Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Analía Bortolozzi and Version 2 by Lindsay Dong.
Pathologically, Parkinson’s disease (PD) is characterized by the loss of dopamine (DA) neurons in the substantia nigra pars compacta (SNc) and the presence of intracellular inclusions, called Lewy bodies and Lewy neurites, composed mostly of α-synuclein (α-Syn). Much of PD research has focused on the role of α-Syn aggregates in the degeneration of SNc DA neurons due to the impact of striatal DA deficits on classical motor phenotypes. However, abundant Lewy pathology is also found in other brain regions including the midbrain raphe nuclei, which may contribute to non-motor symptoms. Indeed, dysfunction of the serotonergic (5-HT) system, which regulates mood and emotional pathways, occurs during the premotor phase of PD.
- depression
- Parkinson’s disease
- α-synuclein
- serotonin
- raphe nuclei
1. Introduction
Parkinson’s disease (PD) is clinically characterized based on classic motor features including the presence of hypokinesia, rigidity, resting tremor, and impaired postural control [1][2][3][1,2,3]. A wide variety of incapacitating non-motor symptoms are also present over the course of the illness. These non-motor signs include autonomic and neuropsychiatric features such as fatigue, apathy, anxiety and depression, as well as cognitive deficits. Neuropsychiatric symptoms are inherent to the disease and are neither a result nor a side effect of long-term dopaminergic treatment [4][5][4,5]. These comorbidities are frequent and can be found in all stages of PD, from the premotor and the early untreated phases of the disease to the advanced stages of PD [6][7][8][9][10][11][12][13][6,7,8,9,10,11,12,13]. Among them, depression is one of the most prevalent neuropsychiatric symptoms, ranging from 35 to 50% of patients with PD [14][15][16][14,15,16]. Depressive disorder represents a huge burden on the quality of life in many PD patients, but is frequently undiagnosed and left untreated [17][18][19][20][17,18,19,20]. Therefore, understanding the neurobiology of depression in PD is critical to achieving the optimal care needed by patients with PD.
While the etiology of PD still remains unclear, one major neuropathological hallmark of PD is the degeneration and subsequent loss of DA neurons in the substantia nigra pars compacta (SNc) leading to prototypic motor deficits [21][22][23][21,22,23]. The SNc involves a neuronal population projecting to the caudate and putamen and is critical for the regulation of basal ganglia circuitry [24][25][24,25]. Lewy pathology (LP), which can also be observed across the central, peripheral, and enteric nervous systems (CNS, PNS, and ENS), is another major pathological finding present in about 70% of “clinically typical PD cases” [26][27][26,27]. This includes both Lewy bodies (LB) and Lewy neurites (LN), which are composed of a variety of different molecules, proteins, and organelles, including ubiquitin, tubulin, neurofilaments, lipids, and mitochondria. Among them, aggregates of α-synuclein (α-Syn) protein represent one of the main LP components [28][29][30][31][32][28,29,30,31,32]. To explain the widespread localization of LP and the onset of the various non-motor symptoms of PD, a critical point to consider is the dysfunction of other neuronal populations and neurotransmitter systems in regions of the CNS and PNS, other than the SNc DA neurons. Indeed, several studies reported LB-associated deficits—most likely occurring even prior to DA neurons—in cholinergic neurons in the pedunculopontine nucleus, nucleus basalis of Meynert and of the dorsal motor nucleus of the vagus, as well as in norepinephrine—NE neurons of the locus coeruleus (LC), and serotonin—5-HT (5-hydroxytryptamine) neurons of the raphe nuclei (RN) [33][34][35][33,34,35]. Furthermore, altered GABAergic and glutamatergic signaling was also reported in the amygdala and several cortical brain regions that may play important roles in the complex cognitive features of PD [34][36][37][34,36,37].
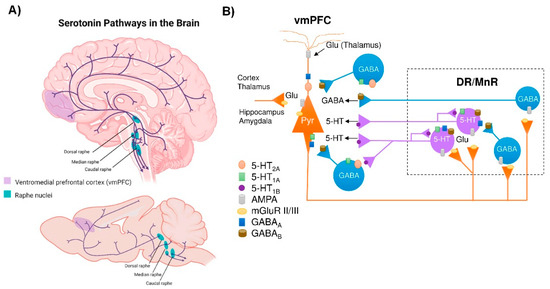
2. Connectivity of the Brain Serotonin System
The brain 5-HT system exerts its widespread effects from a group of relatively small brainstem nuclei known as the RN. Raphe 5-HT-producing neurons send ascending projections to the entire brain as well as descending projections to the spinal cord [38][47] (Figure 1A). These projections form classical synaptic connections, as well as varicosities with no associated postsynaptic structure [39][40][48,49]. Upon release, 5-HT acts primarily on G-protein coupled receptors (5-HT1, 5-HT2, 5-HT4, 5-HT5, 5-HT6, 5-HT7, and a single ionotropic receptor 5-HT3) encoded by more than a dozen distinct genes and many more isoforms, which are differentially expressed in the brain [41][42][50,51]. Indeed, all brain regions express multiple 5-HT receptors in a receptor subtype-specific pattern [43][52]. In addition, individual neurons may express several 5-HT receptor subtypes. For instance, pyramidal neurons in layer V of the ventromedial prefrontal cortex (vmPFC) express 5-HT1A and 5-HT2A receptors, which exert opposite effects on neuronal firing activity [44][45][53,54]. Hence, the plethora of effects of the brain’s 5-HT system is partly explained by the fact that 5-HT neurons are optimally positioned to affect the activity of a wide range of brain networks.Figure 1. Central serotonergic pathways. (A) Schematic representation of the raphe nuclei in humans (top) and mice (bottom), which give rise to ascending projections to large regions of the brain, as well as descending projections predominantly innervate the cerebellum and its input structures and to the spinal cord. (B) Diagram showing how the ventromedial prefrontal cortex (vmPFC) and the dorsal and median raphe nuclei (DR and MrR, respectively) are anatomically and functionally connected in both directions. Pyramidal glutamatergic neurons from vmPFC send axons to raphe nuclei, where they form excitatory synapses (AMPA receptors) with 5-HT and GABAergic neurons. Stimulation of glutamatergic neurons in vmPFC primarily triggers inhibitory responses in 5-HT neurons mediated by (i) the activation of local GABAergic circuits that control the activity of 5-HT neurons in the raphe nuclei and (ii) 5-HT1A autoreceptor-dependent self-inhibitory responses following excitatory activation of 5-HT neurons. In addition, DR/MnR 5-HT neurons control the activity of glutamatergic neurons in the vmPFC through inhibitory 5-HT1A receptors and excitatory 5-HT2A receptors expressed in glutamatergic and GABAergic neurons. Similarly, the activity of the vmPFC-DR/MnR pathway may be affected by the activation of 5-HT4 receptors on glutamatergic neurons and 5-HT3 receptors on GABAergic interneurons in the outer layer of the vmPFC (not shown in the diagram). Adapted from [44][45][46][53,54,78].
Among the different raphe nuclei, the dorsal raphe nucleus (DR) is the largest serotonergic nucleus, containing approximately one third of all 5-HT neurons in the brain [38][47]. As such, the human DR comprises about 250,000 neurons, out of a total of 1011 neurons in the whole brain—approximately 20,000 5-HT-producing neurons in the rat—and its axons branch widely, innervating almost all brain areas. This can be illustrated in the rat cortex, where >106 serotonergic nerve endings/mm3 were noted. In addition, each cortical neuron may receive around 200 varicosities [47][55]. Unlike cortical and subcortical glutamatergic projection neurons that exhibit precise short- or long-distance connectivity with other neuronal groups [48][56], DR 5-HT cells send highly divergent ascending projections connecting brain areas with different functions [49][57].
3. α-Synuclein and Serotonin Neurotransmission
α-Syn is a small, natively unfolded protein belonging to the synuclein family that also encompasses β-synuclein (β-Syn) and γ-synuclein (γ-Syn). These are evolutionarily conserved proteins that have currently only been described in vertebrates, supporting the notion that they regulate some essential physiological functions [50][51][52][53][82,83,84,85]. Between them, α-Syn is the most studied protein of this family, due to its crucial role in the pathogenesis of PD and other synucleinopathies [54][86]. This protein is characterized by a remarkable conformational plasticity, adopting different conformations depending on the environment, i.e., neighboring proteins, lipid membranes, redox state, and local pH [55][56][57][58][87,88,89,90]. In fact, α-Syn adopts a monomeric, random coil conformation in an aqueous solution, while its interaction with lipid membranes drives the transition of the molecule part into α-helical structure. The central unstructured region of α-Syn is involved in fibril formation by converting to well-defined, β-sheet rich secondary structures. These structural and biophysical properties probably hold the key to their normal and abnormal function [59][60][91,92]. α-Syn is abundantly expressed in all neuronal types, where it localizes in presynaptic terminals [61][62][63][93,94,95] and modulates synaptic functions [64][65][66][96,97,98]. However, α-Syn is among the last presynaptic proteins to become enriched at the synapse [62][94] and unlike γ-Syn, it does not seem to be involved in synaptic development [67][68][99,100].
In addition to being involved in synaptic vesicular trafficking, α-Syn is also directly engaged in the regulation of monoamine (DA, NE, and 5-HT) neurotransmission homeostasis—β-Syn and γ-Syn are also involved in this regulation, although their role is less known [53][69][70][71][72][73][74][85,113,114,115,116,117,118]. Monoamine transporters (MAT) are transmembrane proteins solely responsible for the synaptic reuptake of DA, NE and 5-HT, and partly maintain the homeostasis of monoaminergic neurotransmission. MAT are important pharmacological targets in the therapy of various neuropsychiatric diseases, such as anxiety, depression, and suicidal behavior, among others, due to their crucial role within the brain in the replacement of monoamine neurotransmitters [75][76][119,120]. Direct interactions between α-Syn and MAT proteins have been described, indicating an important role for the synucleins in regulating MAT function, trafficking and distribution at the synapse.
Previous studies showed that the cell-surface expression and function of the 5-HT transporter (SERT) in co-transfected cells are negatively modulated by α-Syn in a non-Abeta-amyloid component (NAC) domain-dependent manner [71][115]. In addition, pioneering reports also showed direct interactions of α-Syn-SERT and γ-Syn-SERT proteins in cultured cells and in rat brain tissue, assessed by immunoprecipitation [71][77][115,121]. α-Syn-induced modulation of SERT trafficking is microtubule-dependent, as the microtubule-destabilizing agent nocodazole disrupts the effects of α-Syn on SERT function, reversing the inhibition of uptake in co-transfected cells [72][116]. More recently, in vivo studies indicated that down-regulation of α-Syn expression in raphe 5-HT neurons induced by an antisense oligonucleotide (ASO) leaves an increased synaptic 5-HT concentration, which was dependent on the reduction of SERT activity, as assessed by the selective SERT inhibitor citalopram [74][118]. The overexpression of α-Syn in raphe nuclei produced the opposite effects, with mice exhibiting a drop in extracellular 5-HT levels that was dependent on SERT function [78][110].
Moreover, α-Syn is also involved in the vesicular storage of monoamine neurotransmitters by the vesicular monoamine transporter 2 (VMAT2). VMAT2 mobilizes monoamines from the neuronal cytoplasm into vesicles, where they are repackaged for release at synapses [79][80][122,123]. VMAT2 co-localizes with α-Syn protein in the Lewy bodies from PD brains [81][124], and overexpression of α-Syn negatively impairs VMAT2 expression/function, leading to increased levels of cytosolic monoamine in presynaptic terminals, which in turn induce neurotoxicity [69][113]. These findings suggest that α-Syn may maintain high VMAT2 activity to protect monoamine neurons form cell death [82][125].