Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Tan, J.; Zhao, H.; Li, J.; Gong, Y.; Li, X. The Features of Magnaporthe oryzae Genome. Encyclopedia. Available online: https://encyclopedia.pub/entry/43137 (accessed on 25 July 2026).
Tan J, Zhao H, Li J, Gong Y, Li X. The Features of Magnaporthe oryzae Genome. Encyclopedia. Available at: https://encyclopedia.pub/entry/43137. Accessed July 25, 2026.
Tan, Jinyi, Haikun Zhao, Josh Li, Yihan Gong, Xin Li. "The Features of Magnaporthe oryzae Genome" Encyclopedia, https://encyclopedia.pub/entry/43137 (accessed July 25, 2026).
Tan, J., Zhao, H., Li, J., Gong, Y., & Li, X. (2023, April 17). The Features of Magnaporthe oryzae Genome. In Encyclopedia. https://encyclopedia.pub/entry/43137
Tan, Jinyi, et al. "The Features of Magnaporthe oryzae Genome." Encyclopedia. Web. 17 April, 2023.
Copy Citation
Magnaporthe oryzae (synonym of Pyricularia oryzae), which causes rice blast disease, is a plant pathogenic fungus belonging to the Ascomycota phylum.
Magnaporthe oryzae
plant fungal pathogen
rice blast
1. Introduction
In 1879, Magnaporthe oryzae was initially named Trichothecium griseum by Cooke without a detailed description [1]. In the following century, several names were given to the fungus based on the teleomorph and anamorph stages, such as Ceratosphaeria grisea by Herber (1971) and Dactylaria oryzae by Sawada (1917). In 1990, based on host specificity, physiological differences, and genetic evidence, Rossman corrected its name to Pyricularia oryzae [2]. P. oryzae was used to refer to the asexual stage, and the sexual stage was named Magnaporthe grisea by Couch and Kohn (2002). Additional phylogenetic analysis and interstrain fertility tests showed that M. grisea should be assigned to Digitaria (crabgrass)-infecting isolates, whereas M. oryzae causes damage to rice and other important crops in the Poaceae family, such as millet and wheat [3].
Magnaporthe oryzae is one of the most devastating agricultural pathogens since it primarily attacks cultivated rice (Oryza sativa), an important staple food feeding over 50% of the world’s population [4][5]. Its infection can be destructive under favorable conditions, contributing to 10–30% of the annual rice yield loss [6][7]. Given that it can affect both temperate and tropical rice growth, the disease has been widely distributed in 85 countries with various environmental conditions [7][8][9]. As a consequence, it is ranked the most damaging fungal pathogen in the world [9].
Magnaporthe oryzae can cause disease on rice plants at all developmental stages and can infect different tissues, including leaves, stems, nodes, and panicles [10]. Its aerial conidiophores produce three-celled and teardrop-like conidia that are arranged in a sympodial manner. The attachment of these conidia to the host surface initiates the infection cycle [11]. Conidia adhere to the host with the help of mucilage, a thick and gluey substance stored at the spore tip. They then germinate and generate germ tubes before appressoria differentiation [10][11][12]. The death of three-celled conidia triggers the formation of single-celled appressoria [13]. Then, appressoria become melanized and accumulate glycerol, generating turgor to create pressure for penetration into the host epidermal cells [13]. Upon penetration, it colonizes plants using invasive hyphae that grow intracellularly [14]. Notably, membrane caps, formed on the invading hyphal tips, are used for cell-to-cell movement via manipulating the plasmodesmata [14]. Thus, the membrane interactions between M. oryzae and plant hosts are important for the invasion of hyphae into plant cells [15].
Upon penetration, fungal pathogens can secret effectors to promote virulence and suppress host immunity [16]. In turn, plants have evolved resistance genes (R) that encode R proteins to recognize the presence of effectors and enable effector-triggered immunity (ETI) [16][17]. In M. oryzae, when the first hyphal cell invades, effectors accumulate in the biotrophic interfacial complex (BIC), the site for effector delivery [15][18]. Two types of effectors were defined in M. oryzae based on their localization in plant cells: cytoplasmic effectors entering the host cell cytoplasm via the BIC, and BIC-independent apoplastic effectors being delivered into the apoplast [19][20].
Due to its notable appressoria formation, the secretion of effectors during the invasion, and the effector delivery to the apoplast and cytoplasm of plant cells, M. oryzae has been used as a model plant fungal pathogen to understand pathogen genomics and pathogen–host interactions [11].
2. Genome Sequencing
The first analysis of the M. oryzae genome was published (as M. grisea) in 2005 using the whole-genome shotgun (WGS) approach, with 7× coverage. The genome of M. oryzae 70-15, originally derived from a cross between isolates of rice and weeping lovegrass, consisted of seven chromosomes with an estimated size of 38.8 Mb (GenBank accession number: AACU00000000.3) [21][22]. However, this draft genome sequence was compromised because of significant retrotransposon content. After improvements in verifying unique sequence anchors, extending contigs and scaffolds, and filling the remaining gaps, the genome sequence of the M. oryzae strain 70-15 was refined at the Broad Institute in 2015 with an estimated size of 41 Mb. The high-confidence annotation and gene models were also generated using multiple methods including RNA-seq data and expressed sequence tags (ESTs) alignments, and homologous gene and Basic Local Alignment Search Tool (BLAST) searches [23]. Therefore, this 41 Mb genome sequence of strain 70-15 became the standard reference for M. oryzae, consisting of seven chromosomes with about 51.6% GC content (BioProject Accession number: PRJNA13840).
The initial analysis of the M. oryzae strain 70-15 sequenced in 2005 revealed a family of G-protein-coupled receptors that are specific to M. oryzae and have an expression in infection-related development [22]. Furthermore, they also unveiled three mitogen-activated protein kinase (MAPK) cascades including appressoria development and penetration peg formation, which are crucial in the plant infection process, as well as the relevant genes involved in these signaling cascades. In addition, a significant number of predicted secretion proteins and putative effector proteins, as well as synthases/synthetases involved in secondary metabolic pathways, were identified [22]. Many of these became targets for reverse genetic analysis.
Subsequently, the genome sequences of many other M. oryzae strains became available. For example, Xue et al. (2012) [24] sequenced two M. oryzae field isolates, Y34 (isolates from China) and P131 (from Japan), using Sanger and 454 sequencing. Genome comparisons revealed that the genome size and overall structure were similar among different strains, but more genes that might be involved in host–pathogen interactions were found in the field isolates [24]. They also found different distributions of transposon-like elements among these three strains, indicating genetic variations [24]. More recently, Dr. T. R. Sharma’s group sequenced the whole genome of two isolates from India, an avirulent isolate RML-29 and a highly virulent isolate RP-2421, and identified several isolate-specific genes and potential effectors through pan-genome analysis, which might be involved in fungal pathogenicity and fungal–plant interactions [25][26].
Repetitive transposable elements (TEs) may play important roles in both genome and pathogenic variations in M. oryzae [22][24][27]. They may mediate fungal virulence via the inactivation or deletion of pathogen-associated molecular patterns (PAMPs)-encoding genes and can trigger plant defense responses [28][29]. The M. oryzae genome contains approximately 10% of the repetitive sequences in various isolates [22][23][24][27]. Xue et al. (2012) [24] also uncovered that 23.8% of the TE-disrupted genes were predicted to encode signal peptide sequences, highlighting that this percentage was higher than the average percentage of the whole genome. The single-molecule real-time sequencing also revealed that TEs are involved in chromosomal translocation and secreted proteins (SPs) polymorphisms [27]. In addition, transposon and effector-rich mini-chromosomes were observed in the M. oryzae MoT isolate, which contribute to the field adaptation [30]. These findings suggested a significant role of TEs in defining host specificity and fungal virulence.
3. Transcriptomic and Secretome Analysis
The availability of whole-genome sequencing data has facilitated the transcriptomic and secretome analysis of M. oryzae. The first comprehensive genome-wide transcriptional profiling study of M. oryzae was carried out using microarray analyses. Changes in genome-wide gene expression during the early stages of spore germination and appressoria formation were identified [31]. A microarray analysis of the fungal-invading hyphae at an early stage (36 h post inoculation) uncovered several putative effectors, including fungal biotrophy-associated secreted (BAS) proteins [20]. Moreover, microarray data of invasive growth on rice and barley at a later stage (72 hpi) also revealed multiple genes associated with stress responses and invasive growth [32].
In recent years, high-throughput RNA sequencing has become an efficient approach to provide high-quality transcriptome data. Many transcriptomic analyses were carried out to study host–pathogen interactions. For example, Kawahara et al. (2012) [33] investigated the mixed transcriptome of the pathogen M. oryzae and host O. sativa, revealing 240 upregulated genes encoding potential secreted proteins and many known infection-related genes in M. oryzae [33]. Another study on plant–fungal interactions between the host rice plants and M. oryzae discovered several novel effectors and virulence-related genes, including 98-06 isolate-unique genes IUG6 and IUG9, which were involved in the fungal pathogenicity and located in the BIC [34].
Secretome analysis is another useful approach to identifying crucial genes through studying secreted proteins and their secretion pathways. One M. oryzae secretome study during the early stages of infection identified 53 secreted proteins, including proteins that functioned by modifying fungal lipid and cell walls, detoxifying reactive oxygen species, as well as encouraging fungal metabolism [35]. Liu et al. (2021) [36] also identified several secreted proteins requiring N-glycosylation, which play essential roles in fungal pathogenicity and cell wall integrity [36]. Combining transcriptomic and secretome analyses, these datasets provide an abundant reservoir of candidates for reverse genetic analysis to help understand the molecular mechanisms in M. oryzae–rice interactions.
Mutant analysis is a definitive process used to establish a causal relationship between a gene and a specific biological process. Many methods were applied to generate mutants in M. oryzae, such as T-DNA insertion, RNA interference (RNAi), and homologous recombination (deletion). Among them, gene deletion via homologous recombination has been the most commonly used method in the past few decades. The latest CRISPR technology has also been developed to improve gene-editing efficiency [37][38]. The rest of this research will give an overview of all M. oryzae genes studied to date using such approaches. A Venn diagram of the listed genes is provided for readers’ overview (Figure 1). Over 400 genes have been summarized in this research, and it is clear to see that most of them contribute to both the development and virulence of M. oryzae. A chromosomal map including the significant genes involved in both processes was generated to detect possible cluster patterns and genomic features (Figure 2).
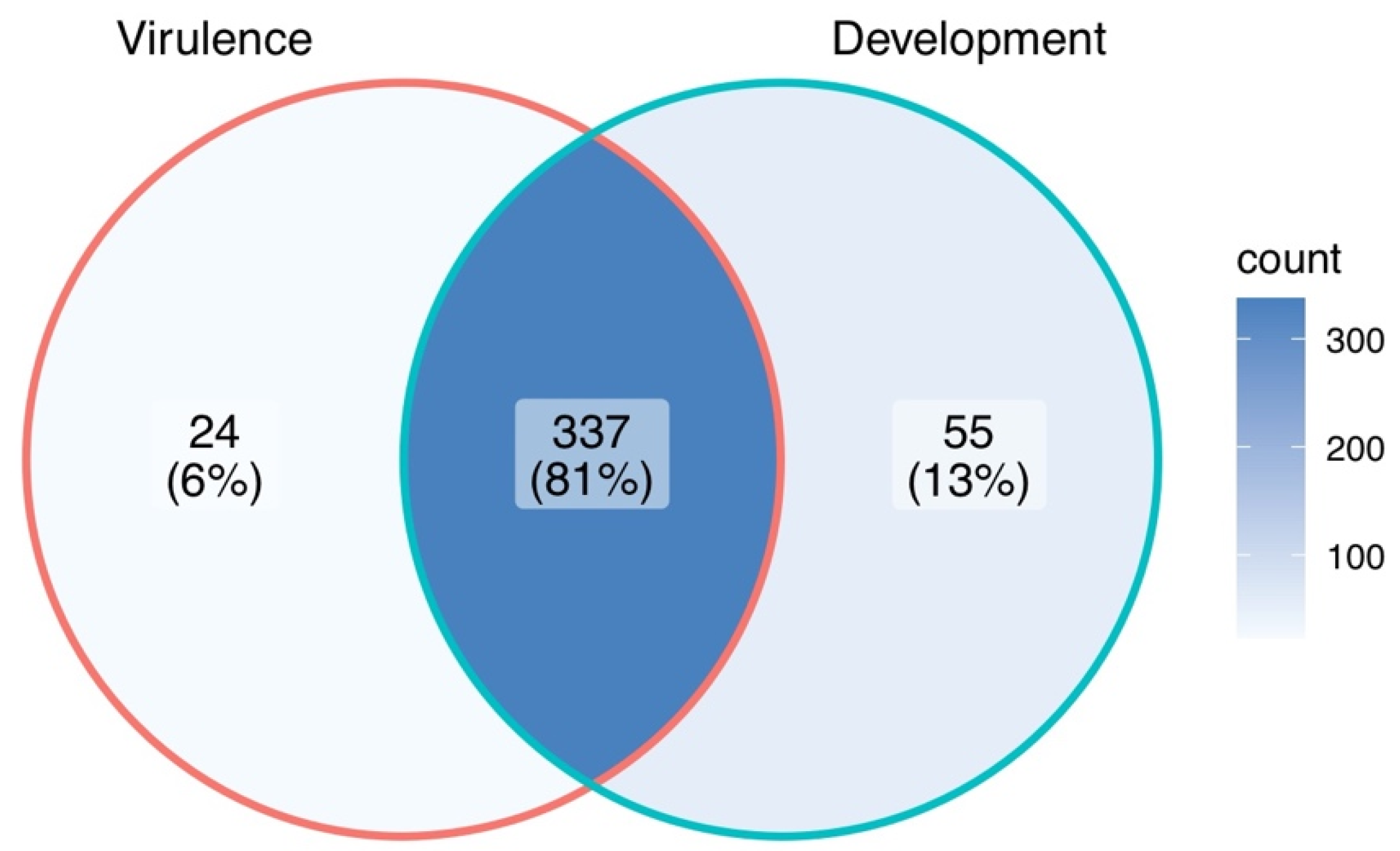
Figure 1. A Venn diagram summary of M. oryzae genes studied with mutant analysis.
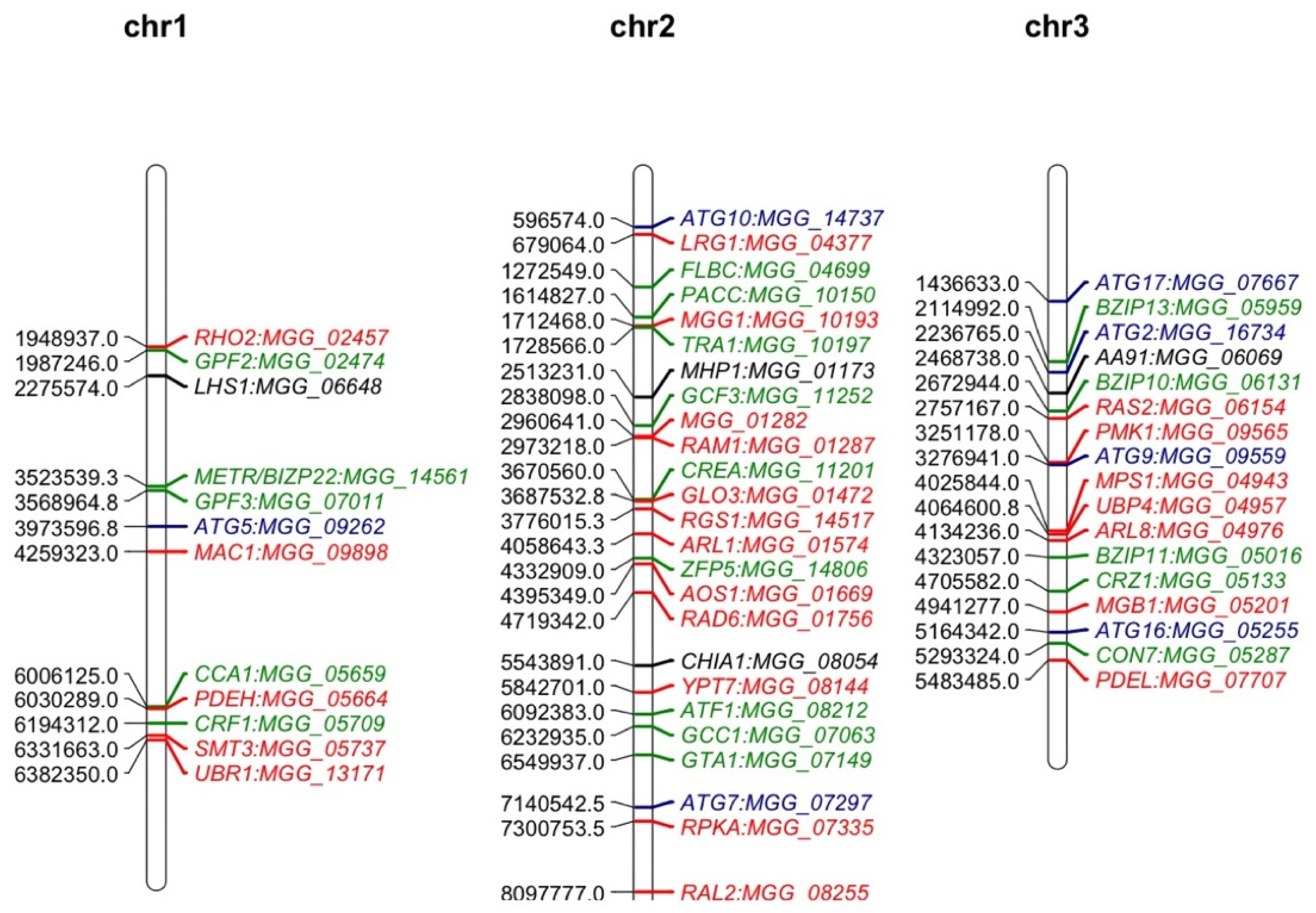
Figure 2. Map positions of important genes studied in the M. oryzae genome through mutant analysis. The numbers on the left of each chromosome represent the locations of these genes. Genes in this map contribute to both development and virulence of M. oryzae. Genes labeled in red are components in different important signaling pathways. Genes labeled in green are transcription factor-encoding genes. Genes labeled in blue are autophagy related while the ones in black are genes encoding effectors. The chromosomal map was drawn using ‘MapChart’ software.
References
- Fairbrothers, D.E. An annotated bibliography of the floristic publications of New Jersey from 1753–1961. Bull. Torrey Bot. Club 1964, 91, 47–56.
- Rossman, A.Y.; Howard, R.J.; Valent, B. Pyricularia grisea, the Correct Name for the Rice Blast Disease Fungus. Mycologia 1990, 82, 509–512.
- Couch, B.C.; Kohn, L.M. A multilocus gene genealogy concordant with host preference indicates segregation of a new species, Magnaporthe oryzae, from M. grisea. Mycologia 2002, 94, 683–693.
- TeBeest, D.O.; Guerber, C.; Ditmore, M. Rice Blast. Available online: https://www.apsnet.org/edcenter/disandpath/fungalasco/pdlessons/Pages/RiceBlast.aspx (accessed on 15 January 2023).
- Meng, Q.; Gupta, R.; Min, C.W.; Kwon, S.W.; Wang, Y.; Je, B.I.; Kim, Y.J.; Jeon, J.S.; Agrawal, G.K.; Rakwal, R.; et al. Proteomics of Rice—Magnaporthe oryzae interaction: What have we learned so far? Front. Plant Sci. 2019, 10, 1383.
- Boddy, L. Pathogens of Autotrophs. In The Fungi; Academic Press: Cambridge, MA, USA, 2016; pp. 245–292.
- Ryan, J.R. Biosecurity and Bioterrorism: Containing and Preventing Biological Threats, 2nd ed.; Butterworth-Heinemann: Oxford, UK, 2016; pp. 1–373.
- Campos-Soriano, L.; Valè, G.; Lupotto, E.; Segundo, B.S. Investigation of rice blast development in susceptible and resistant rice cultivars using a gfp-expressing Magnaporthe oryzae isolate. Plant Pathol. 2013, 62, 1030–1037.
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430.
- Wilson, R.A.; Talbot, N.J. Under pressure: Investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195.
- Ebbole, D.J. Magnaporthe as a Model for Understanding Host-Pathogen Interactions. Annu. Rev. Phytopathol. 2007, 45, 437–456.
- Ribot, C.; Hirsch, J.; Balzergue, S.; Tharreau, D.; Nottéghem, J.-L.; Lebrun, M.-H.; Morel, J.-B. Susceptibility of rice to the blast fungus, Magnaporthe grisea. J. Plant Physiol. 2008, 165, 114–124.
- Veneault-Fourrey, C.; Barooah, M.; Egan, M.; Wakley, G.; Talbot, N.J. Autophagic Fungal Cell Death Is Necessary for Infection by the Rice Blast Fungus. Science 2006, 312, 580–583.
- Kankanala, P.; Czymmek, K.; Valent, B. Roles for Rice Membrane Dynamics and Plasmodesmata during Biotrophic Invasion by the Blast Fungus. Plant Cell 2007, 19, 706–724.
- Valent, B.; Khang, C.H. Recent advances in rice blast effector research. Curr. Opin. Plant Biol. 2010, 13, 434–441.
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329.
- Giraldo, M.; Valent, B. Filamentous plant pathogen effectors in action. Nat. Rev. Microbiol. 2013, 11, 800–814.
- Khang, C.H.; Berruyer, R.; Giraldo, M.C.; Kankanala, P.; Park, S.-Y.; Czymmek, K.; Kang, S.; Valent, B. Translocation of Magnaporthe oryzae Effectors into Rice Cells and Their Subsequent Cell-to-Cell Movement. Plant Cell 2010, 22, 1388–1403.
- Giraldo, M.C.; Dagdas, Y.F.; Gupta, Y.K.; Mentlak, T.A.; Yi, M.; Martinez-Rocha, A.L.; Saitoh, H.; Terauchi, R.; Talbot, N.J.; Valent, B. Two distinct secretion systems facilitate tissue invasion by the rice blast fungus Magnaporthe oryzae. Nat. Commun. 2013, 4, 1996.
- Mosquera, G.; Giraldo, M.C.; Khang, C.H.; Coughlan, S.; Valent, B. Interaction Transcriptome Analysis Identifies Magnaporthe oryzae BAS1-4 as Biotrophy-Associated Secreted Proteins in Rice Blast Disease. Plant Cell 2009, 21, 1273–1290.
- Chao, C.-C.T.; Ellingboe, A.H. Selection for mating competence in Magnaporthe grisea pathogenic to rice. Can. J. Bot. 1991, 69, 2130–2134.
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.R.; Pan, H.; et al. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 2005, 434, 980–986.
- Okagaki, L.H.; Nunes, C.C.; Sailsbery, J.; Clay, B.; Brown, D.; John, T.; Oh, Y.; Young, N.; Fitzgerald, M.; Haas, B.J.; et al. Genome Sequences of Three Phytopathogenic Species of the Magnaporthaceae Family of Fungi. G3 Genes Genomes Genet. 2015, 5, 2539–2545.
- Xue, M.; Yang, J.; Li, Z.; Hu, S.; Yao, N.; Dean, R.A.; Zhao, W.; Shen, M.; Zhang, H.; Li, C.; et al. Comparative Analysis of the Genomes of Two Field Isolates of the Rice Blast Fungus Magnaporthe oryzae. PLoS Genet. 2012, 8, e1002869.
- Ray, S.; Singh, P.K.; Gupta, D.K.; Mahato, A.K.; Sarkar, C.; Rathour, R.; Singh, N.K.; Sharma, T.R. Analysis of Magnaporthe oryzae Genome Reveals a Fungal Effector, Which Is Able to Induce Resistance Response in Transgenic Rice Line Containing Resistance Gene, Pi54. Front. Plant Sci. 2016, 7, 1140.
- Singh, P.K.; Mahato, A.K.; Jain, P.; Rathour, R.; Sharma, V.; Sharma, T.R. Comparative Genomics Reveals the High Copy Number Variation of a Retro Transposon in Different Magnaporthe Isolates. Front. Microbiol. 2019, 10, 966.
- Bao, J.; Chen, M.; Zhong, Z.; Tang, W.; Lin, L.; Zhang, X.; Jiang, H.; Zhang, D.; Miao, C.; Tang, H.; et al. PacBio Sequencing Reveals Transposable Elements as a Key Contributor to Genomic Plasticity and Virulence Variation in Magnaporthe oryzae. Mol. Plant 2017, 10, 1465–1468.
- Farman, M.L.; Eto, Y.; Nakao, T.; Tosa, Y.; Nakayashiki, H.; Mayama, S.; Leong, S.A. Analysis of the Structure of the AVR1-CO39 Avirulence Locus in Virulent Rice-Infecting Isolates of Magnaporthe grisea. Mol. Plant-Microbe Interact. 2002, 15, 6–16.
- Kang, S.; Lebrun, M.H.; Farrall, L.; Valent, B. Gain of Virulence Caused by Insertion of a Pot3 Transposon in a Magnaporthe grisea Avirulence Gene. Mol. Plant-Microbe Interact. 2001, 14, 671–674.
- Peng, Z.; Oliveira-Garcia, E.; Lin, G.; Hu, Y.; Dalby, M.; Migeon, P.; Tang, H.; Farman, M.; Cook, D.; White, F.F.; et al. Effector gene reshuffling involves dispensable mini-chromosomes in the wheat blast fungus. PLoS Genet. 2019, 15, e1008272.
- Oh, Y.; Donofrio, N.; Pan, H.; Coughlan, S.; Brown, D.E.; Meng, S.; Mitchell, T.; Dean, R.A. Transcriptome analysis reveals new insight into appressorium formation and function in the rice blast fungus Magnaporthe oryzae. Genome Biol. 2008, 9, R85.
- Mathioni, S.M.; Beló, A.; Rizzo, C.J.; Dean, R.A.; Donofrio, N.M. Transcriptome profiling of the rice blast fungus during invasive plant infection and in vitro stresses. BMC Genom. 2011, 12, 49.
- Kawahara, Y.; Oono, Y.; Kanamori, H.; Matsumoto, T.; Itoh, T.; Minami, E. Simultaneous RNA-Seq Analysis of a Mixed Transcriptome of Rice and Blast Fungus Interaction. PLoS ONE 2012, 7, e49423.
- Dong, Y.; Li, Y.; Zhao, M.; Jing, M.; Liu, X.; Liu, M.; Guo, X.; Zhang, X.; Chen, Y.; Liu, Y.; et al. Global Genome and Transcriptome Analyses of Magnaporthe oryzae Epidemic Isolate 98-06 Uncover Novel Effectors and Pathogenicity-Related Genes, Revealing Gene Gain and Lose Dynamics in Genome Evolution. PLoS Pathog. 2015, 11, e1004801.
- Jung, Y.-H.; Jeong, S.-H.; Kim, S.H.; Singh, R.; Lee, J.-E.; Cho, Y.-S.; Agrawal, G.K.; Rakwal, R.; Jwa, N.-S. Secretome analysis of Magnaporthe oryzae using in vitro systems. Proteomics 2012, 12, 878–900.
- Liu, N.; Qi, L.; Huang, M.; Chen, D.; Yin, C.; Zhang, Y.; Wang, X.; Yuan, G.; Wang, R.-J.; Yang, J.; et al. Comparative Secretome Analysis of Magnaporthe oryzae Identified Proteins Involved in Virulence and Cell Wall Integrity. Genom. Proteom. Bioinform. 2022, 20, 728–746.
- Foster, A.J.; Martin-Urdiroz, M.; Yan, X.; Wright, H.S.; Soanes, D.M.; Talbot, N.J. CRISPR-Cas9 ribonucleoprotein-mediated co-editing and counterselection in the rice blast fungus. Sci. Rep. 2018, 8, 1–12.
- Huang, J.; Rowe, D.; Subedi, P.; Zhang, W.; Suelter, T.; Valent, B.; Cook, D.E. CRISPR-Cas12a induced DNA double-strand breaks are repaired by multiple pathways with different mutation profiles in Magnaporthe oryzae. Nat. Commun. 2022, 13, 7168.
More
Information
Subjects:
Pathology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
727
Revisions:
2 times
(View History)
Update Date:
18 Apr 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No