascular Endothelial Growth Factor: A Critical Cytokine in Tumor Angiogenesis and a Potential Target for Diagnosis and Therapy. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2002, 20, 4368–4380.
- Ho, J.W.Y.; Pang, R.W.C.; Lau, C.; Sun, C.K.; Yu, W.C.; Fan, S.T.; Poon, R.T.P. Significance of circulating endothelial progenitor cells in hepatocellular carcinoma. Hepatology 2006, 44, 836–843.
- Jain, R.K. Molecular regulation of vessel maturation. Nat. Med. 2003, 9, 685–693.
- Semela, D.; Dufour, J.-F. Angiogenesis and hepatocellular carcinoma. J. Hepatol. 2004, 41, 864–880.
- Satija, S.; Kaur, H.; Tambuwala, M.M.; Sharma, P.; Vyas, M.; Khurana, N.; Sharma, N.; Bakshi, H.A.; Charbe, N.B.; Zacconi, F.C.; et al. Hypoxia-Inducible Factor (HIF): Fuel for Cancer Progression. Curr. Mol. Pharmacol. 2021, 14, 321–332.
- Wicks, E.E.; Semenza, G.L.; Wicks, E.E.; Semenza, G.L. Hypoxia-Inducible Factors: Cancer Progression and Clinical Translation. J. Clin. Investig. 2022, 132, e159839.
- Kallio, P.J.; Pongratz, I.; Gradin, K.; McGuire, J.; Poellinger, L. Activation of Hypoxia-Inducible Factor 1alpha: Posttranscriptional Regulation and Conformational Change by Recruitment of the Arnt Transcription Factor. Proc. Natl. Acad. Sci. USA 1997, 94, 5667–5672.
- Batie, M.; Frost, J.; Shakir, D.; Rocha, S. Regulation of chromatin accessibility by hypoxia and HIF. Biochem. J. 2022, 479, 767–786.
- Luo, S.-Y.; Wang, J.-Q.; Liu, C.; Gao, X.-M.; Zhang, Y.-B.; Ding, J.; Hou, C.-C.; Zhu, J.-Q.; Lou, B.; Shen, W.-L.; et al. Hif-1α/Hsf1/Hsp70 signaling pathway regulates redox homeostasis and apoptosis in large yellow croaker (Larimichthys crocea) under environmental hypoxia. Zool. Res. 2021, 42, 746–760.
- Liao, S.-H.; Zhao, X.-Y.; Han, Y.-H.; Zhang, J.; Wang, L.-S.; Xia, L.; Zhao, K.-W.; Zheng, Y.; Guo, M.; Chen, G.-Q. Proteomics-based identification of two novel direct targets of hypoxia-inducible factor-1 and their potential roles in migration/invasion of cancer cells. Proteomics 2009, 9, 3901–3912.
- Korbecki, J.; Simińska, D.; Gąssowska-Dobrowolska, M.; Listos, J.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Chronic and Cycling Hypoxia: Drivers of Cancer Chronic Inflammation through HIF-1 and NF-κB Activation: A Review of the Molecular Mechanisms. Int. J. Mol. Sci. 2021, 22, 10701.
- Soni, S.; Padwad, Y.S. HIF-1 in cancer therapy: Two decade long story of a transcription factor. Acta Oncol. 2017, 56, 503–515.
- Infantino, V.; Santarsiero, A.; Convertini, P.; Todisco, S.; Iacobazzi, V. Cancer Cell Metabolism in Hypoxia: Role of HIF-1 as Key Regulator and Therapeutic Target. Int. J. Mol. Sci. 2021, 22, 5703.
- Méndez-Blanco, C.; Fondevila, F.; García-Palomo, A.; González-Gallego, J.; Mauriz, J.L. Sorafenib Resistance in Hepatocarcinoma: Role of Hypoxia-Inducible Factors. Exp. Mol. Med. 2018, 50, 1–9.
- Dong, Z.Z.; Yao, M.; Wang, L.; Wu, W.; Gu, X.; Yao, D.F. Hypoxia-inducible factor-1alpha: Molecular-targeted therapy for hepatocellular carcinoma. Mini-Rev. Med. Chem. 2013, 13, 1295–1304.
- Wilson, G.K.; Tennant, D.A.; McKeating, J.A. Hypoxia inducible factors in liver disease and hepatocellular carcinoma: Current understanding and future directions. J. Hepatol. 2014, 61, 1397–1406.
- Arciero, C.A.; Sigurdson, E.R. Liver-Directed Therapies for Hepatocellular Carcinoma. J. Natl. Compr. Cancer Netw. 2006, 4, 768–774.
- Amini, A.; Moghaddam, S.M.; Morris, D.L.; Pourgholami, M.H. The critical role of vascular endothelial growth factor in tumor angiogenesis. Curr. Cancer Drug Targets 2012, 12, 23–43.
- Li, Y.; Baccouche, B.; Olayinka, O.; Serikbaeva, A.; Kazlauskas, A. The Role of the Wnt Pathway in VEGF/Anti-VEGF-Dependent Control of the Endothelial Cell Barrier. Investig. Opthalmology Vis. Sci. 2021, 62, 17.
- Bayona-Bafaluy, M.P.; Esteban, O.; Ascaso, J.; Montoya, J.; Ruiz-Pesini, E. Oxidative Phosphorylation Inducers Fight Patho-logical Angiogenesis. Drug Discov. Today 2019, 24, 1731–1734.
- Li, J.; Zhou, W.; Chen, W.; Wang, H.; Zhang, Y.; Yu, T. Mechanism of the hypoxia inducible factor 1/hypoxic response element pathway in rat myocardial ischemia/diazoxide post-conditioning. Mol. Med. Rep. 2020, 21, 1527–1536.
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264.
- Levitin, F.; Lee, S.C.-E.S.; Hulme, S.; Rumantir, R.A.; Wong, A.S.; Meester, M.R.; Koritzinsky, M. Oxygen-independent disulfide bond formation in VEGF-A and CA9. J. Biol. Chem. 2021, 296, 100505.
- Kaseb, A.O.; Hanbali, A.; Cotant, M.; Hassan, M.M.; Wollner, I.; Philip, P.A. Vascular Endothelial Growth Factor in the Management of Hepatocellular Carcinoma: A Review of Literature. Cancer 2009, 115, 4895–4906.
- Poon, R.T.-P.; Fan, S.-T.; Wong, J. Clinical Implications of Circulating Angiogenic Factors in Cancer Patients. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2001, 19, 1207–1225.
- Poon, R.T.-P.; Fan, S.-T.; Wong, J. Clinical Significance of Angiogenesis in Gastrointestinal Cancers: A Target for Novel Prognostic and Therapeutic Approaches. Ann. Surg. 2003, 238, 9–28.
- Heldin, C.-H. Targeting the PDGF signaling pathway in tumor treatment. Cell Commun. Signal. 2013, 11, 97.
- Wu, E.; Palmer, N.; Tian, Z.; Moseman, A.P.; Galdzicki, M.; Wang, X.; Berger, B.; Zhang, H.; Kohane, I.S. Comprehensive Dissection of PDGF-PDGFR Signaling Pathways in PDGFR Genetically Defined Cells. PLoS ONE 2008, 3, e3794.
- Chae, Y.K.; Ranganath, K.; Hammerman, P.S.; Vaklavas, C.; Mohindra, N.; Kalyan, A.; Matsangou, M.; Costa, R.; Carneiro, B.; Villaflor, V.M.; et al. Inhibition of the fibroblast growth factor receptor (FGFR) pathway: The current landscape and barriers to clinical application. Oncotarget 2016, 8, 16052–16074.
- Cao, Y.; Cao, R.; Hedlund, E.-M. R Regulation of tumor angiogenesis and metastasis by FGF and PDGF signaling pathways. J. Mol. Med. 2008, 86, 785–789.
- Tsunoda, S.; Nakamura, T.; Sakurai, H.; Saiki, I. Fibroblast growth factor-2-induced host stroma reaction during initial tumor growth promotes progression of mouse melanoma via vascular endothelial growth factor A-dependent neovascularization. Cancer Sci. 2007, 98, 541–548.
- Motoo, Y.; Sawabu, N.; Yamaguchi, Y.; Terada, T.; Nakanuma, Y. Sinusoidal Capillarization of Human Hepatocellular Carcinoma: Possible Promotion by Fibroblast Growth Factor. Oncology 1993, 50, 270–274.
- Lieu, C.; Heymach, J.; Overman, M.; Tran, H.; Kopetz, S. Beyond VEGF: Inhibition of the Fibroblast Growth Factor Pathway and Antiangiogenesis. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 6130–6139.
- Gao, L.; Wang, X.; Tang, Y.; Huang, S.; Hu, C.-A.A.; Teng, Y. FGF19/FGFR4 signaling contributes to the resistance of hepatocellular carcinoma to sorafenib. J. Exp. Clin. Cancer Res. 2017, 36, 8.
- Kaseb, A.; Janku, F.; Bupathi, M. Angiopoietin 2 as a therapeutic target in hepatocellular carcinoma treatment: Current perspectives. OncoTargets Ther. 2014, 7, 1927–1932.
- Maisonpierre, P.C.; Suri, C.; Jones, P.F.; Bartunkova, S.; Wiegand, S.J.; Radziejewski, C.; Compton, D.; McClain, J.; Aldrich, T.H.; Papadopoulos, N.; et al. Angiopoietin-2, a Natural Antagonist for Tie2 That Disrupts in vivo Angiogenesis. Science 1997, 277, 55–60.
- Abou-Alfa, G.K.; Blanc, J.-F.; Miles, S.; Ganten, T.; Trojan, J.; Cebon, J.; Liem, A.K.; Lipton, L.; Gupta, C.; Wu, B.; et al. Phase II Study of First-Line Trebananib Plus Sorafenib in Patients with Advanced Hepatocellular Carcinoma. Oncologist 2017, 22, 780-e65.
- Dallas, N.A.; Samuel, S.; Xia, L.; Fan, F.; Gray, M.J.; Lim, S.J.; Ellis, L.M. Endoglin (CD105): A Marker of Tumor Vasculature and Potential Target for Therapy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 1931–1937.
- Nassiri, F.; Cusimano, M.D.; Scheithauer, B.W.; Rotondo, F.; Fazio, A.; Yousef, G.M.; Syro, L.V.; Kovacs, K.; Lloyd, R. V Endoglin (CD105): A Review of Its Role in Angiogenesis and Tumor Diagnosis, Progression and Therapy. Anticancer Res. 2011, 31, 2283–2290.
- Alev, C.; McIntyre, B.A.S.; Ota, K.; Sheng, G. Dynamic expression of Endoglin, a TGF-beta co-receptor, during pre-circulation vascular development in chick. Int. J. Dev. Biol. 2010, 54, 737–742.
- Li, Y.; Zhai, Z.; Liu, D.; Zhong, X.; Meng, X.; Yang, Q.; Liu, J.; Li, H. CD105 promotes hepatocarcinoma cell invasion and metastasis through VEGF. Tumor Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2015, 36, 737–745.
More
Information
Subjects:
Gastroenterology & Hepatology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
729
Revisions:
2 times
(View History)
Update Date:
26 Jun 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No