Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | In Soo Kim | -- | 1557 | 2023-03-15 10:01:10 | | | |
| 2 | Sirius Huang | Meta information modification | 1557 | 2023-03-16 02:46:29 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Kim, I.S.; Silwal, P.; Jo, E. Overview of Peroxisome Proliferator-Activated Receptors. Encyclopedia. Available online: https://encyclopedia.pub/entry/42221 (accessed on 26 July 2026).
Kim IS, Silwal P, Jo E. Overview of Peroxisome Proliferator-Activated Receptors. Encyclopedia. Available at: https://encyclopedia.pub/entry/42221. Accessed July 26, 2026.
Kim, In Soo, Prashanta Silwal, Eun-Kyeong Jo. "Overview of Peroxisome Proliferator-Activated Receptors" Encyclopedia, https://encyclopedia.pub/entry/42221 (accessed July 26, 2026).
Kim, I.S., Silwal, P., & Jo, E. (2023, March 15). Overview of Peroxisome Proliferator-Activated Receptors. In Encyclopedia. https://encyclopedia.pub/entry/42221
Kim, In Soo, et al. "Overview of Peroxisome Proliferator-Activated Receptors." Encyclopedia. Web. 15 March, 2023.
Copy Citation
Peroxisome proliferator-activated receptors (PPARs) α, β, and γ are nuclear receptors that orchestrate the transcriptional regulation of genes involved in a variety of biological responses, such as energy metabolism and homeostasis, regulation of inflammation, cellular development, and differentiation. The many roles played by the PPAR signaling pathways indicate that PPARs may be useful targets for various human diseases.
peroxisome proliferator-activated receptor
infection
bacteria
virus
parasite
1. Molecular Characteristics of PPARs
Peroxisomes, 0.5 μm diameter single-membrane cytoplasmic organelles, play essential roles in the oxidation of various biomolecules [1][2]. Peroxisome proliferators are multiple chemicals that increase the abundance of peroxisomes in cells [3][4]. These molecules also increase gene expression for β-oxidation of long-chain fatty acids and cytochrome P450 (CYP450) [5][6]. Given the gene transcriptional modulation of peroxisome proliferators, PPARs have been identified as nuclear receptors [7][8][9][10][11][12][13]. The PPAR subfamily consists of three isoforms, PPARα, PPARβ/δ, and PPARγ [14]. The three PPARs differ in tissue-specific expression patterns and ligand-biding domains, each performing distinct functions. PPARA, encoding PPARα, is located in chromosome 22q13.31 in humans and is mainly expressed in the liver, intestine, kidney, heart, and muscle [15][16]. PPARγ has four alternative splicing forms from PPARG located in chromosome 3p25.2 and is highly expressed in adipose tissue, the spleen, and intestine [17][18]. PPARδ, encoded by PPARD, is located in chromosome 6q21.31 and presents ubiquitously [13][19]. Thus, it is essential in the study of PPARs to consider their tissue distribution and functions.
PPAR is a nuclear receptor superfamily class II member that heterodimerizes with RXR [20][21]. The PPAR structure includes the A/B, C, D, and E domains from N-terminus to C-terminus [22]. The N-terminal A/B domain (NTD) is a ligand-independent transactivation domain containing the activator function (AF)-1 region. The NTD is targeted for variable post-translational modifications, including SUMOylation, phosphorylation, acetylation, O-GlcNAcylation, and ubiquitination, resulting in transcriptional regulatory activities [23]. DNA-binding C domain (DBD) has two DNA-binding zinc finger motifs containing cysteines, which dock to PPREs. PPARs reside upstream of RXR upon the direct repeat (DR)-1 motifs, which are composed of two hexanucleotide consensus sequences with one spacing nucleotide (AGGTCA N AGGTCA) [24]. The hinge D region is a linker between the C and E domains, which contains a nuclear localization signal, and is the site for post-translational modifications such as phosphorylation, acetylation, and SUMOylation [23]. The ligand-binding E domain (LBD) carries the hydrophobic ligand-binding pocket and the AF-2 region. The absence of agonists enables LBD to recruit co-repressors containing the CoRNR motifs [25]. Engaging agonists to LBD elicits conformational changes of AF-2 to facilitate interaction with LXXLL motifs of many co-activators [26]. Like other nuclear receptor superfamily class II members, such as thyroid hormone receptor (TR), retinoic acid receptor (RAR), and vitamin D receptor (VDR), PPARs function as heterodimers with RXR through LBD [27][28]. LBD is also targeted for SUMOylation and ubiquitination [23]. Advancement of research on the PPAR structure helps thoroughly dissect the roles of PPARs. The roles of specific PPAR subtypes will be discussed in the following subsections.
1.1. Roles of PPARα
PPARα is predominantly expressed in the liver but is also found in other tissues, including the heart, muscle, and kidney [16][29]. PPARα regulates the expression of genes involved in metabolism and inflammation. Activation of PPARα leads to the upregulation of genes involved in fatty acid oxidation and the downregulation of genes involved in fatty acid synthesis [30]. PPARα also modulates other genes, including genes involved in the transport and uptake of fatty acids and the synthesis and secretion of lipoproteins [29][30]. In addition, activation of PPARα has been shown to improve insulin sensitivity, reduce oxidative stress, and reduce inflammation in preclinical studies [30][31][32]. PPARα activation has been shown to modify the expression of immune response genes, including those encoding cytokines and chemokines, which are signaling molecules that regulate the immune response [32][33]. PPARα activation has also been demonstrated to reduce the production of pro-inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α) and interleukin (IL)-6 [34][35]. PPARα has been shown to interfere with the DNA binding of both AP-1 and NF-κB [33][34][36]. Thus, the roles of PPARα in infectious diseases should be studied in wide ranging aspects, including metabolism and inflammation.
In the context of infection, PPARα has been shown to play an essential role in the hepatic metabolic response to infection. During an infectious challenge, the liver coordinates several metabolic changes to support the host defense response, including the mobilization of energy stores, production of acute-phase proteins, and synthesis of new metabolites. Activation of PPARα in the liver leads to the upregulation of genes involved in fatty acid oxidation and ketogenesis with fibroblast growth factor 21 (FGF21) production [37]. FGF21 is a hormone produced by the liver that has been shown to promote ketogenesis and reduce glucose utilization [38][39]. The ketogenesis regulation of PPARα with FGF21 is essential for reacting to microbial or viral sepsis [40][41][42]. In conclusion, the hepatic PPARα metabolic response to infection is crucial to the host defense response.
1.2. Roles of PPARβ/δ
PPARβ is expressed in diverse tissues, including adipose tissue, muscle, and the liver [13][43], and is activated by multiple ligands, such as fatty acids and their derivatives [31]. PPARβ is involved in regulating lipid metabolism and energy homeostasis, as well as controlling inflammation and immune function [44]. PPARβ activation has been demonstrated to have pro- and anti-inflammatory effects based on the situation [44]. The role of PPARβ in tumorigenesis is debatable. PPARβ activation has been found in some cases to have anti-tumorigenic effects, such as causing apoptosis and inhibiting cell proliferation [45][46]. In other cases, however, activation of PPARβ has been shown to promote tumorigenesis by enhancing cell survival, promoting angiogenesis, and reducing cellular differentiation [47][48][49][50]. Overall, the role of PPARβ activation in cancer is not entirely known and is complex. Similarly, the function of PPARβ in infection is not well understood. Additional research is required to comprehend the function of PPARβ in the context of immunology against cancers and infectious diseases.
1.3. Roles of PPARγ
PPARγ is expressed in a variety of tissues, including adipose tissue, muscle, and the liver [17][18][43], and is activated by diverse ligands, including fatty acids and their derivatives, as well as synthetic chemicals known as thiazolidinediones [29][31]. PPARγ is responsible for regulating lipid metabolism, glucose homeostasis, and inflammation [51][52]. Numerous inflammatory mediators and cytokines are inhibited by PPARγ ligands in various cell types, including monocytes/macrophages, epithelial cells, smooth muscle cells, endothelial cells, dendritic cells, and lymphocytes. In addition, PPARγ diminishes the activities of transcription factors AP-1, STAT, NF-κB, and NFAT to adversely regulate inflammatory gene expression [53][54][55]. As a result, PPARγ has been demonstrated to have a protective function against infections by modulating the immune response and lowering inflammation. However, other researchers have hypothesized that PPARγ activation may impair the function of immune cells, such as macrophages, and contribute to the development of infections. Therefore, the role of PPARγ in disease is complex and context-dependent, and more research is needed to fully understand the molecular mechanisms by which PPARγ regulates the host response to infection.
2. Regulatory Mechanisms of PPARs
The PPAR ligand-binding pocket is large and capable of engaging diverse ligands [56][57]. Endogenous ligands vary depending on the PPAR isoform, including n-3 polyunsaturated fatty acids such as docosahexaenoic acid and eicosapentaenoic acid for all PPARs, leukotriene B4 for PPARα, carbaprostacyclin for PPARδ, and prostaglandin J2 for PPARγ [58]. Representative synthetic agonists include fibrates (PPARα agonists) and thiazolidinediones (PPARγ agonists) [31]. Fibrates, such as fenofibrate, clofibrate, and gemfibrozil, are widely used for treating dyslipidemia. Thiazolidinediones, such as rosiglitazone, pioglitazone, and lobeglitazone, improve insulin resistance [31]. Most clinical studies on PPAR actions in infectious diseases have been conducted retrospectively, and no clinical studies currently in progress are listed in ClinicalTrials.gov (https://clinicaltrials.gov/ (accessed on 13 February 2023)). Since widely used PPAR agonists exist, clinical research can be conducted through a deeper understanding of PPAR roles in infectious diseases.
PPAR-RXR heterodimerization occurs ligand-independently [27]. The heterodimer appears to exert transcriptional regulation both ligand-dependently and -independently [31]. Although LBD may interact with either co-repressor or co-activator in the state of not binding with an agonist, binding to a ligand elicits stabilized co-activator-LBD interaction, thus increasing transactivation [31][59]. Further, recent studies have shown that PPARs inhibit other transcription factors, such as NF-κB, activator protein-1 (AP-1), signal transducer and activator of transcription (STAT), and nuclear factor of activated T cells (NFAT) [32][53][54][55]. Recent studies revealed the possibility of forming a protein chaperone complex with PPAR-associated proteins, such as heat shock proteins (HSPs). Similar to interactions between other type I intracellular receptors and heat shock proteins, HSP90 repressed PPARα and PPARβ activities but not that of PPARγ [60]. Instead, HSP90 was required for PPARγ signaling in the nonalcoholic fatty liver disease mouse model [61]. Thus, it is necessary to study the various modes of PPAR actions. The intracellular regulatory mechanisms of PPARs are shown in Figure 1.
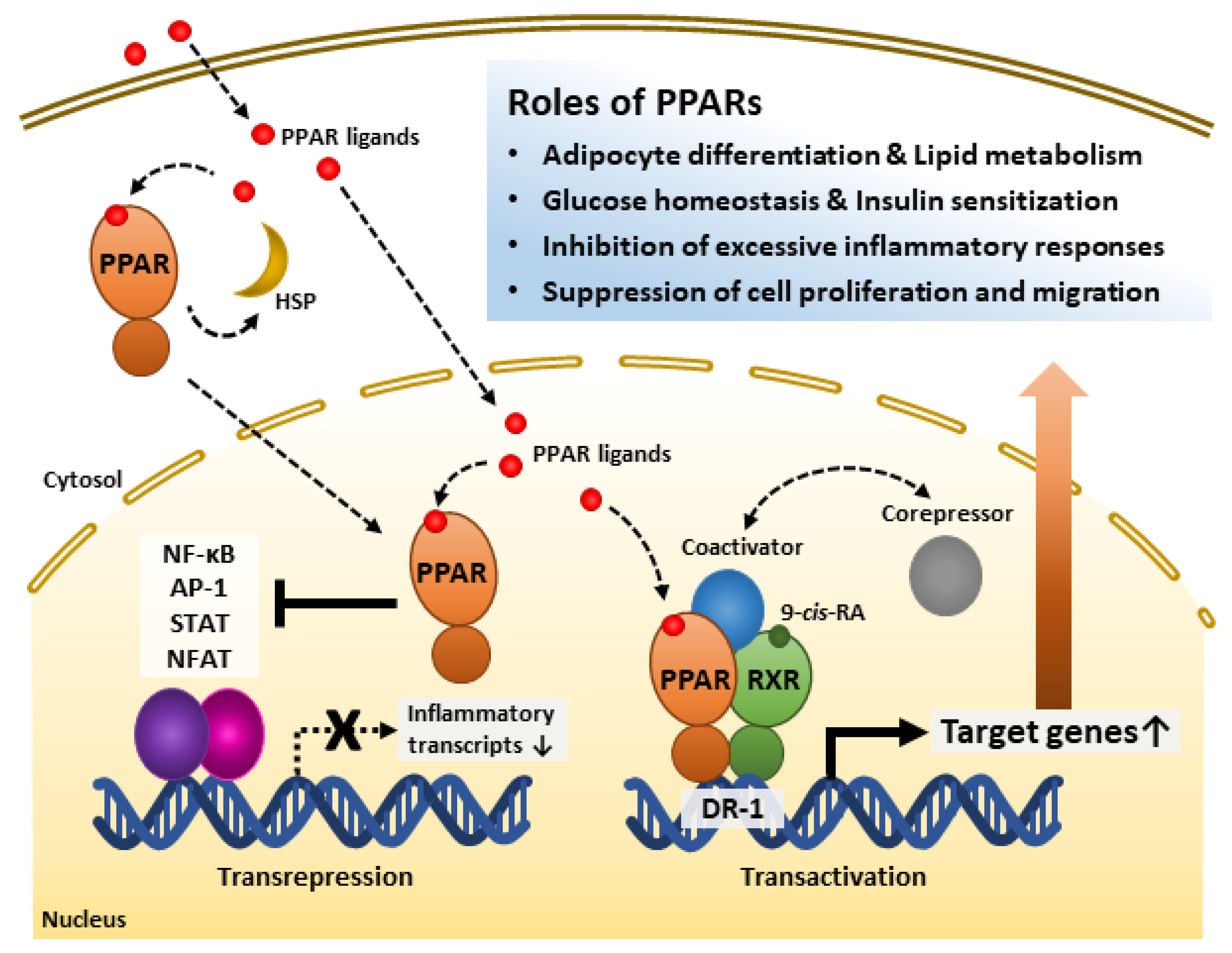
Figure 1. Roles of peroxisome proliferator-activated receptors (PPARs) and their regulatory mechanisms. PPAR ligands bind to the PPAR ligand-binding domain and activate receptors. PPARs interact with heat shock protein (HSP) in the cytosol. PPARs inhibit inflammation-related gene transcription by interfering with transcription factors such as NF-κB, AP-1, STAT, and NFAT. PPARs form heterodimers with Retinoid X receptor (RXR), a receptor of 9-cis-retinoic acid (9-cis-RA), and bind to direct repeat 1 (DR-1), a peroxisome-proliferator-responsive element. The PPAR-RXR heterodimer complex and co-repressors represses target gene transcription. However, the complex with co-activators promotes target gene transcription. Through these mechanisms, PPARs play significant roles in energy metabolism, inflammatory modulation, and the cell cycle. AP-1, Activator protein 1; NF-κB, Nuclear factor kappa-light-chain-enhancer of activated B cells; NFAT, Nuclear factor of activated T cells; STAT, Signal transducer and activator of transcription.
References
- De Duve, C.; Baudhuin, P. Peroxisomes (microbodies and related particles). Physiol. Rev. 1966, 46, 323–357.
- Okumoto, K.; Tamura, S.; Honsho, M.; Fujiki, Y. Peroxisome: Metabolic Functions and Biogenesis. Adv. Exp. Med. Biol. 2020, 1299, 3–17.
- Reddy, J.K.; Krishnakantha, T.P.; Azarnoff, D.L.; Moody, D.E. 1-methyl-4piperidyl-bis (P-chlorophenoxy) acetate: A new hypolipidemic peroxisome proliferator. Res. Commun. Chem. Pathol. Pharmacol. 1975, 10, 589–592.
- Reddy, J.K.; Lalwai, N.D. Carcinogenesis by hepatic peroxisome proliferators: Evaluation of the risk of hypolipidemic drugs and industrial plasticizers to humans. Crit. Rev. Toxicol. 1983, 12, 1–58.
- Reddy, J.K.; Goel, S.K.; Nemali, M.R.; Carrino, J.J.; Laffler, T.G.; Reddy, M.K.; Sperbeck, S.J.; Osumi, T.; Hashimoto, T.; Lalwani, N.D.; et al. Transcription regulation of peroxisomal fatty acyl-CoA oxidase and enoyl-CoA hydratase/3-hydroxyacyl-CoA dehydrogenase in rat liver by peroxisome proliferators. Proc. Natl. Acad. Sci. USA 1986, 83, 1747–1751.
- Hardwick, J.P.; Song, B.J.; Huberman, E.; Gonzalez, F.J. Isolation, complementary DNA sequence, and regulation of rat hepatic lauric acid omega-hydroxylase (cytochrome P-450LA omega). Identification of a new cytochrome P-450 gene family. J. Biol. Chem. 1987, 262, 801–810.
- Issemann, I.; Green, S. Activation of a member of the steroid hormone receptor superfamily by peroxisome proliferators. Nature 1990, 347, 645–650.
- Zhu, Y.; Alvares, K.; Huang, Q.; Rao, M.S.; Reddy, J.K. Cloning of a new member of the peroxisome proliferator-activated receptor gene family from mouse liver. J. Biol. Chem. 1993, 268, 26817–26820.
- Göttlicher, M.; Widmark, E.; Li, Q.; Gustafsson, J.A. Fatty acids activate a chimera of the clofibric acid-activated receptor and the glucocorticoid receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 4653–4657.
- Rolls, B.J.; Hammer, V.A. Fat, carbohydrate, and the regulation of energy intake. Am. J. Clin. Nutr. 1995, 62, 1086s–1095s.
- Dreyer, C.; Krey, G.; Keller, H.; Givel, F.; Helftenbein, G.; Wahli, W. Control of the peroxisomal beta-oxidation pathway by a novel family of nuclear hormone receptors. Cell 1992, 68, 879–887.
- Greene, M.E.; Blumberg, B.; McBride, O.W.; Yi, H.F.; Kronquist, K.; Kwan, K.; Hsieh, L.; Greene, G.; Nimer, S.D. Isolation of the human peroxisome proliferator activated receptor gamma cDNA: Expression in hematopoietic cells and chromosomal mapping. Gene Expr. 1995, 4, 281–299.
- Schmidt, A.; Endo, N.; Rutledge, S.J.; Vogel, R.; Shinar, D.; Rodan, G.A. Identification of a new member of the steroid hormone receptor superfamily that is activated by a peroxisome proliferator and fatty acids. Mol. Endocrinol. 1992, 6, 1634–1641.
- Christofides, A.; Konstantinidou, E.; Jani, C.; Boussiotis, V.A. The role of peroxisome proliferator-activated receptors (PPAR) in immune responses. Metabolism 2021, 114, 154338.
- Sher, T.; Yi, H.F.; McBride, O.W.; Gonzalez, F.J. cDNA cloning, chromosomal mapping, and functional characterization of the human peroxisome proliferator activated receptor. Biochemistry 1993, 32, 5598–5604.
- Tyagi, S.; Gupta, P.; Saini, A.S.; Kaushal, C.; Sharma, S. The peroxisome proliferator-activated receptor: A family of nuclear receptors role in various diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236–240.
- Qi, J.S.; Desai-Yajnik, V.; Greene, M.E.; Raaka, B.M.; Samuels, H.H. The ligand-binding domains of the thyroid hormone/retinoid receptor gene subfamily function in vivo to mediate heterodimerization, gene silencing, and transactivation. Mol. Cell. Biol. 1995, 15, 1817–1825.
- Evans, R.M.; Barish, G.D.; Wang, Y.X. PPARs and the complex journey to obesity. Nat. Med. 2004, 10, 355–361.
- Wang, S.; Dougherty, E.J.; Danner, R.L. PPARγ signaling and emerging opportunities for improved therapeutics. Pharmacol. Res. 2016, 111, 76–85.
- Bain, D.L.; Heneghan, A.F.; Connaghan-Jones, K.D.; Miura, M.T. Nuclear receptor structure: Implications for function. Annu. Rev. Physiol. 2007, 69, 201–220.
- Weikum, E.R.; Liu, X.; Ortlund, E.A. The nuclear receptor superfamily: A structural perspective. Protein Sci. 2018, 27, 1876–1892.
- Desvergne, B.; Wahli, W. Peroxisome proliferator-activated receptors: Nuclear control of metabolism. Endocr. Rev. 1999, 20, 649–688.
- Brunmeir, R.; Xu, F. Functional Regulation of PPARs through Post-Translational Modifications. Int. J. Mol. Sci. 2018, 19, 1738.
- Palmer, C.N.; Hsu, M.H.; Griffin, H.J.; Johnson, E.F. Novel sequence determinants in peroxisome proliferator signaling. J. Biol. Chem. 1995, 270, 16114–16121.
- Hu, X.; Lazar, M.A. The CoRNR motif controls the recruitment of corepressors by nuclear hormone receptors. Nature 1999, 402, 93–96.
- Poulsen, L.; Siersbæk, M.; Mandrup, S. PPARs: Fatty acid sensors controlling metabolism. Semin. Cell Dev. Biol. 2012, 23, 631–639.
- Chandra, V.; Huang, P.; Hamuro, Y.; Raghuram, S.; Wang, Y.; Burris, T.P.; Rastinejad, F. Structure of the intact PPAR-gamma-RXR- nuclear receptor complex on DNA. Nature 2008, 456, 350–356.
- Chan, L.S.; Wells, R.A. Cross-Talk between PPARs and the Partners of RXR: A Molecular Perspective. PPAR Res. 2009, 2009, 925309.
- Lamas Bervejillo, M.; Ferreira, A.M. Understanding Peroxisome Proliferator-Activated Receptors: From the Structure to the Regulatory Actions on Metabolism. Adv. Exp. Med. Biol. 2019, 1127, 39–57.
- Ye, X.; Zhang, T.; Han, H. PPARα: A potential therapeutic target of cholestasis. Front. Pharmacol. 2022, 13, 916866.
- Grygiel-Górniak, B. Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical implications--a review. Nutr. J. 2014, 13, 17.
- Ricote, M.; Glass, C.K. PPARs and molecular mechanisms of transrepression. Biochim. Biophys. Acta 2007, 1771, 926–935.
- Delerive, P.; Gervois, P.; Fruchart, J.C.; Staels, B. Induction of IkappaBalpha expression as a mechanism contributing to the anti-inflammatory activities of peroxisome proliferator-activated receptor-alpha activators. J. Biol. Chem. 2000, 275, 36703–36707.
- Staels, B.; Koenig, W.; Habib, A.; Merval, R.; Lebret, M.; Torra, I.P.; Delerive, P.; Fadel, A.; Chinetti, G.; Fruchart, J.C.; et al. Activation of human aortic smooth-muscle cells is inhibited by PPARalpha but not by PPARgamma activators. Nature 1998, 393, 790–793.
- Madej, A.; Okopien, B.; Kowalski, J.; Zielinski, M.; Wysocki, J.; Szygula, B.; Kalina, Z.; Herman, Z.S. Effects of fenofibrate on plasma cytokine concentrations in patients with atherosclerosis and hyperlipoproteinemia IIb. Int. J. Clin. Pharmacol. Ther. 1998, 36, 345–349.
- Delerive, P.; Martin-Nizard, F.; Chinetti, G.; Trottein, F.; Fruchart, J.C.; Najib, J.; Duriez, P.; Staels, B. Peroxisome proliferator-activated receptor activators inhibit thrombin-induced endothelin-1 production in human vascular endothelial cells by inhibiting the activator protein-1 signaling pathway. Circ. Res. 1999, 85, 394–402.
- Kersten, S.; Seydoux, J.; Peters, J.M.; Gonzalez, F.J.; Desvergne, B.; Wahli, W. Peroxisome proliferator-activated receptor alpha mediates the adaptive response to fasting. J. Clin. Invest. 1999, 103, 1489–1498.
- Montagner, A.; Polizzi, A.; Fouché, E.; Ducheix, S.; Lippi, Y.; Lasserre, F.; Barquissau, V.; Régnier, M.; Lukowicz, C.; Benhamed, F.; et al. Liver PPARα is crucial for whole-body fatty acid homeostasis and is protective against NAFLD. Gut 2016, 65, 1202–1214.
- Fougerat, A.; Schoiswohl, G.; Polizzi, A.; Régnier, M.; Wagner, C.; Smati, S.; Fougeray, T.; Lippi, Y.; Lasserre, F.; Raho, I.; et al. ATGL-dependent white adipose tissue lipolysis controls hepatocyte PPARα activity. Cell Rep. 2022, 39, 110910.
- Wang, A.; Huen, S.C.; Luan, H.H.; Yu, S.; Zhang, C.; Gallezot, J.D.; Booth, C.J.; Medzhitov, R. Opposing Effects of Fasting Metabolism on Tissue Tolerance in Bacterial and Viral Inflammation. Cell 2016, 166, 1512–1525.e1512.
- Huen, S.C.; Wang, A.; Feola, K.; Desrouleaux, R.; Luan, H.H.; Hogg, R.; Zhang, C.; Zhang, Q.J.; Liu, Z.P.; Medzhitov, R. Hepatic FGF21 preserves thermoregulation and cardiovascular function during bacterial inflammation. J. Exp. Med. 2021, 218, e20202151.
- Paumelle, R.; Haas, J.T.; Hennuyer, N.; Baugé, E.; Deleye, Y.; Mesotten, D.; Langouche, L.; Vanhoutte, J.; Cudejko, C.; Wouters, K.; et al. Hepatic PPARα is critical in the metabolic adaptation to sepsis. J. Hepatol. 2019, 70, 963–973.
- Braissant, O.; Foufelle, F.; Scotto, C.; Dauça, M.; Wahli, W. Differential expression of peroxisome proliferator-activated receptors (PPARs): Tissue distribution of PPAR-alpha, -beta, and -gamma in the adult rat. Endocrinology 1996, 137, 354–366.
- Müller, R. PPARβ/δ in human cancer. Biochimie 2017, 136, 90–99.
- Yao, P.L.; Chen, L.; Dobrzański, T.P.; Zhu, B.; Kang, B.H.; Müller, R.; Gonzalez, F.J.; Peters, J.M. Peroxisome proliferator-activated receptor-β/δ inhibits human neuroblastoma cell tumorigenesis by inducing p53- and SOX2-mediated cell differentiation. Mol. Carcinog. 2017, 56, 1472–1483.
- Foreman, J.E.; Sharma, A.K.; Amin, S.; Gonzalez, F.J.; Peters, J.M. Ligand activation of peroxisome proliferator-activated receptor-beta/delta (PPARbeta/delta) inhibits cell growth in a mouse mammary gland cancer cell line. Cancer Lett. 2010, 288, 219–225.
- Genini, D.; Garcia-Escudero, R.; Carbone, G.M.; Catapano, C.V. Transcriptional and Non-Transcriptional Functions of PPARβ/δ in Non-Small Cell Lung Cancer. PLoS ONE 2012, 7, e46009.
- Pedchenko, T.V.; Gonzalez, A.L.; Wang, D.; DuBois, R.N.; Massion, P.P. Peroxisome proliferator-activated receptor beta/delta expression and activation in lung cancer. Am. J. Respir. Cell Mol. Biol. 2008, 39, 689–696.
- Wagner, K.D.; Benchetrit, M.; Bianchini, L.; Michiels, J.F.; Wagner, N. Peroxisome proliferator-activated receptor β/δ (PPARβ/δ) is highly expressed in liposarcoma and promotes migration and proliferation. J. Pathol. 2011, 224, 575–588.
- Bishop-Bailey, D. PPARs and angiogenesis. Biochem. Soc. Trans. 2011, 39, 1601–1605.
- Daynes, R.A.; Jones, D.C. Emerging roles of PPARs in inflammation and immunity. Nat. Rev. Immunol. 2002, 2, 748–759.
- Kostadinova, R.; Wahli, W.; Michalik, L. PPARs in diseases: Control mechanisms of inflammation. Curr. Med. Chem. 2005, 12, 2995–3009.
- Ricote, M.; Li, A.C.; Willson, T.M.; Kelly, C.J.; Glass, C.K. The peroxisome proliferator-activated receptor-gamma is a negative regulator of macrophage activation. Nature 1998, 391, 79–82.
- Chinetti, G.; Griglio, S.; Antonucci, M.; Torra, I.P.; Delerive, P.; Majd, Z.; Fruchart, J.C.; Chapman, J.; Najib, J.; Staels, B. Activation of proliferator-activated receptors alpha and gamma induces apoptosis of human monocyte-derived macrophages. J. Biol. Chem. 1998, 273, 25573–25580.
- Bao, Y.; Li, R.; Jiang, J.; Cai, B.; Gao, J.; Le, K.; Zhang, F.; Chen, S.; Liu, P. Activation of peroxisome proliferator-activated receptor gamma inhibits endothelin-1-induced cardiac hypertrophy via the calcineurin/NFAT signaling pathway. Mol. Cell. Biochem. 2008, 317, 189–196.
- Kamata, S.; Oyama, T.; Saito, K.; Honda, A.; Yamamoto, Y.; Suda, K.; Ishikawa, R.; Itoh, T.; Watanabe, Y.; Shibata, T.; et al. PPARα Ligand-Binding Domain Structures with Endogenous Fatty Acids and Fibrates. iScience 2020, 23, 101727.
- Nolte, R.T.; Wisely, G.B.; Westin, S.; Cobb, J.E.; Lambert, M.H.; Kurokawa, R.; Rosenfeld, M.G.; Willson, T.M.; Glass, C.K.; Milburn, M.V. Ligand binding and co-activator assembly of the peroxisome proliferator-activated receptor-gamma. Nature 1998, 395, 137–143.
- Krey, G.; Braissant, O.; L’Horset, F.; Kalkhoven, E.; Perroud, M.; Parker, M.G.; Wahli, W. Fatty acids, eicosanoids, and hypolipidemic agents identified as ligands of peroxisome proliferator-activated receptors by coactivator-dependent receptor ligand assay. Mol. Endocrinol. 1997, 11, 779–791.
- Aagaard, M.M.; Siersbæk, R.; Mandrup, S. Molecular basis for gene-specific transactivation by nuclear receptors. Biochim. Biophys. Acta 2011, 1812, 824–835.
- Sumanasekera, W.K.; Tien, E.S.; Davis, J.W., 2nd; Turpey, R.; Perdew, G.H.; Vanden Heuvel, J.P. Heat shock protein-90 (Hsp90) acts as a repressor of peroxisome proliferator-activated receptor-alpha (PPARalpha) and PPARbeta activity. Biochemistry 2003, 42, 10726–10735.
- Wheeler, M.C.; Gekakis, N. Hsp90 modulates PPARγ activity in a mouse model of nonalcoholic fatty liver disease. J. Lipid Res. 2014, 55, 1702–1710.
More
Information
Subjects:
Infectious Diseases
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
2.4K
Revisions:
2 times
(View History)
Update Date:
16 Mar 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No