Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Analía Bortolozzi | -- | 2706 | 2023-04-24 10:00:43 | | | |
| 2 | Lindsay Dong | + 2 word(s) | 2708 | 2023-04-25 07:57:29 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Miquel-Rio, L.; Sarriés-Serrano, U.; Pavia-Collado, R.; Meana, J.J.; Bortolozzi, A. Role of α-Synuclein in Serotonin System Regulation. Encyclopedia. Available online: https://encyclopedia.pub/entry/43371 (accessed on 26 June 2026).
Miquel-Rio L, Sarriés-Serrano U, Pavia-Collado R, Meana JJ, Bortolozzi A. Role of α-Synuclein in Serotonin System Regulation. Encyclopedia. Available at: https://encyclopedia.pub/entry/43371. Accessed June 26, 2026.
Miquel-Rio, Lluis, Unai Sarriés-Serrano, Rubén Pavia-Collado, J Javier Meana, Analia Bortolozzi. "Role of α-Synuclein in Serotonin System Regulation" Encyclopedia, https://encyclopedia.pub/entry/43371 (accessed June 26, 2026).
Miquel-Rio, L., Sarriés-Serrano, U., Pavia-Collado, R., Meana, J.J., & Bortolozzi, A. (2023, April 24). Role of α-Synuclein in Serotonin System Regulation. In Encyclopedia. https://encyclopedia.pub/entry/43371
Miquel-Rio, Lluis, et al. "Role of α-Synuclein in Serotonin System Regulation." Encyclopedia. Web. 24 April, 2023.
Copy Citation
Pathologically, Parkinson’s disease (PD) is characterized by the loss of dopamine (DA) neurons in the substantia nigra pars compacta (SNc) and the presence of intracellular inclusions, called Lewy bodies and Lewy neurites, composed mostly of α-synuclein (α-Syn). Much of PD research has focused on the role of α-Syn aggregates in the degeneration of SNc DA neurons due to the impact of striatal DA deficits on classical motor phenotypes. However, abundant Lewy pathology is also found in other brain regions including the midbrain raphe nuclei, which may contribute to non-motor symptoms. Indeed, dysfunction of the serotonergic (5-HT) system, which regulates mood and emotional pathways, occurs during the premotor phase of PD.
depression
Parkinson’s disease
α-synuclein
serotonin
raphe nuclei
1. Introduction
Parkinson’s disease (PD) is clinically characterized based on classic motor features including the presence of hypokinesia, rigidity, resting tremor, and impaired postural control [1][2][3]. A wide variety of incapacitating non-motor symptoms are also present over the course of the illness. These non-motor signs include autonomic and neuropsychiatric features such as fatigue, apathy, anxiety and depression, as well as cognitive deficits. Neuropsychiatric symptoms are inherent to the disease and are neither a result nor a side effect of long-term dopaminergic treatment [4][5]. These comorbidities are frequent and can be found in all stages of PD, from the premotor and the early untreated phases of the disease to the advanced stages of PD [6][7][8][9][10][11][12][13]. Among them, depression is one of the most prevalent neuropsychiatric symptoms, ranging from 35 to 50% of patients with PD [14][15][16]. Depressive disorder represents a huge burden on the quality of life in many PD patients, but is frequently undiagnosed and left untreated [17][18][19][20]. Therefore, understanding the neurobiology of depression in PD is critical to achieving the optimal care needed by patients with PD.
While the etiology of PD still remains unclear, one major neuropathological hallmark of PD is the degeneration and subsequent loss of DA neurons in the substantia nigra pars compacta (SNc) leading to prototypic motor deficits [21][22][23]. The SNc involves a neuronal population projecting to the caudate and putamen and is critical for the regulation of basal ganglia circuitry [24][25]. Lewy pathology (LP), which can also be observed across the central, peripheral, and enteric nervous systems (CNS, PNS, and ENS), is another major pathological finding present in about 70% of “clinically typical PD cases” [26][27]. This includes both Lewy bodies (LB) and Lewy neurites (LN), which are composed of a variety of different molecules, proteins, and organelles, including ubiquitin, tubulin, neurofilaments, lipids, and mitochondria. Among them, aggregates of α-synuclein (α-Syn) protein represent one of the main LP components [28][29][30][31][32]. To explain the widespread localization of LP and the onset of the various non-motor symptoms of PD, a critical point to consider is the dysfunction of other neuronal populations and neurotransmitter systems in regions of the CNS and PNS, other than the SNc DA neurons. Indeed, several studies reported LB-associated deficits—most likely occurring even prior to DA neurons—in cholinergic neurons in the pedunculopontine nucleus, nucleus basalis of Meynert and of the dorsal motor nucleus of the vagus, as well as in norepinephrine—NE neurons of the locus coeruleus (LC), and serotonin—5-HT (5-hydroxytryptamine) neurons of the raphe nuclei (RN) [33][34][35]. Furthermore, altered GABAergic and glutamatergic signaling was also reported in the amygdala and several cortical brain regions that may play important roles in the complex cognitive features of PD [34][36][37].
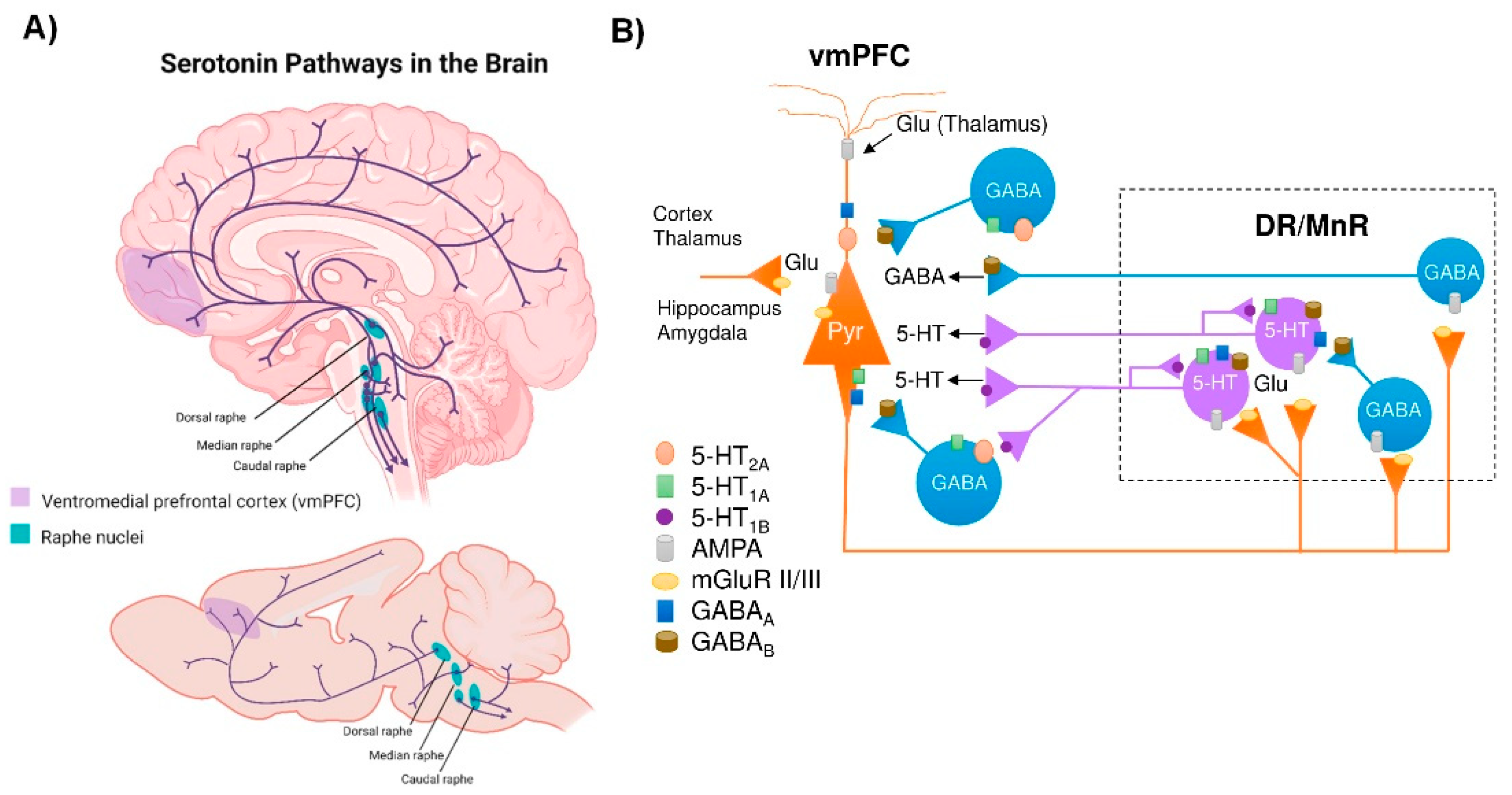
2. Connectivity of the Brain Serotonin System
The brain 5-HT system exerts its widespread effects from a group of relatively small brainstem nuclei known as the RN. Raphe 5-HT-producing neurons send ascending projections to the entire brain as well as descending projections to the spinal cord [38] (Figure 1). These projections form classical synaptic connections, as well as varicosities with no associated postsynaptic structure [39][40]. Upon release, 5-HT acts primarily on G-protein coupled receptors (5-HT1, 5-HT2, 5-HT4, 5-HT5, 5-HT6, 5-HT7, and a single ionotropic receptor 5-HT3) encoded by more than a dozen distinct genes and many more isoforms, which are differentially expressed in the brain [41][42]. Indeed, all brain regions express multiple 5-HT receptors in a receptor subtype-specific pattern [43]. In addition, individual neurons may express several 5-HT receptor subtypes. For instance, pyramidal neurons in layer V of the ventromedial prefrontal cortex (vmPFC) express 5-HT1A and 5-HT2A receptors, which exert opposite effects on neuronal firing activity [44][45]. Hence, the plethora of effects of the brain’s 5-HT system is partly explained by the fact that 5-HT neurons are optimally positioned to affect the activity of a wide range of brain networks.
Figure 1. Central serotonergic pathways. (A) Schematic representation of the raphe nuclei in humans (top) and mice (bottom), which give rise to ascending projections to large regions of the brain, as well as descending projections predominantly innervate the cerebellum and its input structures and to the spinal cord. (B) Diagram showing how the ventromedial prefrontal cortex (vmPFC) and the dorsal and median raphe nuclei (DR and MrR, respectively) are anatomically and functionally connected in both directions. Pyramidal glutamatergic neurons from vmPFC send axons to raphe nuclei, where they form excitatory synapses (AMPA receptors) with 5-HT and GABAergic neurons. Stimulation of glutamatergic neurons in vmPFC primarily triggers inhibitory responses in 5-HT neurons mediated by (i) the activation of local GABAergic circuits that control the activity of 5-HT neurons in the raphe nuclei and (ii) 5-HT1A autoreceptor-dependent self-inhibitory responses following excitatory activation of 5-HT neurons. In addition, DR/MnR 5-HT neurons control the activity of glutamatergic neurons in the vmPFC through inhibitory 5-HT1A receptors and excitatory 5-HT2A receptors expressed in glutamatergic and GABAergic neurons. Similarly, the activity of the vmPFC-DR/MnR pathway may be affected by the activation of 5-HT4 receptors on glutamatergic neurons and 5-HT3 receptors on GABAergic interneurons in the outer layer of the vmPFC (not shown in the diagram). Adapted from [44][45][46].
Among the different raphe nuclei, the dorsal raphe nucleus (DR) is the largest serotonergic nucleus, containing approximately one third of all 5-HT neurons in the brain [38]. As such, the human DR comprises about 250,000 neurons, out of a total of 1011 neurons in the whole brain—approximately 20,000 5-HT-producing neurons in the rat—and its axons branch widely, innervating almost all brain areas. This can be illustrated in the rat cortex, where >106 serotonergic nerve endings/mm3 were noted. In addition, each cortical neuron may receive around 200 varicosities [47]. Unlike cortical and subcortical glutamatergic projection neurons that exhibit precise short- or long-distance connectivity with other neuronal groups [48], DR 5-HT cells send highly divergent ascending projections connecting brain areas with different functions [49].
3. α-Synuclein and Serotonin Neurotransmission
α-Syn is a small, natively unfolded protein belonging to the synuclein family that also encompasses β-synuclein (β-Syn) and γ-synuclein (γ-Syn). These are evolutionarily conserved proteins that have currently only been described in vertebrates, supporting the notion that they regulate some essential physiological functions [50][51][52][53]. Between them, α-Syn is the most studied protein of this family, due to its crucial role in the pathogenesis of PD and other synucleinopathies [54]. This protein is characterized by a remarkable conformational plasticity, adopting different conformations depending on the environment, i.e., neighboring proteins, lipid membranes, redox state, and local pH [55][56][57][58]. In fact, α-Syn adopts a monomeric, random coil conformation in an aqueous solution, while its interaction with lipid membranes drives the transition of the molecule part into α-helical structure. The central unstructured region of α-Syn is involved in fibril formation by converting to well-defined, β-sheet rich secondary structures. These structural and biophysical properties probably hold the key to their normal and abnormal function [59][60]. α-Syn is abundantly expressed in all neuronal types, where it localizes in presynaptic terminals [61][62][63] and modulates synaptic functions [64][65][66]. However, α-Syn is among the last presynaptic proteins to become enriched at the synapse [62] and unlike γ-Syn, it does not seem to be involved in synaptic development [67][68].
In addition to being involved in synaptic vesicular trafficking, α-Syn is also directly engaged in the regulation of monoamine (DA, NE, and 5-HT) neurotransmission homeostasis—β-Syn and γ-Syn are also involved in this regulation, although their role is less known [53][69][70][71][72][73][74]. Monoamine transporters (MAT) are transmembrane proteins solely responsible for the synaptic reuptake of DA, NE and 5-HT, and partly maintain the homeostasis of monoaminergic neurotransmission. MAT are important pharmacological targets in the therapy of various neuropsychiatric diseases, such as anxiety, depression, and suicidal behavior, among others, due to their crucial role within the brain in the replacement of monoamine neurotransmitters [75][76]. Direct interactions between α-Syn and MAT proteins have been described, indicating an important role for the synucleins in regulating MAT function, trafficking and distribution at the synapse.
Previous studies showed that the cell-surface expression and function of the 5-HT transporter (SERT) in co-transfected cells are negatively modulated by α-Syn in a non-Abeta-amyloid component (NAC) domain-dependent manner [71]. In addition, pioneering reports also showed direct interactions of α-Syn-SERT and γ-Syn-SERT proteins in cultured cells and in rat brain tissue, assessed by immunoprecipitation [71][77]. α-Syn-induced modulation of SERT trafficking is microtubule-dependent, as the microtubule-destabilizing agent nocodazole disrupts the effects of α-Syn on SERT function, reversing the inhibition of uptake in co-transfected cells [72]. More recently, in vivo studies indicated that down-regulation of α-Syn expression in raphe 5-HT neurons induced by an antisense oligonucleotide (ASO) leaves an increased synaptic 5-HT concentration, which was dependent on the reduction of SERT activity, as assessed by the selective SERT inhibitor citalopram [74]. The overexpression of α-Syn in raphe nuclei produced the opposite effects, with mice exhibiting a drop in extracellular 5-HT levels that was dependent on SERT function [78].
Moreover, α-Syn is also involved in the vesicular storage of monoamine neurotransmitters by the vesicular monoamine transporter 2 (VMAT2). VMAT2 mobilizes monoamines from the neuronal cytoplasm into vesicles, where they are repackaged for release at synapses [79][80]. VMAT2 co-localizes with α-Syn protein in the Lewy bodies from PD brains [81], and overexpression of α-Syn negatively impairs VMAT2 expression/function, leading to increased levels of cytosolic monoamine in presynaptic terminals, which in turn induce neurotoxicity [69]. These findings suggest that α-Syn may maintain high VMAT2 activity to protect monoamine neurons form cell death [82].
4. Dysfunction of the 5-HT System in PD Patients
The investigation of premotor pathology presents one of the most difficult problems in PD research. Although Braak and colleagues [26][83] proposed a significant premotor phase that may last as long as the symptomatic period, the identification of this phase in clinical practice is elusive. In fact, the profile of PD patients is also associated with diverse symptoms and clinical phenotypes [84]. Cumulative evidence indicates the existence of ongoing pre-SNc DA neurodegeneration during the premotor phase leading to non-motor symptoms, mainly constipation, anxiety and depression, smell loss, and rapid-eye-movement (REM) sleep behavior disorder [85]. A dysfunctional 5-HT system is generally regarded as a risk factor for depression. Consistent with this view, several reports suggest a positive correlation between decreased 5-HT neurotransmission and the severity of depression and anxiety symptoms in PD, most likely caused by pathological changes of the 5-HT neurons in the midbrain raphe nuclei [86][87][88][89][90].
By evaluating SERT availability with positron emission tomography (PET) and single photon emission computed tomography (SPECT) scans using various radioactive ligands, one can assess the integrity of the 5-HT system. The non-specific ligands [123I]β-CIT and [123I]FP-CIT have mostly been employed in in vivo SPECT imaging. Although these ligands have similar affinities for DAT and SERT, their thalamic and midbrain binding are considered to be SERT-specific [91]. Hence, SPECT studies using [123I]β-CIT and [123I]FP-CIT found decreased binding in the thalamus and midbrain of PD patients [92][93][94][95][96]. The PET ligands [11C]-DASB and [11C](+)McN5652 are highly specific for SERT. Using these ligands, several reports indicated reduced binding in different brain regions including the frontal cortex, striatum, and raphe nuclei [97][98][99].
Importantly, neuropathological studies have demonstrated the presence of LBs (α-Syn positive staining) in raphe 5-HT neurons in the early stages of the disease [100][101][102][103]. Previous studies on the propagation of α-Syn proposed that PD begins in the medulla oblongata with LB pathology in the dorsal motor nuclei of the glossopharyngeal and vagal nerves and the adjacent intermediate reticular zone [83][104]. As PD progresses, it is proposed that the LB pathology spreads up the brainstem in an upward direction, affecting the raphe nuclei before reaching the SNc. In late stages, LBs are also found in limbic and cortical brain areas. The caudal groups of the raphe nuclei (e.g., raphe major, raphe obscure, and raphe pallidus) have been widely shown to contain LB-related lesions in the early stages of PD or even before the onset of motor symptoms [83][104][105]. The 5-HT neurons found in the caudal raphe nuclei play a role in a number of autonomic processes, including pain and decreased gastrointestinal motility, which are recognized non-motor symptoms in PD.
5. Dysfunction of the 5-HT System in Animal Models with Overexpression of α-Syn
Abundant evidence suggests that the development of PD may comprise three main phases. The onset of α-Syn buildup in the CNS or PNS/ENS, in the absence of observable clinical symptoms, is referred to as the “preclinical PD” phase. The second phase, often known as the “pre-motor” or “prodromal,” can last for more than 10 years before the disease is clinically diagnosed. It is usually accompanied by the appearance of non-motor symptoms caused in part by pre-SNc abnormalities. During this phase, PD patients may display increased anxiety as early as 16 years prior to disease diagnosis; and depression becomes significantly prevalent among PD patients in the last 3–4 years preceding diagnosis. The third phase is the “motor phase of PD”, which is the one that is clinically visible and easiest to diagnose [106][107]. Understanding the pathophysiological mechanisms underlying non-motor symptoms in PD is important, but requires relevant preclinical animal models. In this sense, one of the main shortcomings of current PD-like animal models is that they focus on DA pathways, which probably do not reflect the complexity underlying the occurrence of these symptoms in patients [108][109].
In addition to the toxin-induced and genetic animal models of PD [109][110][111][112][113][114], in recent decades, an alternative approach to modulate the disease based on the forced expression of wild-type or mutant human α-Syn using (1) transgenic techniques, (2) viral vector mediated transfer of α-Syn, or (3) injection of pathogenic pre-formed α-Syn fibrils (PFFs) has been presented. Thus, an intracellular accumulation of α-Syn in raphe 5-HT neurons and in hippocampal 5-HT fibers, without loss of 5-HT neurons in 12-week-old transgenic mice overexpressing mutant A53T α-Syn, was reported [115]. In parallel, mice showed a reduced 5-HT release and compromised increase in doublecortin+ neuroblasts in the dentate gyrus (DG), indicating a differential neurogenic response [115].
Recently, the adeno-associated virus (AAV)-α-Syn and PFFs models have been specifically adapted for study of α-synucleinopathies using stereotaxic delivery into different brain areas, making them useful tools [116]. Therefore, a model of AAV-induced α-synucleinopathy selectively in 5-HT neurons of rats resulted in progressive degeneration of the 5-HT axon terminals in hippocampus, without the loss of raphe 5-HT neurons [117].
In addition, some studies using cell cultures overexpressing α-Syn showed that 5-hydroxyindoleacetaldehyde (5-HIAL), a 5-HT metabolite product generated by monoamine oxidase (MAO-A), increases α-Syn oligomerization, which may explain the dysfunction of 5-HT neurons in PD [118]. Recent studies also showed the importance of maintaining the integrity of 5-HT systems, as 5-HT itself can affect the growth of amyloid-forming protein fibrils. Indeed, 5-HT or selective serotonin reuptake inhibitors (e.g., escitalopram) activate signaling that alters the processing of α-Syn fibrils as well as amyloid precursor proteins into β-amyloid (Aβ) to prevent protein aggregation by direct binding, and could be beneficial to PD and other neurodegenerative disorders [119][120][121].
6. Conclusions
The frequent occurrence of depression in PD is a prevalent and complex issue. Although often overlooked or underestimated, depression can seriously influence the course of PD and the quality of life of patients. In addition to dopaminergic depletion, several findings highlight the importance of serotonergic degeneration in PD. Thus, changes in 5-HT biochemical markers, LB pathology (α-Syn-positive staining) in raphe nuclei, and structural and functional alterations in the serotonergic system have been described, and it has been shown that these alterations in the serotonergic connectome are mainly associated with the expression of neuropsychiatric symptoms at disease onset. In support of this, the few available animal models demonstrating α-Syn-induced deficits in the serotonergic system recapitulate the mechanisms and early premotor stages of the disease. Altogether, measuring serotonergic integrity might be a useful in vivo tool to use in routines to guide the choice of the pharmacological arsenal in order to alleviate PD-related neuropsychiatric symptoms. Thus, such a measurement could serve as a sensitive marker of PD burden.
References
- Jankovic, J. Parkinson’s disease: Clinical features and diagnosis. J. Neurol. Neurosurg. Psychiatry 2008, 79, 368–376.
- Lima, M.; Martins, E.; Delattre, A.; Proenc, M.; Mori, M.; Carabelli, B. Motor and nonmotor features of Parkinson’s disease: A review of clinical and experimental studies. CNS Neurol. Disord. Drug Targets 2012, 11, 439–449.
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015, 30, 1591–1601.
- Grosch, J.; Winkler, J.; Kohl, Z. Early Degeneration of both dopaminergic and serotonergic axons—A common mechanism in Parkinson’s disease. Front. Cell. Neurosci. 2016, 10, 293.
- Maillet, A.; Krack, P.; Lhommée, E.; Météreau, E.; Klinger, H.; Favre, E.; Le Bars, D.; Schmitt, E.; Bichon, A.; Pelissier, P.; et al. The prominent role of serotonergic degeneration in apathy, anxiety and depression in de novo Parkinson’s disease. Brain 2019, 139, 2486–2502.
- Aarsland, D.; Marsh, L.; Schrag, A. Neuropsychiatric symptoms in Parkinson’s disease. Mov. Disord. 2009, 24, 2175–2186.
- Santangelo, G.; Vitale, C.; Trojano, L.; Longo, K.; Cozzolino, A.; Grossi, D.; Barone, P. Relationship between depression and cognitive dysfunctions in Parkinson’s disease without dementia. J. Neurol. 2009, 256, 632–638.
- Chaudhuri, K.R.; Schapira, A.H. Non-motor symptoms of Parkinson’s disease: Dopaminergic pathophysiology and treatment. Lancet Neurol. 2009, 8, 464–474.
- Thobois, S.; Ardouin, C.; Lhommée, E.; Klinger, H.; Lagrange, C.; Xie, J.; Fraix, V.; Coelho Braga, M.C.; Hassani, R.; Kistner, A.; et al. Non-motor dopamine withdrawal syndrome after surgery for Parkinson’s disease: Predictors and underlying mesolimbic denervation. Brain 2010, 133, 1111–1127.
- De la Riva, P.; Smith, K.; Xie, S.X.; Weintraub, D. Course of psychiatric symptoms and global cognition in early Parkinson disease. Neurology 2014, 83, 1096–1103.
- Dujardin, K.; Langlois, C.; Plomhause, L.; Carette, A.S.; Delliaux, M.; Duhamel, A.; Defebvre, L. Apathy in untreated early-stage Parkinson disease: Relationship with other non-motor symptoms. Mov. Disord 2014, 29, 1796–1801.
- Schrag, A.; Sauerbier, A.; Chaudhuri, K.R. New clinical trials for nonmotor manifestations of Parkinson’s disease. Mov. Disord 2015, 30, 1490–1504.
- Santos García, D.; de Deus Fonticoba, T.; Suárez Castro, E.; Borrué, C.; Mata, M.; Solano Vila, B.; Cots Foraster, A.; Álvarez Sauco, M.; Rodríguez Pérez, A.B.; Vela, L.; et al. Non-motor symptoms burden, mood, and gait problems are the most significant factors contributing to a poor quality of life in non-demented Parkinson’s disease patients: Results from the COPPADIS Study Cohort. Park. Relat. Disord. 2019, 66, 151–157.
- Reijnders, J.S.; Ehrt, U.; Weber, W.E.; Aarsland, D.; Leentjens, F. A systematic review of prevalence studies of depression in Parkinson’s disease. Mov. Disord. 2008, 23, 183–189.
- Aarsland, D.; Påhlhagen, S.; Ballard, C.G.; Ehrt, U.; Svenningsson, P. Depression in Parkinson disease--epidemiology, mechanisms and management. Nat. Rev. Neurol. 2011, 8, 35–47.
- Chuquilín-Arista, F.; Álvarez-Avellón, T.; Menéndez-González, M. Prevalence of Depression and Anxiety in Parkinson Disease and Impact on Quality of Life: A Community-Based Study in Spain. J. Geriatr. Psychiatry Neurol. 2020, 33, 207–213.
- Karlsen, K.H.; Tandberg, E.; Aarsland, D.; Larsen, J.P. Health related quality of life in Parkinson’s disease: A prospective longitudinal study. J. Neurol. Neurosurg. Psychiatry 2000, 69, 584–589.
- Den Oudsten, B.; Van Heck, G.; De Vries, J. Quality of life and related concepts in Parkinson’s disease: A systematic review. Mov. Disord. 2007, 22, 1528–1537.
- Soh, S.; Morris, M.; McGinley, J. Determinants of health-related quality of life in Parkinson’s disease: A systematic review. Park. Relat. Disord. 2011, 17, 1–9.
- Hemmerle, A.; Herman, J.; Seroog, K. Stress, depression and Parkinson’s disease. Exp. Neurol. 2012, 233, 79–86.
- Greffard, S.; Verny, M.; Bonnet, A.M.; Beinis, J.Y.; Gallinari, C.; Meaume, S.; Piette, F.; Hauw, J.J.; Duyckaerts, C. Motor score of the Unified Parkinson Disease Rating Scale as a good predictor of Lewy body-associated neuronal loss in the substantia nigra. Arch. Neurol. 2006, 63, 584–588.
- Halliday, G.; Lees, A.; Stern, M. Milestones in Parkinson’s disease—Clinical and pathologic features. Mov. Disord. 2011, 26, 1015–1021.
- Hornykiewicz, O. 50 years of levodopa. Mov. Disord. 2015, 30, 1008.
- Foffani, G.; Obeso, J.A. A Cortical Pathogenic Theory of Parkinson’s Disease. Neuron 2018, 99, 1116–1128.
- McGregor, M.M.; Nelson, A.B. Circuit Mechanisms of Parkinson’s Disease. Neuron 2019, 101, 1042–1056.
- Braak, H.; Ghebremedhin, E.; Rub, U.; Bratzke, H.; Del Tredici, K. Stages in the development of Parkinson’s disease-related pathology. Cell Tissue Res. 2004, 318, 121–134.
- Braak, H.; Del Tredici, K. Neuropathological staging of brain pathology in sporadic Parkinson’s disease: Separating the wheat from the Chaff. J. Park. Dis. 2017, 7, S71–S85.
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.-Y.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. Alpha-Synuclein in Lewy bodies. Nature 1997, 388, 839–840.
- Dickson, D.W.; Braak, H.; Duda, J.E.; Duyckaerts, C.; Gasser, T.; Halliday, G.M.; Hardy, J.; Leverenz, J.B.; Del Tredici, K.; Wszolek, Z.K.; et al. Neuropathological assessment of Parkinson’s disease: Refining the diagnostic criteria. Lancet Neurol. 2009, 8, 1150–1157.
- Spillantini, M.G.; Goedert, M. Neurodegeneration and the ordered assembly of alpha-synuclein. Cell Tissue Res. 2018, 373, 137–148.
- Shults, C.W. Lewy bodies. Proc. Natl. Acad. Sci. USA 2006, 103, 1661–1668.
- Shahmoradian, S.H.; Lewis, A.J.; Genoud, C.; Hench, J.; Moors, T.E.; Navarro, P.P.; Castaño-Díez, D.; Schweighauser, G.; Graff-Meyer, A.; Goldie, K.N.; et al. Lewy pathology in Parkinson’s disease consists of crowded organelles and lipid membranes. Nat. Neurosci. 2019, 22, 1099–1109.
- Brichta, L.; Greengard, P.; Flajolet, M. Advances in the pharmacological treatment of Parkinson’s disease: Targeting neurotransmitter systems. Trends Neurosci. 2013, 36, 543–554.
- Giguère, N.; Burke Nanni, S.; Trudeau, L.E. On Cell Loss and Selective Vulnerability of Neuronal Populations in Parkinson’s Disease. Front. Neurol. 2018, 9, 455.
- Van Den Berge, N.; Ulusoy, A. Animal models of brain-first and body-first Parkinson’s disease. Neurobiol. Dis. 2022, 163, 105599.
- Harding, A.J.; Stimson, E.; Henderson, J.M.; Halliday, G.M. Clinical correlates of selective pathology in the amygdala of patients with Parkinson’s disease. Brain 2002, 125, 2431–2445.
- Stoyka, L.E.; Arrant, A.E.; Thrasher, D.R.; Russell, D.L.; Freire, J.; Mahoney, C.L.; Narayanan, A.; Dib, A.G.; Standaert, D.G.; Volpicelli-Daley, L.A. Behavioral defects associated with amygdala and cortical dysfunction in mice with seeded α-synuclein inclusions. Neurobiol. Dis. 2020, 134, 104708.
- Hornung, J.P. The human raphe nuclei and the serotonergic system. J. Chem. Neuroanat. 2003, 26, 331–343.
- Descarries, L.; Riad, M.; Parent, M. Ultrastructure of the Serotonin Innervation in the Mammalian Central Nervous System. In Handbook of Behavioral Neurobiology of Serotonin; Müller, C.P., Jacobs, B.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2010.
- Sparta, D.R.; Stuber, G.D. Cartography of serotonergic circuits. Neuron 2014, 83, 513–515.
- Bockaert, J.; Claeysen, S.A.D.; Marin, P. Classification and Signaling Characteristics of 5-HT Receptors. In Handbook of Behavioral Neurobiology of Serotonin; Müller, C.P., Jacobs, B.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2010.
- Weissbourd, B.; Ren, J.; DeLoach, K.E.; Guenthner, C.J.; Miyamichi, K.; Luo, L. Presynaptic partners of dorsal raphe serotonergic and GABAergic neurons. Neuron 2014, 83, 645–662.
- Mengod, G.; Palacios, J.M.; Cortés, R. Cartography of 5-HT1A and 5-HT2A Receptor Subtypes in Prefrontal Cortex and Its Projections. ACS Chem. Neurosci. 2015, 6, 1089–1098.
- Amargós-Bosch, M.; Bortolozzi, A.; Puig, M.V.; Serrats, J.; Adell, A.; Celada, P.; Toth, M.; Mengod, G.; Artigas, F. Co-expression and in vivo interaction of serotonin1A and serotonin2A receptors in pyramidal neurons of prefrontal cortex. Cereb. Cortex 2004, 14, 281–299.
- Santana, N.; Bortolozzi, A.; Serrats, J.; Mengod, G.; Artigas, F. Expression of serotonin1A and serotonin2A receptors in pyramidal and GABAergic neurons of the rat prefrontal cortex. Cereb. Cortex 2004, 14, 1100–1109.
- López-Terrones, E.; Celada, P.; Riga, M.S.; Artigas, F. Preferential In Vivo Inhibitory Action of Serotonin in Rat Infralimbic versus Prelimbic Cortex: Relevance for Antidepressant Treatments. Cereb. Cortex 2022, 32, 3000–3013.
- Janušonis, S. Serotonin in space: Understanding single fibers. ACS Chem. Neurosci. 2017, 8, 893–896.
- Gabbott, P.L.; Warner, T.A.; Jays, P.R.; Salway, P.; Busby, S.J. Prefrontal cortex in the rat: Projections to subcortical autonomic, motor, and limbic centers. J. Comp. Neurol. 2005, 492, 145–177.
- Muzerelle, A.; Scotto-Lomassese, S.; Bernard, J.F.; Soiza-Reilly, M.; Gaspar, P. Conditional anterograde tracing reveals distinct targeting of individual serotonin cell groups (B5-B9) to the forebrain and brainstem. Brain Struct. Funct. 2016, 221, 535–561.
- George, J.M. The synucleins. Genome Biol. 2002, 3, 1–6.
- Li, J.Y.; Henning Jensen, P.; Dahlström, A. Differential localization of α-, β-and γ-synucleins in the rat CNS. Neuroscience 2002, 113, 463–478.
- Longhena, F.; Faustini, G.; Spillantini, M.G.; Bellucci, A. Living in Promiscuity: The Multiple Partners of Alpha-Synuclein at the Synapse in Physiology and Pathology. Int. J. Mol. Sci. 2019, 20, 141.
- Pavia-Collado, R.; Rodríguez-Aller, R.; Alarcón-Arís, D.; Miquel-Rio, L.; Ruiz-Bronchal, E.; Paz, V.; Campa, L.; Galofré, M.; Sgambato, V.; Bortolozzi, A. Up and Down γ-Synuclein Transcription in Dopamine Neurons Translates into Changes in Dopamine Neurotransmission and Behavioral Performance in Mice. Int. J. Mol. Sci. 2022, 23, 1807.
- Goedert, M.; Jakes, R.; Spillantini, M.G. The Synucleinopathies: Twenty Years On. J. Parkinsons Dis. 2017, 7, S51–S69.
- Eliezer, D.; Kutluay, E.; Bussell, R., Jr.; Browne, G. Conformational properties of alpha-synuclein in its free and lipid-associated states. J. Mol. Biol. 2001, 307, 1061–1073.
- Deleersnijder, A.; Gerard, M.; Debyser, Z.; Baekelandt, V. The remarkable conformational plasticity of alpha-synuclein: Blessing or curse? Trends Mol. Med. 2013, 19, 368–377.
- Lv, Z.; Krasnoslobodtsev, A.V.; Zhang, Y.; Ysselstein, D.; Rochet, J.C.; Blanchard, S.C.; Lyubchenko, Y.L. Effect of acidic pH on the stability of alpha-synuclein dimers. Biopolymers 2016, 105, 715–724.
- Galvagnion, C. The Role of Lipids Interacting with alpha-Synuclein in the Pathogenesis of Parkinson’s Disease. J. Park. Dis. 2017, 7, 433–450.
- Sulzer, D.; Edwards, R.H. The physiological role of α-synuclein and its relationship to Parkinson’s Disease. J. Neurochem. 2019, 150, 475–486.
- Vidović, M.; Rikalovic, M.G. Alpha-Synuclein Aggregation Pathway in Parkinson’s Disease: Current Status and Novel Therapeutic Approaches. Cells 2022, 11, 1732.
- Maroteaux, L.; Campanelli, J.T.; Scheller, R.H. Synuclein: A neuron-specific protein localized to the nucleus and presynaptic nerve terminal. J. Neurosci. 1988, 8, 2804–2815.
- Withers, G.S.; George, J.M.; Banker, G.A.; Clayton, D.F. Delayed localization of synelfin (synuclein, NACP) to presynaptic terminals in cultured rat hippocampal neurons. Brain Res. Dev. Brain Res. 1997, 99, 87–94.
- Kahle, P.J.; Neumann, M.; Ozmen, L.; Muller, V.; Jacobsen, H.; Schindzielorz, A.; Okochi, M.; Leimer, U.; van Der Putten, H.; Probst, A.; et al. Subcellular localization of wild-type and Parkinson’s disease-associated mutant alpha -synuclein in human and transgenic mouse brain. J. Neurosci. 2000, 20, 6365–6373.
- Abeliovich, A.; Schmitz, Y.; Farinas, I.; Choi-Lundberg, D.; Ho, W.H.; Castillo, P.E.; Shinsky, N.; Verdugo, J.M.; Armanini, M.; Ryan, A.; et al. Mice lacking alpha-synuclein display functional deficits in the nigrostriatal dopamine system. Neuron 2000, 25, 239–252.
- Nemani, V.M.; Lu, W.; Berge, V.; Nakamura, K.; Onoa, B.; Lee, M.K.; Chaudhry, F.A.; Nicoll, R.A.; Edwards, R.H. Increased expression of alpha-synuclein reduces neurotransmitter release by inhibiting synaptic vesicle reclustering after endocytosis. Neuron 2010, 65, 66–79.
- Burre, J. The Synaptic Function of alpha-Synuclein. J. Parkinsons Dis. 2015, 5, 699–713.
- Burre, J.; Sharma, M.; Sudhof, T.C. Cell Biology and Pathophysiology of alpha-Synuclein. Cold Spring Harb. Perspect. Med. 2018, 8, a024091.
- Fernández-Nogales, M.; López-Cascales, M.T.; Murcia-Belmonte, V.; Escalante, A.; Fernández-Albert, J.; Muñoz-Viana, R.; Barco, A.; Herrera, E. Multiomic Analysis of Neurons with Divergent Projection Patterns Identifies Novel Regulators of Axon Pathfinding. Adv Sci. 2022, 9, e2200615.
- Sidhu, A.; Wersinger, C.; Vernier, P. Does alpha-synuclein modulate dopaminergic synaptic content and tone at the synapse? FASEB J. 2004, 18, 637–647.
- Wersinger, C.; Jeannotte, A.; Sidhu, A. Attenuation of the norepinephrine transporter activity and trafficking via interactions with alpha-synuclein. Eur. J. Neurosci. 2006, 24, 3141–3152.
- Wersinger, C.; Rusnak, M.; Sidhu, A. Modulation of the trafficking of the human serotonin transporter by human alpha-synuclein. Eur. J. Neurosci. 2006, 24, 55–64.
- Oaks, A.W.; Sidhu, A. Synuclein modulation of monoamine transporters. FEBS Lett. 2011, 585, 1001–1006.
- Butler, B.; Saha, K.; Rana, T.; Becker, J.P.; Sambo, D.; Davari, P.; Goodwin, J.S.; Khoshbouei, H. Dopamine Transporter Activity Is Modulated by alpha-Synuclein. J. Biol. Chem. 2015, 290, 29542–29554.
- Alarcón-Arís, D.; Recasens, A.; Galofré, M.; Carballo-Carbajal, I.; Zacchi, N.; Ruiz-Bronchal, E.; Pavia-Collado, R.; Chica, R.; Ferrés-Coy, A.; Santos, M.; et al. Selective α-Synuclein Knockdown in Monoamine Neurons by Intranasal Oligonucleotide Delivery: Potential Therapy for Parkinson’s Disease. Mol. Ther. 2018, 26, 550–567.
- Torres, G.E.; Gainetdinov, R.R.; Caron, M.G. Plasma membrane monoamine transporters: Structure, regulation and function. Nat. Rev. Neurosci. 2003, 4, 13–25.
- Hahn, M.K.; Blakely, R.D. Monoamine transporter gene structure and polymorphisms in relation to psychiatric and other complex disorders. Pharm. J. 2002, 2, 217–235.
- Wersinger, C.; Sidhu, A. Partial regulation of serotonin transporter function by gamma-synuclein. Neurosci. Lett. 2009, 453, 157–161.
- Miquel-Rio, L.; Alarcón-Arís, D.; Torres-López, M.; Cóppola-Segovia, V.; Pavia-Collado, R.; Paz, V.; Ruiz-Bronchal, E.; Campa, L.; Casal, C.; Montefeltro, A.; et al. Human α-synuclein overexpression in mouse serotonin neurons triggers a depressive-like phenotype. Rescue by oligonucleotide therapy. Transl. Psychiatry 2022, 12, 79.
- Narboux-Nême, N.; Sagné, C.; Doly, S.; Diaz, S.L.; Martin, C.B.; Angenard, G.; Martres, M.P.; Giros, B.; Hamon, M.; Lanfumey, L.; et al. Severe serotonin depletion after conditional deletion of the vesicular monoamine transporter 2 gene in serotonin neurons: Neural and behavioral consequences. Neuropsychopharmacology 2011, 36, 2538–2550.
- Eiden, L.E.; Weihe, E. VMAT2: A dynamic regulator of brain monoaminergic neuronal function interacting with drugs of abuse. Ann. N. Y. Acad. Sci. 2011, 1216, 86–98.
- Yamamoto, S.; Fukae, J.; Mori, H.; Mizuno, Y.; Hattori, N. Positive immunoreactivity for vesicular monoamine transporter 2 in Lewy bodies and Lewy neurites in substantia nigra. Neurosci. Lett. 2006, 396, 187–191.
- Taylor, T.N.; Caudle, W.M.; Shepherd, K.R.; Noorian, A.; Jackson, C.R.; Iuvone, P.M.; Weinshenker, D.; Greene, J.G.; Miller, G.W. Nonmotor symptoms of Parkinson’s disease revealed in an animal model with reduced monoamine storage capacity. J. Neurosci. 2009, 29, 8103–8113.
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.; Jansen Steur, E.N.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211.
- De Pablo-Fernández, E.; Lees, A.J.; Holton, J.L.; Warner, T.T. Neuropathological progression of clinical Parkinson disease subtypes. Nat. Rev. Neurol. 2019, 15, 361.
- Iranzo, A.; Tolosa, E.; Gelpi, E.; Molinuevo, J.L.; Valldeoriola, F.; Serradell, M.; Sanchez-Valle, R.; Vilaseca, I.; Lomeña, F.; Vilas, D.; et al. Neurodegenerative disease status and post-mortem pathology in idiopathic rapid-eye-movement sleep behaviour disorder: An observational cohort study. Lancet Neurol. 2013, 12, 443–453.
- Boileau, I.; Warsh, J.J.; Guttman, M.; Saint-Cyr, J.A.; McCluskey, T.; Rusjan, P.; Houle, S.; Wilson, A.A.; Meyer, J.H.; Kish, S.J. Elevated serotonin transporter binding in depressed patients with Parkinson’s disease: A preliminary PET study with DASB. Mov. Disord. 2008, 23, 1776–1780.
- Politis, M.; Wu, K.; Loane, C.; Turkheimer, F.E.; Molloy, S.; Brooks, D.J.; Piccini, P. Depressive symptoms in PD correlate with higher 5-HTT binding in raphe and limbic structures. Neurology 2010, 75, 1920–1927.
- Ballanger, B.; Klinger, H.; Eche, J.; Lerond, J.; Vallet, A.E.; Le Bars, D.; Tremblay, L.; Sgambato-Faure, V.; Broussolle, E.; Thobois, S. Role of serotonergic 1A receptor dysfunction in depression associated with Parkinson’s disease. Mov. Disord. 2012, 27, 84–89.
- Ohno, Y.; Shimizu, S.; Tokudome, K.; Kunisawa, N.; Sasa, M. New insights into the therapeutic role of the serotonergic system in Parkinson’s disease. Prog. Neurobiol. 2015, 134, 104–121.
- Wilson, H.; Dervenoulas, G.; Pagano, G.; Koros, C.; Yousaf, T.; Picillo, M.; Polychronis, S.; Simitsi, A.; Giordano, B.; Chappell, Z.; et al. Serotonergic pathology and disease burden in the premotor and motor phase of A53T α-synuclein parkinsonism: A cross-sectional study. Lancet Neurol. 2019, 18, 748–759.
- Tan, S.K.; Hartung, H.; Sharp, T.; Temel, Y. Serotonin-dependent depression in Parkinson’s disease: A role for the subthalamic nucleus? Neuropharmacology 2011, 61, 387–399.
- Haapaniemi, T.H.; Ahonen, A.; Torniainen, P.; Sotaniemi, K.A.; Myllylä, V.V. β-CIT SPECT demonstrates decreased brain dopamine and serotonin transporter levels in untreated parkinsonian patients. Mov. Disord. 2001, 16, 124–130.
- Kim, S.E.; Choi, J.Y.; Choe, Y.S.; Choi, Y.; Lee, W.Y. Serotonin transporters in the midbrain of Parkinson’s disease patients: A study with 123I-β-CIT SPECT. J. Nucl. Med. 2003, 44, 870–876.
- Caretti, V.; Stoffers, D.; Winogrodzka, A.; Isaias, I.U.; Costantino, G.; Pezzoli, G.; Ferrarese, C.; Antonini, A.; Wolters, E.C.; Booij, J. Loss of thalamic serotonin transporters in early drug-naïve Parkinson’s disease patients is associated with tremor: An β-CIT SPECT study. J. Neural Transm. 2008, 115, 721–729.
- Roselli, F.; Pisciotta, N.M.; Pennelli, M.; Aniello, M.S.; Gigante, A.; De Caro, M.F.; Ferrannini, E.; Tartaglione, B.; Niccoli-Asabella, A.; Defazio, G.; et al. Midbrain SERT in degenerative parkinsonisms: A 123I-FP-CIT SPECT study. Mov. Disord. 2010, 25, 1853–1859.
- Qamhawi, Z.; Towey, D.; Shah, B.; Pagano, G.; Seibyl, J.; Marek, K.; Borghammer, P.; Brooks, D.J.; Pavese, N. Clinical correlates of raphe serotonergic dysfunction in early Parkinson’s disease. Brain 2015, 138, 2964–2973.
- Kerenyi, L.; Ricaurte, G.A.; Schretlen, D.J.; McCann, U.; Varga, J.; Mathews, W.B.; Ravert, H.T.; Dannals, R.F.; Hilton, J.; Wong, D.F.; et al. Positron emission tomography of striatal serotonin transporters in Parkinson disease. Arch. Neurol. 2003, 60, 1223–1229.
- Guttman, M.; Boileau, I.; Warsh, J.; Saint-Cyr, J.A.; Ginovart, N.; McCluskey, T.; Houle, S.; Wilson, A.; Mundo, E.; Rusjan, P.; et al. Brain serotonin transporter binding in non-depressed patients with Parkinson’s disease. Eur. J. Neurol. 2007, 14, 523–528.
- Albin, R.L.; Koeppe, R.A.; Bohnen, N.I.; Wernette, K.; Kilbourn, M.A.; Frey, K.A. Spared caudal brainstem SERT binding in early Parkinson’s disease. J. Cereb. Blood Flow Metab. 2008, 28, 441–444.
- Halliday, G.M.; Blumbergs, P.C.; Cotton, R.G.; Blessing, W.W.; Geffen, L.B. Loss of brainstem serotonin- and substance P-containing neurons in Parkinson’s disease. Brain Res. 1990, 510, 104–107.
- Halliday, G.M.; Li, Y.W.; Blumbergs, P.C.; Joh, T.H.; Cotton, R.G.; Howe, P.R.; Blessing, W.W.; Geffen, L.B. Neuropathology of immunohistochemically identified brainstem neurons in Parkinson’s disease. Ann. Neurol. 1990, 27, 373–385.
- Kish, S.J.; Tong, J.; Hornykiewicz, O.; Rajput, A.; Chang, L.J.; Guttman, M.; Furukawa, Y. Preferential loss of serotonin markers in caudate versus putamen in Parkinson’s disease. Brain 2008, 131, 120–131.
- Politis, M.; Wu, K.; Loane, C.; Quinn, N.P.; Brooks, D.J.; Oertel, W.H.; Björklund, A.; Lindvall, O.; Piccini, P. Serotonin neuron loss and nonmotor symptoms continue in Parkinson’s patients treated with dopamine grafts. Sci. Transl. Med. 2012, 4, 128ra41.
- Del Tredici, K.; Rüb, U.; De Vos, R.A.; Bohl, J.R.; Braak, H. Where does Parkinson’s disease pathology begin in the brain? J. Neuropathol. Exp. Neurol. 2002, 61, 413–426.
- Parkkinen, L.; Pirttila, T.; Alafuzoff, I. Applicability of current staging/categorization of alpha-synuclein pathology and their clinical relevance. Acta Neuropathol. 2008, 115, 399–407.
- Akhtar, R.S.; Stern, M.B. New concepts in the early and preclinical detection of Parkinson’s disease: Therapeutic implications. Expert. Rev. Neurother. 2012, 12, 1429–1438.
- Darweesh, S.K.; Verlinden, V.J.; Stricker, B.H.; Hofman, A.; Koudstaal, P.J.; Ikram, M.A. Trajectories of prediagnostic functioning in Parkinson’s disease. Brain 2017, 140, 429–441.
- Faivre, F.; Joshi, A.; Bezard, E.; Barrot, M. The hidden side of Parkinson’s disease: Studying pain, anxiety and depression in animal models. Neurosci. Biobehav. Rev. 2019, 96, 335–352.
- Bové, J.; Perier, C. Neurotoxin-based models of Parkinson’s disease. Neuroscience 2012, 211, 51–76.
- Koprich, J.B.; Kalia, L.V.; Brotchie, J.M. Animal models of α-synucleinopathy for Parkinson disease drug development. Nat. Rev. Neurosci. 2017, 18, 515–529.
- Volta, M.; Melrose, H. LRRK2 mouse models: Dissecting the behavior, striatal neurochemistry and neurophysiology of PD pathogenesis. Biochem. Soc. Trans. 2017, 45, 113–122.
- Bastías-Candia, S.; Zolezzi, J.M.; Inestrosa, N.C. Revisiting the Paraquat-Induced Sporadic Parkinson’s Disease-Like Model. Mol. Neurobiol. 2019, 56, 1044–1055.
- Creed, R.B.; Goldberg, M.S. New Developments in Genetic rat models of Parkinson’s Disease. Mov. Disord. 2018, 33, 717–729.
- Francardo, V. Modeling Parkinson’s disease and treatment complications in rodents: Potentials and pitfalls of the current options. Behav. Brain Res. 2018, 352, 142–150.
- Deusser, J.; Schmidt, S.; Ettle, B.; Plötz, S.; Huber, S.; Müller, C.P.; Masliah, E.; Winkler, J.; Kohl, Z. Serotonergic dysfunction in the A53T alpha-synuclein mouse model of Parkinson’s disease. J. Neurochem. 2015, 135, 589–597.
- Björklund, A.; Nilsson, F.; Mattsson, B.; Hoban, D.B.; Parmar, M.A. Combined α-Synuclein/Fibril (SynFib) Model of Parkinson-Like Synucleinopathy Targeting the Nigrostriatal Dopamine System. J. Parkinsons Dis. 2022. preprint.
- Wan, O.W.; Shin, E.; Mattsson, B.; Caudal, D.; Svenningsson, P.; Björklund, A. α-Synuclein induced toxicity in brain stem serotonin neurons mediated by an AAV vector driven by the tryptophan hydroxylase promoter. Sci. Rep. 2016, 6, 26285.
- Jinsmaa, Y.; Cooney, A.; Sullivan, P.; Sharabi, Y.; Goldstein, D.S. The serotonin aldehyde, 5-HIAL, oligomerizes alpha-synuclein. Neurosci. Lett. 2015, 590, 134–137.
- Falsone, S.F.; Leitinger, G.; Karner, A.; Kungl, A.J.; Kosol, S.; Cappai, R.; Zangger, K. The neurotransmitter serotonin interrupts α-synuclein amyloid maturation. Biochim. Biophys. Acta 2011, 1814, 553–561.
- Tin, G.; Mohamed, T.; Shakeri, A.; Pham, A.T.; Rao, P.P.N. Interactions of Selective Serotonin Reuptake Inhibitors with β-Amyloid. ACS Chem. Neurosci. 2019, 10, 226–234.
- Cirrito, J.R.; Wallace, C.E.; Yan, P.; Davis, T.A.; Gardiner, W.D.; Doherty, B.M.; King, D.; Yuede, C.M.; Lee, J.M.; Sheline, Y.I. Effect of escitalopram on Aβ levels and plaque load in an Alzheimer mouse model. Neurology 2020, 95, e2666–e2674.
More
Information
Subjects:
Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.0K
Revisions:
2 times
(View History)
Update Date:
25 Apr 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No