Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Zoya Salahuddin | -- | 1946 | 2023-04-04 10:05:42 | | | |
| 2 | Dean Liu | -47 word(s) | 1899 | 2023-04-06 03:14:21 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Kafoud, A.; Salahuddin, Z.; Ibrahim, R.S.; Al-Janahi, R.; Mazurakova, A.; Kubatka, P.; Büsselberg, D. Mechanisms of Polyphenols as Treatment Against Neuroblastoma. Encyclopedia. Available online: https://encyclopedia.pub/entry/42765 (accessed on 28 July 2026).
Kafoud A, Salahuddin Z, Ibrahim RS, Al-Janahi R, Mazurakova A, Kubatka P, et al. Mechanisms of Polyphenols as Treatment Against Neuroblastoma. Encyclopedia. Available at: https://encyclopedia.pub/entry/42765. Accessed July 28, 2026.
Kafoud, Aisha, Zoya Salahuddin, Raghad Sabaawi Ibrahim, Reem Al-Janahi, Alena Mazurakova, Peter Kubatka, Dietrich Büsselberg. "Mechanisms of Polyphenols as Treatment Against Neuroblastoma" Encyclopedia, https://encyclopedia.pub/entry/42765 (accessed July 28, 2026).
Kafoud, A., Salahuddin, Z., Ibrahim, R.S., Al-Janahi, R., Mazurakova, A., Kubatka, P., & Büsselberg, D. (2023, April 04). Mechanisms of Polyphenols as Treatment Against Neuroblastoma. In Encyclopedia. https://encyclopedia.pub/entry/42765
Kafoud, Aisha, et al. "Mechanisms of Polyphenols as Treatment Against Neuroblastoma." Encyclopedia. Web. 04 April, 2023.
Copy Citation
Neuroblastoma (NB) is an extracranial tumor of the peripheral nervous system arising from neural crest cells. It is the most common malignancy in infants and the most common extracranial solid tumor in children. The treatment for high-risk NB involves chemotherapy and surgical resection followed by high-dose chemotherapy with autologous stem-cell rescue and radiation treatment. However, those with high-risk NB are susceptible to relapse and the long-term side effects of standard chemotherapy. Polyphenols, including the sub-class of flavonoids, contain more than one aromatic ring with hydroxyl groups.
neuroblastoma
cancer
flavonoids
polyphenols
1. Calpain-Dependent Apoptotic Pathway
The release of calcium (Ca2+) from the endoplasmic reticulum (ER) leads to the activation of calpains. Calpain is a Ca2+-activated endo-protease involved in apoptotic mechanisms [1]. Exposure to the compounds listed in Table 1 activates proteolytic pathways involving calpain, leading to NB-cell apoptosis. Specifically, apigenin, epigallocatechin (EGC), epigallocatechin gallate (EGCG), and genistein trigger ER stress, thus increasing intracellular free Ca2+ and causing calpain activation at the ER membrane [2][3], resulting in the degradation of cytoskeletal proteins and the destabilization of cellular integrity in SH-SY5Y cells. Caspase release is also stimulated, with caspase-3 activating caspase-activated DNase (CAD), contributing to DNA fragmentation [4][5]. Caspase-12 is also activated when apigenin and genistein are applied, reinforcing their apoptotic effects [3]. Several studies concerning other cancer types reinforced the calpain–caspase apoptotic pathway, illustrated in Figure 1 [1][2][3][5][6].
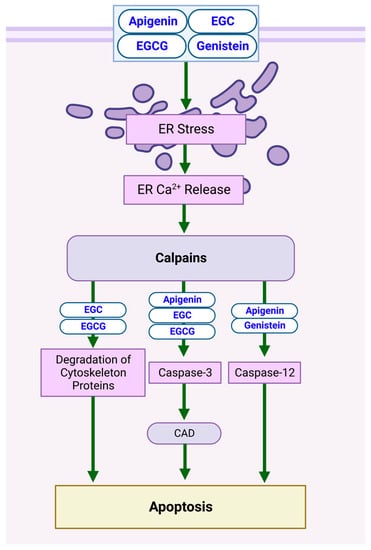
Figure 1. Schematic diagram demonstrating the apoptotic effects of flavonoids on NB cell lines via a calpain-dependent pathway. The induction of ER stress induces Ca2+ release at the ER membrane, triggering caspase and CAD release and degradation of cytoskeletal proteins. Specific biomarkers can be viewed in Table 1. Created with BioRender.com.
Table 1. Results of in vitro studies of polyphenols’ apoptotic effects on NB cell lines via a calpain-dependent pathway *.
| Compound | Cell Line | Incubation Period | Concentration(s) | Biomarker Changes | Reference |
|---|---|---|---|---|---|
| Flavonoids | |||||
| Apigenin | SH-SY5Y | 24 h | 50 µM | ↑ Intracellular free [Ca2+] ↑ Calpain activation ↑ Caspase-12, -3 ↑ CAD |
[3] |
| EGC | SH-SY5Y | 24 h | 50 µM | ↑ Intracellular free [Ca2+] ↑ Calpain activation ↑ Cytoskeletal protein degradation ↑ Caspase-3 ↑ CAD |
[3] |
| EGCG | SH-SY5Y | 24 h | 50 µM | ↑ Intracellular free [Ca2+] ↑ Calpain activation ↑ Cytoskeletal protein degradation ↑ Caspase-3 ↑ CAD |
[3] |
| Genistein | SH-SY5Y | 24 h | 100 µM | ↑ Intracellular free [Ca2+] ↑ Calpain activation ↑ Caspase-12 |
[3] |
* ↑ denotes increase of biomarker, while ↓ denotes decrease.
2. Anti-Proliferative Pathways
In vitro studies on the anti-cancer effects of flavonoids and non-flavonoid polyphenols in NB cell lines reveal remarkable anti-proliferative effects, as demonstrated in Figure 2 and Table 2. Quercetin exhibits anti-proliferative effects by increasing p27 mRNA expression, inhibiting the formation and activity of the cyclin–cyclin-dependent kinase (cyclin/CDK) complex, disrupting the cell cycle in NB. Quercetin also reduces B-cell lymphoma-w (Bcl-w) mRNA expression, which decreases tumor-gene expression and induces apoptosis in NB cells [7]. Quercetin can further inhibit NB-cell growth by blocking voltage-gated potassium (K+) channel activity [8]. Other flavonoids, such as apigenin and 2-(cis-1,2-dihydroxy-4-oxo-cyclohex-5-enyl)-5,7-dihydroxy-chromone (DEDC), increase p53 and p21 mRNA expression while decreasing cyclin-B1 expression in NB [9][10]. Didymin decreases the proliferation of NB cells via the downregulation of the phosphoinositide 3-kinase (PI3K) and Akt pathways, accompanied by reduced vimentin levels, indicating a decrease in cell motility. Furthermore, proto-oncogene N-Myc transcription was inhibited by didymin. Increased Raf-1 kinase inhibitor protein (RKIP) levels inhibit the mitogen-activated protein kinase (MAPK) pathway, also decreasing proliferation [11]. Isoliquiritigenin inhibits cell motility and increases the activation of extracellular regulated kinase 1/2 (pERK1/2), which inhibits NB-cell migration and proliferation while arresting the cell cycle in the S phase [12]. Isoliquiritigenin and rutin both enhance G2/M-phase arrest in NB [12][13].
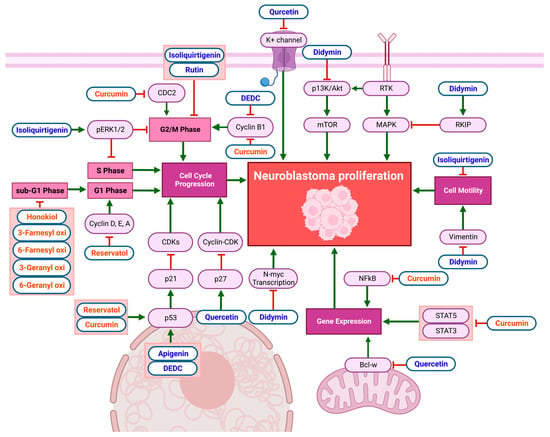
Figure 2. Schematic diagram demonstrating the anti-proliferative effects of flavonoids (blue) and non-flavonoid polyphenols (orange) on NB cell lines. The inhibition of cell-cycle progression, cell motility, and gene expression limits NB-cell proliferation. Compounds caused cell-cycle arrest in the S phase, sub-G1 phase, G1 phase, and G2/M phase. Specific biomarkers can be viewed in Table 2. Created with BioRender.com.
Non-flavonoid polyphenols affect multiple cell lines through multiple pathways. Curcumin decreases CDC2 and cyclin-B1, resulting in NB-cell-cycle arrest in the G2/M phase [14]. Furthermore, it reduces NF-κB activator protein (AP-1) and STAT3 and STAT5 activation, suppressing gene transcription [15]. Honokiol inhibits NB-cell-cycle progression at the sub-G1 phase [16]. Resveratrol reduced Cyclin D1 levels in NB cells, causing cell-cycle arrest in the S phase [17]. Similarly, the treatment of NB cells with resveratrol resulted in a significant drop in pAkt, Cyclin D, E, A, and CDK2 levels and increased p53 and NF-κB, resulting in cell-cycle arrest in the S phase [18]. Resveratrol causes p21 levels to rise, which inhibits CDK levels and causes cell-cycle arrest in the G1, G2/M, and S phases in NB cell lines [19]. Prenyl hydroxy coumarin derivatives also have notable anti-proliferative effects on NB cell lines, inducing cell-cycle arrest in the sub-G1 phase, with no effect on normal lymphocytic cells [20].
In addition to isolated flavonoids and non-flavonoid polyphenolic compounds, recent research supports the potent anti-cancer effects of whole plants or plant extracts characterized by numerous phytochemicals acting synergistically or additively [21][22][23][24]. For example, a recent study by Morandi et al. (2021) demonstrated the capacity of olive-leaf extract (rich in phenolic compounds) to inhibit the proliferation of NB cells through cell arrest in the G0/G1 phase and the accumulation of cells in the sub-G0 phase, accompanied by the induction of apoptosis [25]. Numerous other recent studies highlight the anti-cancer potential of plant extracts; for example, the fruit extract of Kigelia Africana, a plant rich in flavonoids that are used in traditional African medicine, inhibited proliferation and other mechanisms associated with carcinogenesis in NB cells [26]. Interestingly, research results obtained by Roomi et al. (2013) suggested the therapeutic potential of a nutrient mixture of lysine, proline, ascorbic acid, and green-tea extracts for NB management through the inhibition of tumor growth and proliferation and the induction of apoptosis in neuroblastoma models in vitro and in vivo [27]. Indeed, green tea is rich in numerous phytochemicals, mainly catechins. Green-tea catechins, including ECG, EGCG, and EGC, are phytochemicals with strong anti-cancer effects [28][29].
Table 2. Results of in vitro studies of polyphenols’ anti-proliferative effects on NB cell lines *.
| Compound | Cell Line | Incubation Period | Concentration(s) | Biomarker Changes and Effects | Reference | |
|---|---|---|---|---|---|---|
| Flavonoids | ||||||
| Apigenin | NUB-7 and LAN-5 | 24 h | 10, 50, 100, 150, 200 µM IC50: 35 μM in NUB-7 IC50: 22 μM in LAN-5 |
↑ p53 ↑ p21WAF−1/CIP−1 |
↓ Proliferation | [10] |
| DEDC | SH-SY5Y | 24 h | 7.5 µg/mL | ↑ p53 mRNA ↑ p21 mRNA ↓ Cyclin-B1 |
[9] | |
| Didymin | CHLA-90 and SK-N-BE2 (p53-mutant) l + SMS-KCNR and LAN-5 (p53 wild-type) | 24 h | 50 μmol/L | ↓ P13K ↓ Akt |
↓ Proliferation | [11] |
| ↓ Vimentin | ↓ Motility of tumor cells | |||||
| ↓ N-Myc transcription | ||||||
| ↑ RKIP | ↓ MAPK pathway ↓ Proliferation |
|||||
| Isoliquiritigenin | SH-SY5Y | 24 h | 10–100 µM IC50: 25.4 µM |
↑ pERK1/2 | ↓ Cell migration ↓ Proliferation ↑ S + G2/M-phase arrest |
[12] |
| Rutin | LAN-5 | 24 h | 0, 25, 50, 100 μM | ↑ G2/M-phase arrest | [13] | |
| Quercetin | Neuro2a (mouse cell line) | 24 h | 10, 20, 40, 80, 120 μM IC50: 40 µM |
↑ p27 | ↓ Cyclin–CDK complex binding | [7] |
| ↓ Bcl-w | ↓ Tumor-cell-gene expression | |||||
| Quercetin | Neuroblastoma X glioma NG 108-15 cells (mouse cell line) | 48 h | 10 µM, 20 µM IC50: 10 µM |
↓ K+-channel activity | ↓ Cell growth | [8] |
| Non-Flavonoid Polyphenols | ||||||
| Curcumin | SK-N-SH | 24 h | 8, 16, 32 µM | ↓ CDC2 ↓ Cyclin B1 |
↑ G2/M-phase arrest | [14] |
| Curcumin | GI-L-IN, HTLA-230, SH-SY5Y, LAN5, SK-NBE2c, and IMR-32 | 18–72 h | 0.1–25 µM | ↓ NFκβ activator protein (AP-1) ↓ STAT3, STAT5 activation |
↓ Cell growth | [15] |
| Curcumin | NUB-7, LAN-5, IMR-32 and SK-N-BE(2) | 2–8 days | 0–100 µM * * Significantly inhibited proliferation in the range of 5–10 µM |
↑ p53 translocation from cytoplasm to nucleus ↑ p21WAF−1/CIP−1 |
↑ G1-, G2/M-, and S-phase arrest | [19] |
| Honokiol | Neuro-2a (mouse cell line) and NB41A3 | 72 h | 2.5, 5, 10, 20, 30, 40, 50, 60, 80, 100 µM LC50: 63.3 µM |
↑ Sub-G1-phase arrest | [16] | |
| Prenyl hydroxy-coumarins | Neuro-2a (mouse cell line) | 24, 48, 72 h | 6.25–200 µg/mL | ↑ Sub-G1-phase arrest | [20] | |
| Resveratrol | B103 (rat cell line) | 48 h | 5–20 µM IC50: 17.86 µM |
↓ Cyclin D1 | ↑ G1-phase arrest | [17] |
| Resveratrol | B65 (rat dopaminergic cell line) | 24 h | 25, 50, 100 µM | ↓ pAkt ↓ Cyclin D, E, A ↓ CDK2 ↑ p53 ↑ NFκβ |
↑ S-phase arrest | [18] |
| Resveratrol | NUB-7, LAN-5, IMR-32 and SK-N-BE(2) | 2–8 days | 25–160 µM | ↑ p53 translocation from cytoplasm to nucleus ↑ p21WAF−1/CIP−1 |
↑ G1-, G2/M-, and S-phase arrest | [30] |
* ↑ denotes increase of biomarker, while ↓ denotes decrease.
3. Mitochondrial and ER-Stress-Related Apoptotic Pathways
Results of several studies have also identified Mitochondrial and ER-Stress-Related Apoptotic Pathways, as demonstrated in Figure 3 and Table 3.
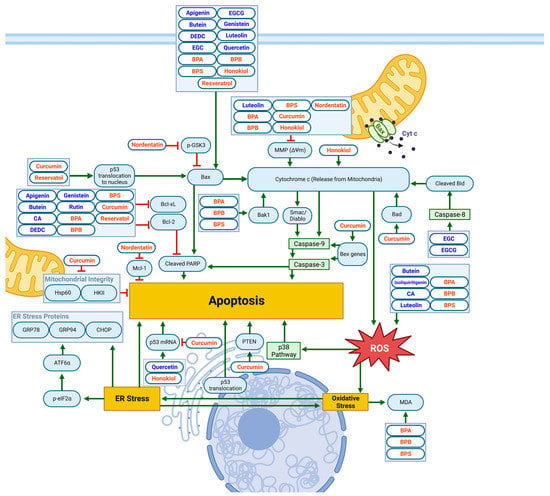
Figure 3. Schematic diagram demonstrating the apoptotic effects of flavonoids (blue) and non-flavonoid polyphenols (orange) on NB cell lines via mitochondrial or ER/oxidative-stress-related pathways. Elevated Bax/Bcl-2 ratio, increased PARP cleavage, loss of MMP and cytochrome C release, and ROS generation all contribute to apoptotic cell death. Specific biomarkers can be viewed in Table 3. Created with BioRender.com.
Table 3. Results of in vitro studies of polyphenols’ apoptotic effects on NB cell lines via receptor-mediated pathways *.
| Compound | Cell Line | Incubation Period | Concentration(s) | Biomarker Changes | Reference |
|---|---|---|---|---|---|
| Flavonoids | |||||
| DEDC | SH-SY5Y | 24 h | 7.5 µg/mL | ↓ Phosphor-STAT3 expression (ROS mediated) | [3] |
| Genistein | SK-N-DZ | 24 h | 10 µM | ↑ TNF-α ↑ FasL ↑ TRADD ↑ FADD |
[3] |
| EGC | SH-SY5Y | 24 h | 50 µM | ↑ Caspase-8 activation ↑ Proteolytic cleavage of Bid to tBid ↑ Bax oligomerization |
[3] |
| EGCG | SH-SY5Y | 24 h | 100 µM | ↑ Caspase-8 activation ↑ Proteolytic cleavage of Bid to tBid ↑ Bax oligomerization |
[3] |
| Rutin | LAN-5 | 24 h | 25, 50, 100 μM | ↑ TNF-α secretion | [13] |
| Non-Flavonoid Polyphenols | |||||
| Curcumin | LAN-5 | 3, 5, 24 h | 5, 10, 15, 20 µM | ↑ Bad ↑ PTEN ↑ ROS |
[31] |
| Honokiol | Neuro-2a (mouse cell line) | 30, 60, 120 µM | 24, 48, 72 h | ↑ RIP3 ↑ ROS |
[32] |
* ↑ denotes increase of biomarker, while ↓ denotes decrease.
The flavonoid DEDC decreases the expression of phospho-STAT3 in a ROS-mediated manner in NB cells [9]. The downregulation of phospho-STAT3 leads to the upregulation of p53 and p21, downregulating Cyclin B1, resulting in cell-cycle arrest and apoptosis in the G2/M phase [9].
The flavonoids genistein and rutin have similar mechanisms, leading to apoptotic effects. Rutin increases NB’s necrosis factor α (TNF-α) secretion [13]. Similarly, genistein increased TNF-α and Fas ligand (FasL), TRADD, and FADD in NB [33]. The upregulation of TNF and FasL lead to upregulated TNF receptor type 1-associated DEATH domain protein (TRADD), and Fas-associated DEATH domain protein (FADD) activates caspase-8, which cleaves Bid to tBid. Increased levels of tBid encourage conformational changes in Bax, forming oligomer channels, which promote cytochrome c release. The release of tBid activates caspase-3, leading to nuclear-DNA fragmentation and apoptosis. In addition, both studies found a decrease in antiapoptotic Bcl-2, an increase in pro-apoptotic Bax and, therefore, an overall increase in the Bax:Bcl-2 ratio, which triggers the mitochondrial pathway of apoptosis by releasing cytochrome c in NB. This mechanism is also present when EGC and EGCG increase the activation of caspase-8 and the proteolytic cleavage of Bid to tBid, and Bax oligomerization leads to apoptosis [3].
Similarly, EGC and/or EGCG showed significant effects through the induction of both mitochondria-mediated (as evidenced by increased Bax and decreased Bcl-2) and receptor-mediated apoptotic pathways (demonstrated via the activation of caspase-8 and the cleavage of Bid into tBid) in two NB-cell lines. However, the effects of the EGC and EGCG on apoptosis were accompanied by decreased oncogenic miRNAs and increased tumor-suppressor miRNAs. Furthermore, the authors concluded that the overexpression of tumor-suppressor miRNA-7-1 increases the efficacy of EGCG in apoptosis induction, suggesting the potential of this combination therapy in NB [29].
The non-flavonoid polyphenol curcumin decreases heat shock protein-60 (Hsp60) and hexokinase II (HK-II) levels and increases Bcl-2 associated agonist of cell death (Bad), phosphatase and tensin homolog (PTEN), and ROS in NB cells [31]. The mitochondrial protein Bad increases mitochondrial membrane permeability, promoting cytochrome c release and apoptosis via an intrinsic apoptotic pathway [31]. Phosphatase and tensin homolog is associated with the (PI3K)/Akt pathway—a “highly oncogenic pro-survival” signaling pathway. The PTEN antagonizes the (PI3K)/Akt pathway, leading to ROS-mediated apoptosis [31]. Honokiol, another non-flavonoid polyphenol, demonstrated upregulation of ROS, resulting in increased receptor-interacting protein kinase 3 (RIP3), a critical regulator of programmed necrosis [32].
Resveratrol showed its capacity to increase caspase-3, resulting in an inhibited cell cycle in NB cells [18]. Activated caspase-3 plays a role in cell maintenance and survival through the cleaving of multiple structural and regulatory proteins. Additionally, activated caspase-3 contributes to mitochondrial apoptotic processes, such as releasing cytochrome c and Bax translocation [34].
References
- Momeni, H.R. Role of Calpain in Apoptosis. Cell J. (Yakhteh) 2011, 13, 65.
- Martinez, J.A.; Zhang, Z.; Svetlov, S.I.; Hayes, R.L.; Wang, K.K.; Larner, S.F. Calpain and Caspase Processing of Caspase-12 Contribute to the ER Stress-Induced Cell Death Pathway in Differentiated PC12 Cells. Apoptosis 2010, 15, 1480–1493.
- Das, A.; Banik, N.L.; Ray, S.K. Mechanism of Apoptosis with the Involvement of Calpain and Caspase Cascades in Human Malignant Neuroblastoma SH-SY5Y Cells Exposed to Flavonoids. Int. J. Cancer 2006, 119, 2575–2585.
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2018, 11, 28.
- Ray, S.K.; Fidan, M.; Nowak, M.W.; Wilford, G.G.; Hogan, E.L.; Banik, N.L. Oxidative Stress and Ca2+ Influx Upregulate Calpain and Induce Apoptosis in PC12 Cells. Brain Res. 2000, 852, 326–334.
- Sergeev, I.N. Genistein Induces Ca2+-Mediated, Calpain/Caspase-12-Dependent Apoptosis in Breast Cancer Cells. Biochem. Biophys. Res. Commun. 2004, 321, 462–467.
- Sugantha Priya, E.; Selvakumar, K.; Bavithra, S.; Elumalai, P.; Arunkumar, R.; Raja Singh, P.; Brindha Mercy, A.; Arunakaran, J. Anti-Cancer Activity of Quercetin in Neuroblastoma: An in Vitro Approach. Neurol. Sci. 2014, 35, 163–170.
- Rouzaire-Dubois, B.; Gérard, V.; Dubois, J.M. Involvement of K+ Channels in the Quercetin-Induced Inhibition of Neuroblastoma Cell Growth. Pflügers Archiv. 1993, 423, 202–205.
- Liu, H.; Jiang, C.; Xiong, C.; Ruan, J. DEDC, a New Flavonoid Induces Apoptosis via a ROS-Dependent Mechanism in Human Neuroblastoma SH-SY5Y Cells. Toxicol. Vitr. 2012, 26, 16–23.
- Torkin, R.; Lavoie, J.F.; Kaplan, D.R.; Yeger, H. Induction of Caspase-Dependent, P53-Mediated Apoptosis by Apigenin in Human Neuroblastoma. Mol. Cancer 2005, 4, 1–11.
- Singhal, J.; Nagaprashantha, L.D.; Vatsyayan, R.; Ashutosh; Awasthi, S.; Singhal, S.S. Didymin Induces Apoptosis by Inhibiting N-Myc and Upregulating RKIP in Neuroblastoma. Cancer Prev. Res. 2012, 5, 473–483.
- Escobar, S.J.d.M.; Fong, G.M.; Winnischofer, S.M.B.; Simone, M.; Munoz, L.; Dennis, J.M.; Rocha, M.E.M.; Witting, P.K. Anti-Proliferative and Cytotoxic Activities of the Flavonoid Isoliquiritigenin in the Human Neuroblastoma Cell Line SH-SY5Y. Chem. Biol. Interact. 2019, 299, 77–87.
- Chen, H.; Miao, Q.; Geng, M.; Liu, J.; Hu, Y.; Tian, L.; Pan, J.; Yang, Y. Anti-Tumor Effect of Rutin on Human Neuroblastoma Cell Lines through Inducing G2/M Cell Cycle Arrest and Promoting Apoptosis. Sci. World J. 2013, 2013, 269165.
- Ye, Z.; Chen, D.; Zheng, R.; Chen, H.; Xu, T.; Wang, C.; Zhu, S.; Gao, X.; Zhang, J.; Li, D.; et al. Curcumin Induced G2/M Cycle Arrest in SK-N-SH Neuroblastoma Cells through the ROS-Mediated P53 Signaling Pathway. J. Food Biochem. 2021, 45, e13888.
- Pisano, M.; Pagnan, G.; Dettori, M.A.; Cossu, S.; Caffa, I.; Sassu, I.; Emionite, L.; Fabbri, D.; Cilli, M.; Pastorino, F.; et al. Enhanced Anti-Tumor Activity of a New Curcumin-Related Compound against Melanoma and Neuroblastoma Cells. Mol. Cancer 2010, 9, 1–12.
- Lin, J.W.; Chen, J.T.; Hong, C.Y.; Lin, Y.L.; Wang, K.T.; Yao, C.J.; Lai, G.M.; Chen, R.M. Honokiol Traverses the Blood-Brain Barrier and Induces Apoptosis of Neuroblastoma Cells via an Intrinsic Bax-Mitochondrion-Cytochrome c-Caspase Protease Pathway. Neuro Oncol. 2012, 14, 302–314.
- Rahman, M.A.; Kim, N.H.; Kim, S.H.; Oh, S.M.; Huh, S.O. Anti-proliferative and Cytotoxic Effects of Resveratrol in Mitochondria-Mediated Apoptosis in Rat B103 Neuroblastoma Cells. Korean J. Physiol. Pharm. 2012, 16, 321–326.
- Pizarro, J.G.; Verdaguer, E.; Ancrenaz, V.; Junyent, F.; Sureda, F.; Pallàs, M.; Folch, J.; Camins, A. Resveratrol Inhibits Proliferation and Promotes Apoptosis of Neuroblastoma Cells: Role of Sirtuin 1. Neurochem. Res. 2011, 36, 187–194.
- Liontas, A.; Yeger, H. Curcumin and Resveratrol Induce Apoptosis and Nuclear Translocation and Activation of P53 in Human Neuroblastoma. Anticancer Res. 2004, 24, 987.
- Sargolzaei, J.; Sadeghian, H.; Golahmadi, S.; Soukhtanloo, M. Cytotoxic Effects of Hydroxy Coumarin Derivations to Neuroblastoma N2a Cells. Iran. J. Pharm. Sci. 2020, 16, 95–106.
- Kubatka, P.; Kapinová, A.; Kello, M.; Kruzliak, P.; Kajo, K.; Výbohová, D.; Mahmood, S.; Murin, R.; Viera, T.; Mojžiš, J.; et al. Fruit Peel Polyphenols Demonstrate Substantial Anti-Tumour Effects in the Model of Breast Cancer. Eur. J. Nutr. 2016, 55, 955–965.
- Kapinova, A.; Stefanicka, P.; Kubatka, P.; Zubor, P.; Uramova, S.; Kello, M.; Mojzis, J.; Blahutova, D.; Qaradakhi, T.; Zulli, A.; et al. Are Plant-Based Functional Foods Better Choice against Cancer than Single Phytochemicals? A Critical Review of Current Breast Cancer Research. Biomed. Pharmacother. 2017, 96, 1465–1477.
- Hosseini, A.; Ghorbani, A. Cancer Therapy with Phytochemicals: Evidence from Clinical Studies. Avicenna J. Phytomed. 2015, 5, 84.
- Kubatka, P.; Kello, M.; Kajo, K.; Samec, M.; Liskova, A.; Jasek, K.; Koklesova, L.; Kuruc, T.; Adamkov, M.; Smejkal, K.; et al. Rhus Coriaria L. (Sumac) Demonstrates Oncostatic Activity in the Therapeutic and Preventive Model of Breast Carcinoma. Int. J. Mol. Sci. 2021, 22, 183.
- Morandi, F.; Bensa, V.; Calarco, E.; Pastorino, F.; Perri, P.; Corrias, M.V.; Ponzoni, M.; Brignole, C. The Olive Leaves Extract Has Anti-Tumor Effects against Neuroblastoma through Inhibition of Cell Proliferation and Induction of Apoptosis. Nutrients 2021, 13, 2178.
- Ahrens, S.; Appl, B.; Trochimiuk, M.; Dücker, C.; Feixas Serra, G.; Oliver Grau, A.; Reinshagen, K.; Pagerols Raluy, L. Kigelia Africana Inhibits Proliferation and Induces Cell Death in Stage 4 Neuroblastoma Cell Lines. Biomed. Pharmacother. 2022, 154, 113584.
- Roomi, M.W.; Kalinovsky, T.; Roomi, N.W.; Niedzwiecki, A.; Rath, M. Inhibition of the SK-N-MC Human Neuroblastoma Cell Line in Vivo and in Vitro by a Novel Nutrient Mixture. Oncol. Rep. 2013, 29, 1714–1720.
- Mazurakova, A.; Samec, M.; Koklesova, L.; Biringer, K.; Kudela, E.; Al-Ishaq, R.K.; Pec, M.; Giordano, F.A.; Büsselberg, D.; Kubatka, P.; et al. Anti-Prostate Cancer Protection and Therapy in the Framework of Predictive, Preventive and Personalised Medicine—Comprehensive Effects of Phytochemicals in Primary, Secondary and Tertiary Care. EPMA J. 2022, 13, 461–486.
- Chakrabarti, M.; Ai, W.; Banik, N.L.; Ray, S.K. Overexpression of MiR-7-1 Increases Efficacy of Green Tea Polyphenols for Induction of Apoptosis in Human Malignant Neuroblastoma SH-SY5Y and SK-N-DZ Cells. Neurochem. Res. 2013, 38, 420–432.
- ISSN 1791-7530 (Online)|Anti-cancer Research|The ISSN Portal. Available online: https://portal.issn.org/resource/ISSN/1791-7530 (accessed on 9 November 2022).
- Picone, P.; Nuzzo, D.; Caruana, L.; Messina, E.; Scafidi, V.; Di Carlo, M. Curcumin Induces Apoptosis in Human Neuroblastoma Cells via Inhibition of AKT and Foxo3a Nuclear Translocation. Free. Radic. Res. 2014, 48, 1397–1408.
- Zhang, J.; Liu, Q.; Shi, L.; Qin, P.; Wang, Q. Honokiol Triggers Receptor-Interacting Protein Kinase 3-Mediated Cell Death of Neuroblastoma Cells by Upregulating Reactive Oxygen Species. Mol. Med. Rep. 2017, 16, 8525–8529.
- George, J.; Banik, N.L.; Ray, S.K. Genistein Induces Receptor and Mitochondrial Pathways and Increases Apoptosis during BCL-2 Knockdown in Human Malignant Neuroblastoma SK-N-DZ Cells. J. Neurosci. Res. 2010, 88, 877–886.
- Seervi, M.; Xue, D. Mitochondrial Cell Death Pathways in Caenorhabiditis Elegans. Curr. Top. Dev. Biol. 2015, 114, 43–65.
More
Information
Subjects:
Biochemistry & Molecular Biology; Oncology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
768
Revisions:
2 times
(View History)
Update Date:
06 Apr 2023
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No