Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Majid Shakeri | -- | 2913 | 2023-04-19 15:37:25 | | | |
| 2 | Sirius Huang | Meta information modification | 2913 | 2023-04-20 10:06:40 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Ebrahimi, R.; Ebrahimi, M.; Shakeri, M. Heavy Metals-Induced Oxidative Stress in Poultry. Encyclopedia. Available online: https://encyclopedia.pub/entry/43242 (accessed on 01 August 2026).
Ebrahimi R, Ebrahimi M, Shakeri M. Heavy Metals-Induced Oxidative Stress in Poultry. Encyclopedia. Available at: https://encyclopedia.pub/entry/43242. Accessed August 01, 2026.
Ebrahimi, Rohollah, Mahdi Ebrahimi, Majid Shakeri. "Heavy Metals-Induced Oxidative Stress in Poultry" Encyclopedia, https://encyclopedia.pub/entry/43242 (accessed August 01, 2026).
Ebrahimi, R., Ebrahimi, M., & Shakeri, M. (2023, April 19). Heavy Metals-Induced Oxidative Stress in Poultry. In Encyclopedia. https://encyclopedia.pub/entry/43242
Ebrahimi, Rohollah, et al. "Heavy Metals-Induced Oxidative Stress in Poultry." Encyclopedia. Web. 19 April, 2023.
Copy Citation
Oxidative stress in chickens caused by dietary, environmental, and pathological variables influences how well chickens perform as well as the quality of meat and eggs. Lead (Pb) and cadmium (Cd) are two examples of heavy metals that are harmful for chicken health. They can cause oxidative stress by increasing the production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) and blocking antioxidants from protecting cells from increased amounts of free radicals. The oxidative state of heavy metals, their interactions with endogenous antioxidants, and chemical processes all affect how hazardous they are to the body.
heavy metals
phytogenic compounds
oxidative stress
poultry
1. Introduction
Poultry production has one of the fastest rates of development among animal industries, and it greatly enhances nutrition and food security. Over the past few years, broiler chicken genetics, feed conversion ratio, fat reduction and breast size, as well as egg production and quality, have all seen considerable improvements [1][2][3]. In the poultry industry, the cost of feed makes up a sizeable amount of the total cost of producing meat and eggs. The general condition of a chicken’s health, especially the gastrointestinal tract (GIT), plays an important role in the final cost of the product. The GIT of the birds is exposed to a variety of factors through feed and environment, which affect chicken health [4][5]. The main environmental pollution entry points for undesired compounds into the food chain include air, water, soil, and feed. According to Sharma et al., [6], heavy metals are one of the environmental contaminants that enter animal feed from natural, industrial, and agricultural sources. Lead (Pb) can be absorbed through the skin; however, it is primarily absorbed through the digestive and respiratory systems [7]. According to the data that are now available, environmental pollution, the availability of supplements and concentrates, or the technology and equipment used during manufacturing make the heavy metal contamination of animal feeds inevitable [8]. Health risks to both humans and animals can result from hazardous compounds including cadmium (Cd), Pb, and arsenic (As) entering the food chain [9]. Pb is neurotoxic and can harm hemopoiesis, renal function, the nervous system, and the gastrointestinal system in addition to impairing metabolism [10]. Pb distribution in the body first depends on blood flow into various organs, while about 95% of Pb is deposited in skeletal bones as insoluble phosphate [11]. Due to the fact that it comes from the environment and various dietary sources, the diet could be a source of Cd pollution [10]. Cd is then transferred from other animals to people through the food chain, where it damages the lungs, liver, and kidneys and causes hypertension [8]. According to one study, broiler meat products (e.g., burgers and frankfurters) had higher levels of Cd, Cu, Mn, Ni, Pb, and Co compared to raw meat and table eggs [12]. Overall, the findings demonstrated that eating chicken meat products posed a health risk to consumers since Pb and Ni levels were four and seven times higher than the permitted maximum limit, respectively, while the levels of Pb and Ni may vary from a country to a country. Additionally, even in very low amounts, arsenic is a highly hazardous element in feed and drinking water. The hazardous dose of As for poultry is between 40 and 50 mg/kg of chicken feed; at 40 mg/kg, egg production is reduced, but at 50 mg/kg, feed consumption is reduced [13]. Heavy metals are one of these elements; when they interact with the chicken’s body, they create free radicals. If there are too many of these reactive oxygen species/reactive nitrogen species (ROS/RNS), a type of unstable molecule that contains oxygen/nitrogen and that easily reacts with other molecules in a cell, the DNA, protein, and lipid structures, this may harm and impair cell function (Figure 1). At specific concentrations, both ROS and RNS function as homeostasis-related signaling molecules. Oxygen metabolism produces ROS, such as superoxide, hydrogen peroxide, and hydroxyl radicals, which are further balanced by the rates of oxidant generation and oxidant removal. Certain cells of the intestinal mucosa and submucosal areas express the RNS that are byproducts of nitric oxide synthases (NOS). Nitric oxide radical (NO), created when the NOS converts arginine to citrulline, is essential for cellular processes such as neurotransmission and immunomodulation. Yet, excessive NO generation harms the intestinal mucous membrane and impairs nutritional absorption. Due to the high concentration of polyunsaturated fatty acids in cell membrane lipids and lipoproteins, both ROS and RNS can contribute to lipid peroxidation. Lipid peroxidation produces 4-hydroxynonenal as its byproduct, which worsens oxidative damage to cell membranes, compromises cell signaling, causes mitochondrial dysfunction, and disturbs redox equilibrium.
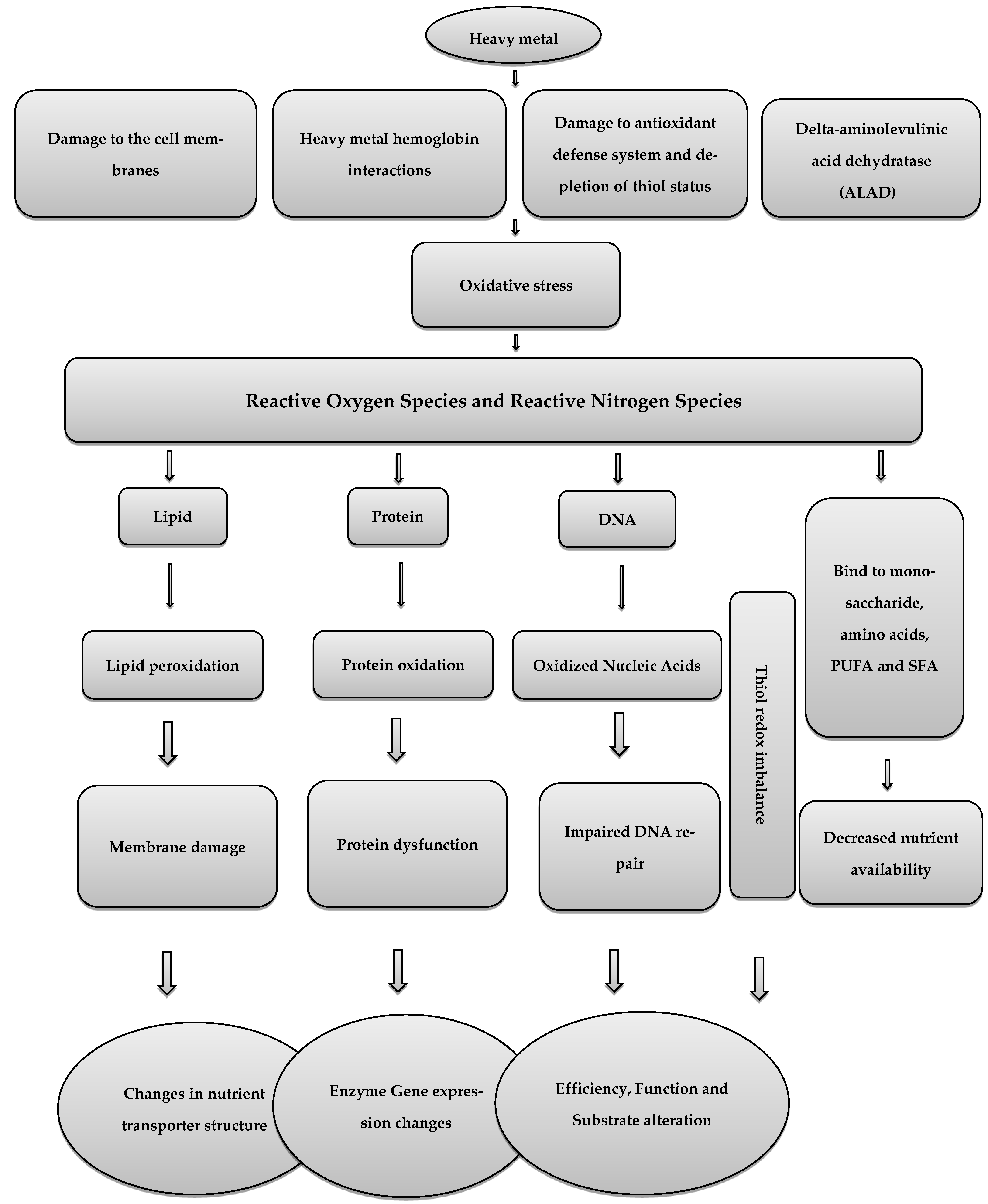
Figure 1. The effect of oxidative stress caused by heavy metals on the enterocytes of poultry.
2. Heavy Metals and Oxidative Stress in Poultry
2.1. Pb
Because of Pb’s non-biodegradable nature and the ongoing use of items related to it, there are severe challenges to global food security [14]. Poultry is one of the most important agricultural sectors, and each year, more than 50 billion birds are raised for food [15]. On broiler growth, even 1.0 mg/kg of trace Pb concentration in feed can have a significant effect, including linear decreases in body weight gain, linear decrease in delta-aminolevulinic acid dehydratase (ALAD) activity and dose-related increase in Pb in the blood, kidney, liver and tibia in broiler chickens [16]. Two healthy elements of eggs, ovalbumin and phosvitin, have been proven to bind a variety of metals [17], increasing the risk of heavy metal accumulation in eggs. The rate of Pb transmission from female birds to eggs was assumed to be highly associated with the quantities in livers [18]. A previous investigation suggested that Pb may alter the way Ca is digested, making it easier for Pb to be incorporated into eggshells [19]. Therefore, egg production may offer a means for female birds to eliminate Pb other than through urine or feces [20]. A study showed that Pb can lead to oxidative stress in chickens and hasten biological component oxidation processes by generating free radicals [21]. The key contributor to the ionic mechanism of Pb poisoning, which eventually interferes with cell metabolism, is the ability of Pb metal ions to replace other bivalent cations such as Ca2+, Mg2+, and Fe2+ as well as monovalent cations such as Na+. The ionic mechanism of Pb poisoning significantly affects a wide range of biological activities, including cell adhesion, intracellular and intercellular communication, protein folding, maturation, apoptosis, ionic transport, enzyme control, and neurotransmitter release. Protein kinase C, a gene that regulates neuronal excitation and memory storage, is impacted by the fact that Pb can replace calcium even in picomolar quantities [22]. Pb ions may alter the morphology and operation of the mitochondria, which would cause cell death. The induction of the mitochondrial permeability transition pore (mPTP) is significantly influenced by the production of ROS [23]. The accumulation of ROS brought on by Pb, however, has been shown to impact energy metabolism and alter DNA through point mutations, rearrangements, and fragmentation [23]. It has also proposed that the disruptive effects of Pb ions on mitochondrial respiratory complexes are the root cause of Pb (II)-induced liver damage. The release of cytochrome C and opening the permeability transition pore (PTP), which results in mitochondrial dysfunction and possibly cell death signaling, is what causes the toxicity [24]. Overall, Pb can cause oxidative damage in various organs by having a direct impact on the peroxidation of membrane lipids and lowering antioxidant properties [25]. The antioxidant parameters GPx, CAT, SOD, glutathione S—Transferase (GST), and glutathione (GSH) are considerably decreased by exposure to Pb, whereas the oxidative parameters MDA and H2O2 are increased [26]. In Pb poisoning, caspase-3 is activated once mitochondrial cytochrome C is released, which inhibits Bcl-2 and causes apoptosis [27]. Additionally, blocking the release of neurotransmitters and inhibiting the N-methyl-D-aspartate (NMDA) receptor may cause Pb neurotoxicity and cognitive impairment [28].
2.2. Cd and As
Hazardous heavy metal Cd is widely present in nature and has a variety of adverse effects for human/animal health. The creation of nickel-cadmium batteries, electroplating, burning of fossil fuels, and mining waste are all examples of human activities that contribute to the widespread occurrence of Cd [9]. Due of these factors, almost everything we ingest, such as food, drinks, and air, includes Cd [10]. Continuous exposure to low levels of Cd causes daily buildup in several tissues and adverse effects on various organs [8]. Due to its chemical similarity, it can mimic and substitute some nutritional metals in a range of biological structures [29]. Recent research suggests that Cd may be dangerous to health even at low dosages because of its propensity to accumulate [30]. For animal husbandry in some countries, Cd contamination has been a problem, and in some instances, the amount of Cd in manure and animal feed can even approach 130 mg/kg [31]. Long-term exposure to Cd can result in hepatotoxicity, renal failure, and neurotoxicity [32]. The rapid impacts of Cd exposure in vivo include the generation of ROS in the mitochondrial electron transfer chain being stimulated, NADPH oxidase activity in the plasma being inhibited, and physiological antioxidants such as glutathione being depleted. The buildup of Cd in tissues could slow the pace of growth [33]. The kidney and liver are the main tissue targets for Cd toxicity. Cd produces oxidative stress by increasing the formation of free radicals. Increased ROS can lead to lipid peroxidation, DNA oxidation, sulfhydryl depletion, and a disruption of calcium homoeostasis [34]. The specific mechanism of Cd poisoning is unknown, although its effects on cells are widely known [35]. Cd concentration increases by a factor of 3000 when it binds to metallothionein or other proteins with a high cysteine content. After causing hepatotoxicity in the liver, the cysteine–metallothionein complex builds up in the renal tissue of the kidney and causes nephrotoxicity. Aspartate, glutamate, histidine, and cysteine are also among the ligands that Cd can bind, and this can cause an iron deficiency [36]. Due to their similar oxidation states, Cd and zinc could have been substituted in metallothionein to prevent it from acting as a cell’s free radical scavenger. Cd and Pb both have the capacity to mimic significant metals and/or replace them if necessary. These ions attach to calmodulin [37], protein kinase C [38], troponin C, and synaptic proteins [39] that include magnesium-, zinc-, and calcium-specific binding sites. The most stable complexes are created when these so-called “soft metals” join forces with the mixed N-S donor atom ligand. Recent research on mouse renal tubular epithelial cells has shown that Cd can trigger the release of Ca (II) from ER reserves through the phospholipase C (PLC)-IP3 pathway, which participates in the formation of ROS [40]. Cd can bind to lipids, proteins, and nucleic acids after entering the body. Thiol (-SH) groups are frequently used for binding to enzymes and proteins, and they modify cysteine residues in proteins. This type of protein inactivation has the power to disturb the intracellular redox balance. As a result, liver harm develops as a result of an unbalanced antioxidant defense [7]. Additionally, it has been thought that Cd indirectly produces ROS. The antioxidant defense of cells could be overwhelmed by O2, hydroxyl (OH), and the NO radicals that Cd could indirectly produce [41]. This might be the result of cellular proteins containing more Cd than iron and copper. The buildup of too much Fe and Cu is the cause of the oxidative stress. Additionally, replacing the necessary minerals throws off the cellular metabolism of the cell. Alternately, Cd might interfere with glutathione, causing oxidative stress to develop [42].
Long-term exposure to arsenic has been demonstrated to be harmful to the liver, lung epithelial transformed cells, and skin, and it has also been shown that arsenic alters multiple cellular pathways, including cytokine expression, apoptosis promotion and resistance, and increased oxidative stress [43]. These alterations result in the manifestation of disease [44]. In chicken hearts, subchronic arsenism-induced oxidative stress is also thought to trigger inflammation, and it is believed that ROS overproduction activates the NF-B pathway, which in turn causes an increase in the expression of pro-inflammatory mediators such as TNF, prostaglandin E synthase, cyclooxygenase-2, and inducible nitric oxide synthase [44]. Arsenic pathogenesis is characterized by oxidative damage caused by ROS. Arsenic also causes morphological abnormalities in the mitochondria’s structural integrity. Cells are more susceptible to the harmful effects of arsenic as a result of glutathione-depleting substances paired with cascade mechanisms of free radical generation resulting from the superoxide radical. The formation of ROS/RNS, including peroxyl radicals, the superoxide radical, singlet oxygen, hydroxyl radicals via the Fenton reaction, hydrogen peroxide, the dimethylarsenic radical, the dimethylarsenic peroxyl radical, and/or oxidant-induced DNA damage, is increased in both humans and animals exposed to As [45].
2.3. Mitigation of Oxidative Stress in Poultry
Dietary antioxidants help to keep the intestinal mucosa healthy while lowering intestinal free radicals [46]. Numerous studies indicate that oxidative stress rids birds of various pathogenic and welfare problems [47]. Therefore, the poultry industry must design a practical strategy to prevent oxidative stress [4]. To reduce oxidative distress in poultry [48], a variety of dietary therapies are available based on the best supplementation of antioxidative vitamins (E, A, C, and B2) and micronutrients (Se, Cu, and Zn). Researchers have recently become interested in the chemicals discussed below because of their chelating and growth-stimulating functions, as well as the antioxidant qualities of useful plant components.
The liver is known to be protected by polyphenolic compounds from a variety of xenobiotics, including Pb and diethyl nitrosamine, which can cause hepatotoxicity [49]. Over the past ten years, many in vitro and in vivo studies have suggested that tea and tea polyphenols have potent antioxidant activity as well as a variety of other potentially useful medicinal properties, including the capacity to inhibit tumor growth, metastasis and carcinogenesis in various animals. The main polyphenolic components in tea are catechins. Tea catechins are effective scavengers of superoxide, hydrogen peroxide, hydroxyl radicals, and nitric oxide in various compounds. They were also able to bind with metals, thanks to their catechol structure [50]. The protective effect tea catechins exert on oxidative damage in HepG2 cells exposed to Pb may be due to their capacity to bind metal ions and scavenge free radicals [51]. According to [52], experiments are being conducted to find ways to lessen the harmful effects of Cd and Pb on the body by chelating these metals with nutrients, which reduces their absorption by tissues or boosts the body’s oxidative capacity. However, there are presently no effective techniques to reduce the levels of Cd and Pb in food and hence lessen the risk of oxidative stress being induced in internal organs. Supplemental lycopene and fucoxanthin shield rat kidney, bone, and brain tissue against the effects of Cd-mediated oxidative stress [53].
Recent studies have used astaxanthin, a red carotenoid pigment found in some marine species and a potent antioxidant without provitamin-A activity, to improve rooster sperm quality [54]. According to [55], at a concentration of 25 mg/kg, astaxanthin nanoparticles act as a potent antioxidant to shield rooster testes from the oxidative stress caused by Cd injection and maintain the post-thawing quality of rooster sperm. As with other carotenoids, astaxanthin has a low bioavailability despite being a highly lipophilic molecule. As a result, an astaxanthin nano preparation that is more stable and bioavailable has been developed. However, the study investigates how astaxanthin, a lipid-soluble carotenoid, protects against Cd-induced damage to rooster testis and decreased sperm quality. Additionally, it has been noted in the literature that astaxanthin has around 100 times the antioxidant activity of alpha-tocopherol and about 10 times the antioxidant activity of other carotenoids, including zeaxanthin, lutein, canthaxanthin, and beta-carotene [56].
It has been demonstrated that the polyphenolic molecule resveratrol has strong antioxidant properties that can protect against hydroxyl and superoxide radicals produced by heavy metals [57]. Additionally, it may increase the activity of GSH-Px, CAT, GST, SOD, and nicotinamide adenine while activating the key transcription factors that regulate the response to antioxidants (erythroid-derived nuclear factor) [58]. It might also maintain glutathione in its reduced state by blocking the creation of glutathione disulfide. This would prevent the oxidation of macromolecules, inhibit the peroxidation of the apolipoprotein B protein, and protect cells from the onslaught of free radicals [59]. Resveratrol has also been demonstrated to lessen oxidative stress and increase antioxidant status in chickens when provided as a dietary supplement [60]. Additionally, yucca’s resveratrol and other phenolic components may reduce lipid peroxidation and stop the generation of blood platelet free radicals (LPO) [61]. It might also maintain glutathione in its reduced state by stopping the manufacture of glutathione disulfide [62]. Chelation therapy, antioxidant therapy, and the consumption of natural food components are among the therapeutic and preventive approaches available to combat As toxicity. Natural dietary substances and medications with a plant-based origin provide effective and progressive treatment from As-mediated toxicity without causing any distinct side effects. Due in significant part to their robust antioxidant properties, bioactive compounds have generated considerable interest in their potential advantages [63]. In a recent study, 34 medicinal plants and 14 natural compounds, largely in preclinical trials and a few in clinical research, demonstrated considerable protection against As toxicity [64]. According to [65], Allium sativum, Curcuma longa, Silybum marianum, as well as various herbal fibers and algae, are the most effective medicinal plants for treating As toxicity. Organosulfur-containing vegetables are helpful in removing arsenic from the liver. Dialyl sulphide, an organosulfur natural substance present in garlic (Allium sativum), has been shown to reduce toxicity and As-induced mitochondrial dysfunction in mice [66]. Crude extracts of Viscum album and Allium sativum were investigated for their ability to counteract in vivo experimentally generated As toxicity [67]. Experimental evidence of the antioxidant properties of members of the Lamiaceae family was found in Ocimum sanctum leaf extract [68]. Due to its antioxidant activity, Silybum marianum’s flavonolignan, silibinin, exerts beneficial effects on rats exposed to As [69].
References
- Bacou, E.; Walk, C.; Rider, S.; Litta, G.; Perez-Calvo, E. Dietary oxidative distress: A review of nutritional challenges as models for poultry, swine and fish. Antioxidants 2021, 10, 525.
- Shakeri, M.; Oskoueian, E.; Le, H.H.; Shakeri, M. Strategies to combat heat stress in broiler chickens: Unveiling the roles of selenium, vitamin E and vitamin C. Vet. Sci. 2020, 7, 71.
- Shakeri, M.; Le, H.H. Deleterious Effects of Heat Stress on Poultry Production: Unveiling the Benefits of Betaine and Polyphenols. Poultry 2022, 1, 147–156.
- Mishra, B.; Jha, R. Oxidative stress in the poultry gut: Potential challenges and interventions. Front. Vet. Sci. 2019, 6, 60.
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Zhao, W.; Le, H.H.; McQuade, R.; Furness, J.B.; Dunshea, F.R. Dietary betaine improves intestinal barrier function and ameliorates the impact of heat stress in multiple vital organs as measured by evans blue dye in broiler chickens. Animals 2019, 10, 38.
- Sharma, R.K.; Agrawal, M. Biological effects of heavy metals: An overview. J. Environ. Biol. 2005, 26, 301–313.
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic mechanisms of five heavy metals: Mercury, lead, chromium, cadmium, and arsenic. Front. Pharmacol. 2021, 12, 643972.
- Khafaga, A.F.; El-Hack, A.; Mohamed, E.; Taha, A.E.; Elnesr, S.S.; Alagawany, M. The potential modulatory role of herbal additives against Cd toxicity in human, animal, and poultry: A review. Environ. Sci. pollut. Res. 2019, 26, 4588–4604.
- Nai, G.A.; Marin, F.F.; Queiroz, L.M.M.; Estrella, M.P.S. Respiratory tract cadmium-induced injuries—poisoning via intake and water pH could influence their genesis? An experimental study in rats. Comp. Clin. Path. 2017, 26, 997–1002.
- Schaefer, H.R.; Dennis, S.; Fitzpatrick, S. Cadmium: Mitigation strategies to reduce dietary exposure. J. Food Sci. 2020, 85, 260–267.
- Papanikolaou, N.C.; Hatzidaki, E.G.; Belivanis, S.; Tzanakakis, G.N.; Tsatsakis, A.M. Lead toxicity update. A brief review. Med. Sci. Monit. 2005, 11, RA329.
- Korish, M.A.; Attia, Y.A. Evaluation of heavy metal content in feed, litter, meat, meat products, liver, and table eggs of chickens. Animals 2020, 10, 727.
- Baloš, M.Ž.; Jakšić, S.; Pelić, D.L. The role, importance and toxicity of arsenic in poultry nutrition. Worlds Poult. Sci. J. 2019, 75, 375–386.
- Zheng, S.; Wang, Q.; Yuan, Y.; Sun, W. Human health risk assessment of heavy metals in soil and food crops in the Pearl River Delta urban agglomeration of China. Food Chem. 2020, 316, 126213.
- Okoye, P.; Ajiwe, V.; Okeke, O.; Ujah, I.; Asalu, U.; Okeke, D. Estimation of heavy metal levels in the muscle, gizzard, liver and kidney of broiler, layer and local (cockerel) chickens raised within Awka metropolis and its environs, Anambra state, south eastern Nigeria. J. Environ. Prot. 2015, 6, 609.
- Bakalli, R.; Pesti, G.; Ragland, W. The magnitude of lead toxicity in broiler chickens. Vet. Hum. Toxicol. 1995, 37, 15–19.
- Dobrzański, Z.; Gorecki, H.; Chojnacka, K.; Gorecka, H.; Synowiec, M. Effect of dietary humic preparations on the content of trace elements in hens’ eggs. Am. J. Agric. Biol. Sci. 2007, 2, 234–240.
- Abduljaleel, S.A.; Shuhaimi-Othman, M. Metals concentrations in eggs of domestic avian and estimation of health risk from eggs consumption. J. Biol. Sci. 2011, 11, 448–453.
- Hunton, P. Research on eggshell structure and quality: An historical overview. Braz. J. Poult. Sci. 2005, 7, 67–71.
- Dauwe, T.; Janssens, E.; Bervoets, L.; Blust, R.; Eens, M. Heavy-metal concentrations in female laying great tits (Parus major) and their clutches. Arch. Environ. Contam. Toxicol. 2005, 49, 249–256.
- Ebrahimi, R.; Faseleh Jahromi, M.; Liang, J.B.; Soleimani Farjam, A.; Shokryazdan, P.; Idrus, Z. Effect of dietary lead on intestinal nutrient transporters mRNA expression in broiler chickens. BioMed Res. Int. 2015, 2015, 149745.
- Flora, S.; Mittal, M.; Mehta, A. Heavy metal induced oxidative stress & its possible reversal by chelation therapy. Indian J. Med. Res. 2008, 128, 501.
- Ye, F.; Li, X.; Li, F.; Li, J.; Chang, W.; Yuan, J.; Chen, J. Cyclosporin A protects against Lead neurotoxicity through inhibiting mitochondrial permeability transition pore opening in nerve cells. Neurotoxicology 2016, 57, 203–213.
- Ma, L.; Liu, J.-Y.; Dong, J.-X.; Xiao, Q.; Zhao, J.; Jiang, F.-L. Toxicity of Pb2+ on rat liver mitochondria induced by oxidative stress and mitochondrial permeability transition. Toxicol. Res. 2017, 6, 822–830.
- Gurer-Orhan, H.; Sabır, H.U.; Özgüneş, H. Correlation between clinical indicators of lead poisoning and oxidative stress parameters in controls and lead-exposed workers. Toxicology 2004, 195, 147–154.
- Wang, J.; Zhu, H.; Yang, Z.; Liu, Z. Antioxidative effects of hesperetin against lead acetate-induced oxidative stress in rats. Indian J. Pharmacol. 2013, 45, 395.
- Liu, C.-M.; Ma, J.-Q.; Sun, Y.-Z. Puerarin protects rat kidney from lead-induced apoptosis by modulating the PI3K/Akt/eNOS pathway. Toxicol. Appl. Pharmacol. 2012, 258, 330–342.
- Neal, A.P.; Guilarte, T.R. Molecular neurobiology of lead (Pb2+): Effects on synaptic function. Mol. Neurobiol. 2010, 42, 151–160.
- Zhu, M.; Li, H.; Bai, L.; Wang, L.; Zou, X. Histological changes, lipid metabolism, and oxidative and endoplasmic reticulum stress in the liver of laying hens exposed to cadmium concentrations. Poult. Sci. 2020, 99, 3215–3228.
- Branca, J.J.V.; Morucci, G.; Pacini, A. Cadmium-induced neurotoxicity: Still much ado. Neural Regen. Res. 2018, 13, 1879.
- Li, Y.-x.; Xiong, X.; Chun-ye, L.; Feng-song, Z.; Wei, L.; Wei, H. Cadmium in animal production and its potential hazard on Beijing and Fuxin farmlands. J. Hazard. Mater. 2010, 177, 475–480.
- Fan, R.; Hu, P.-c.; Wang, Y.; Lin, H.-y.; Su, K.; Feng, X.-s.; Wei, L.; Yang, F. Betulinic acid protects mice from cadmium chloride-induced toxicity by inhibiting cadmium-induced apoptosis in kidney and liver. Toxicol. Lett. 2018, 299, 56–66.
- Akyolcu, M.; Ozcelik, D.; Dursun, S.; Toplan, S.; Kahraman, R. Accumulation of cadmium in tissue and its effect on live performance. In Proceedings of the Journal de Physique IV (Proceedings), Porto, Portugal, 8–10 September 2003; pp. 33–36.
- Stohs, S.J.; Bagchi, D. Oxidative mechanisms in the toxicity of metal ions. Free Radic. Biol. Med. 1995, 18, 321–336.
- Patrick, L. Toxic metals and antioxidants: Part II. The role of antioxidants in arsenic and cadmium toxicity. Altern. Med. Rev. 2003, 8, 106–128.
- Castagnetto, J.M.; Hennessy, S.W.; Roberts, V.A.; Getzoff, E.D.; Tainer, J.A.; Pique, M.E. MDB: The metalloprotein database and browser at the Scripps Research Institute. Nucleic Acids Res. 2002, 30, 379–382.
- Kern, M.; Wisniewski, M.; Cabell, L.; Audesirk, G. Inorganic lead and calcium interact positively in activation of calmodulin. Neurotoxicology 2000, 21, 353–363.
- Sun, X.; Tian, X.; Tomsig, J.L.; Suszkiw, J.B. Analysis of differential effects of Pb2+ on protein kinase C isozymes. Toxicol. Appl. Pharmacol. 1999, 156, 40–45.
- Marchetti, C. Role of calcium channels in heavy metal toxicity. Int. Sch. Res. Not. 2013, 2013, 184360.
- Gu, J.; Dai, S.; Liu, Y.; Liu, H.; Zhang, Y.; Ji, X.; Yu, F.; Zhou, Y.; Chen, L.; Tse, W.K.F. Activation of Ca2+-sensing receptor as a protective pathway to reduce Cadmium-induced cytotoxicity in renal proximal tubular cells. Sci. Rep. 2018, 8, 1–13.
- Sarkar, A.; Ravindran, G.; Krishnamurthy, V. A brief review on the effect of cadmium toxicity: From cellular to organ level. Int. J. Biotechnol. Res. 2013, 3, 17–36.
- Liu, J.; Qu, W.; Kadiiska, M.B. Role of oxidative stress in cadmium toxicity and carcinogenesis. Toxicol. Appl. Pharmacol. 2009, 238, 209–214.
- Flora, S.J. Arsenic-induced oxidative stress and its reversibility. Free Radic. Biol. Med. 2011, 51, 257–281.
- Zhao, H.; He, Y.; Li, S.; Sun, X.; Wang, Y.; Shao, Y.; Hou, Z.; Xing, M. Subchronic arsenism-induced oxidative stress and inflammation contribute to apoptosis through mitochondrial and death receptor dependent pathways in chicken immune organs. Oncotarget 2017, 8, 40327.
- Jomova, K.; Jenisova, Z.; Feszterova, M.; Baros, S.; Liska, J.; Hudecova, D.; Rhodes, C.; Valko, M. Arsenic: Toxicity, oxidative stress and human disease. J. Appl. Toxicol. 2011, 31, 95–107.
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant defence systems and oxidative stress in poultry biology: An update. Antioxidants 2019, 8, 235.
- Estévez, M. Oxidative damage to poultry: From farm to fork. Poult. Sci. 2015, 94, 1368–1378.
- Lauridsen, C. From oxidative stress to inflammation: Redox balance and immune system. Poult. Sci. 2019, 98, 4240–4246.
- McGill, M.R.; Du, K.; Weemhoff, J.L.; Jaeschke, H. Critical review of resveratrol in xenobiotic-induced hepatotoxicity. Food Chem. Toxicol. 2015, 86, 309–318.
- Rice-Evans, C. Flavonoid antioxidants. Curr. Med. Chem. 2001, 8, 797–807.
- Chen, L.; Yang, X.; Jiao, H.; Zhao, B. Tea catechins protect against lead-induced cytotoxicity, lipid peroxidation, and membrane fluidity in HepG2 cells. Toxicol. Sci. 2002, 69, 149–156.
- Winiarska-Mieczan, A. Protective effect of tea against lead and cadmium-induced oxidative stress—A review. Biometals 2018, 31, 909–926.
- Mężyńska, M.; Brzóska, M.M.; Rogalska, J.; Piłat-Marcinkiewicz, B. Extract from Aronia melanocarpa L. berries prevents cadmium-induced oxidative stress in the liver: A study in a rat model of low-level and moderate lifetime human exposure to this toxic metal. Nutrients 2018, 11, 21.
- Fang, Y.; Zhong, R.; Chen, L.; Feng, C.; Sun, H.; Zhou, D. Effects of astaxanthin supplementation on the sperm quality and antioxidant capacity of ram semen during liquid storage. Small Rumin. Res. 2015, 130, 178–182.
- Najafi, D.; Taheri, R.A.; Najafi, A.; Shamsollahi, M.; Alvarez-Rodriguez, M. Effect of astaxanthin nanoparticles in protecting the post-thawing quality of rooster sperm challenged by cadmium administration. Poult. Sci. 2020, 99, 1678–1686.
- Miki, W. Biological functions and activities of animal carotenoids. Pure Appl. Chem. 1991, 63, 141–146.
- Rubiolo, J.; Vega, F. Resveratrol protects primary rat hepatocytes against necrosis induced by reactive oxygen species. Biomed. Pharmacother. 2008, 62, 606–612.
- Alagawany, M.; El-Hack, A.; Mohamed, E.; El-Kholy, M.S. Productive performance, egg quality, blood constituents, immune functions, and antioxidant parameters in laying hens fed diets with different levels of Yucca schidigera extract. Environ. Sci. pollut. Res. 2016, 23, 6774–6782.
- Chakravarthi, S.; Jessop, C.E.; Bulleid, N.J. The role of glutathione in disulphide bond formation and endoplasmic-reticulum-generated oxidative stress. EMBO Rep. 2006, 7, 271–275.
- Wang, C.; Zhao, F.; Li, Z.; Jin, X.; Chen, X.; Geng, Z.; Hu, H.; Zhang, C. Effects of resveratrol on growth performance, intestinal development, and antioxidant status of broilers under heat stress. Animals 2021, 11, 1427.
- Olas, B.; Wachowicz, B.; Stochmal, A.; Oleszek, W. Inhibition of oxidative stress in blood platelets by different phenolics from Yucca schidigera Roezl. bark. Nutrition 2003, 19, 633–640.
- Nurdiana, S.; Goh, Y.M.; Ahmad, H.; Dom, S.M.; Syimal’ain Azmi, N.; Noor Mohamad Zin, N.S.; Ebrahimi, M. Changes in pancreatic histology, insulin secretion and oxidative status in diabetic rats following treatment with Ficus deltoidea and vitexin. BMC Complement. Altern. Med. 2017, 17, 1–17.
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233.
- Bhattacharya, S. Medicinal plants and natural products in amelioration of arsenic toxicity: A short review. Pharm. Biol. 2017, 55, 349–354.
- Mehrandish, R.; Rahimian, A.; Shahriary, A. Heavy metals detoxification: A review of herbal compounds for chelation therapy in heavy metals toxicity. J. Herbmed Pharmacol. 2019, 8, 69–77.
- Das, B.; Chaudhuri, K. Amelioration of sodium arsenite induced toxicity by diallyl disulfide, a bioactive component of garlic: The involvement of antioxidants and the chelate effect. RSC Adv. 2014, 4, 20964–20973.
- Adegboyega, A.; Odunola, O. The modulatory effects of aqueous extracts of Viscum album and garlic on sodium arsenite induced toxicity in Wistar albino rat. J. Chem. Pharm. Res. 2012, 4, 4698–4701.
- Sharmila Banu, G.; Kumar, G.; Murugesan, A. Effect of ethanolic leaf extract of Trianthema portulacastrum L. on aflatoxin induced hepatic damage in rats. Indian J. Clin. Biochem. 2009, 24, 414–418.
- Bjørklund, G.; Rahaman, M.S.; Shanaida, M.; Lysiuk, R.; Oliynyk, P.; Lenchyk, L.; Chirumbolo, S.; Chasapis, C.T.; Peana, M. Natural dietary compounds in the treatment of arsenic toxicity. Molecules 2022, 27, 4871.
More
Information
Subjects:
Agriculture, Dairy & Animal Science
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.5K
Revisions:
2 times
(View History)
Update Date:
20 Apr 2023
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No