Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Annamaria Lia | -- | 1993 | 2022-12-06 09:23:08 | | | |
| 2 | Vivi Li | + 45 word(s) | 2038 | 2022-12-07 06:14:33 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Arnst, N.; Redolfi, N.; Lia, A.; Bedetta, M.; Greotti, E.; Pizzo, P. Mitochondrial Ca2+ Signaling and Bioenergetics in Alzheimer’s Disease. Encyclopedia. Available online: https://encyclopedia.pub/entry/38109 (accessed on 23 July 2026).
Arnst N, Redolfi N, Lia A, Bedetta M, Greotti E, Pizzo P. Mitochondrial Ca2+ Signaling and Bioenergetics in Alzheimer’s Disease. Encyclopedia. Available at: https://encyclopedia.pub/entry/38109. Accessed July 23, 2026.
Arnst, Nikita, Nelly Redolfi, Annamaria Lia, Martina Bedetta, Elisa Greotti, Paola Pizzo. "Mitochondrial Ca2+ Signaling and Bioenergetics in Alzheimer’s Disease" Encyclopedia, https://encyclopedia.pub/entry/38109 (accessed July 23, 2026).
Arnst, N., Redolfi, N., Lia, A., Bedetta, M., Greotti, E., & Pizzo, P. (2022, December 06). Mitochondrial Ca2+ Signaling and Bioenergetics in Alzheimer’s Disease. In Encyclopedia. https://encyclopedia.pub/entry/38109
Arnst, Nikita, et al. "Mitochondrial Ca2+ Signaling and Bioenergetics in Alzheimer’s Disease." Encyclopedia. Web. 06 December, 2022.
Copy Citation
Alzheimer’s disease (AD) is a hereditary and sporadic neurodegenerative illness defined by the gradual and cumulative loss of neurons in specific brain areas. The processes that cause AD are still under investigation and there are no available therapies to halt it. Progress puts at the forefront the “calcium (Ca2+) hypothesis” as a key AD pathogenic pathway, impacting neuronal, astrocyte and microglial function. An increasing body of evidence points out the early and crucial role of cellular Ca2+ handling dysregulation in AD pathogenesis. Interestingly, Ca2+ is a key regulator of several mitochondrial functions, such as ATP production, and brain cells rely mostly on OXPHOS to match their energy demands.
calcium
Alzheimer’s disease
mitochondria
bioenergetics
neuron
microglia
astrocyte
AD mouse model
iPSCs
1. Introduction
Mitochondria are well-known organelles that function as the powerhouse of the cell. They occupy approximately 20% of the cell volume, although their number and size vary from cell to cell. Their presence is massive in excitable cells, which rely strongly on oxidative phosphorylation (OXPHOS) to produce the adenosine triphosphate (ATP) necessary to accomplish their physiological functions [1]. However, mitochondria are much more than mere producers of energy. Decades of research have shown that mitochondria represent a formidable intracellular signaling platform that orchestrates various cellular functions. They are crucial, for example, in calcium (Ca2+) handling, regulation of cell response to stress, autophagy and cell death. In these processes, their tight interaction with other organelles, in particular the endoplasmic reticulum (ER), is fundamental [2][3]. The central role of mitochondria in cellular physiology is such that alterations in their functionality are involved in the pathogenesis of various disorders, including Alzheimer’s disease (AD).
AD is the most common neurodegenerative disease caused by poorly known pathogenic mechanisms, affecting around 10% of the population aged more than 65 in the United States [4] and around 5% of the European population [5]. Although the sporadic form (SAD) is the most prevalent, the presence of mutations in the amyloid precursor protein (APP), presenilin-1 (PSEN1) and presenilin-2 (PSEN2) genes are marks of familial cases (FAD). FAD forms represent a small percentage (2–3%) of AD cases, and PSEN1 and PSEN2 mutations account for more than 80% of genetic lesions [6]. FAD is characterized by an early onset and a worse prognosis compared to SAD, despite that they share many clinical manifestations and pathological hallmarks, i.e., accumulation and deposition of brain extracellular amyloid beta (Aβ) plaques and intracellular neurofibrillary tangles (NFTs), made by aggregates of hyper-phosphorylated tau. Besides memory and cognitive dysfunctions, behavioral and psychological symptoms of dementia (BPSD) are emerging as key clinical manifestations of AD. The severity of BPSD is shared in FAD and SAD, although their onset differs between the two groups, with a prevalence of BPSD higher in FAD compared to SAD [7]. In an attempt to clarify the pathogenic mechanisms leading to the development of AD, FAD-linked mutations have been exploited for the development of animal models that are useful for preclinical studies [8].
Over time, several hypotheses have been proposed to explain the pathogenesis of AD. The first and the most studied ones focus on the accumulation and deposition of Aβ plaques (amyloid cascade hypothesis [9]) and NFTs (tau hypothesis [10]). Despite immeasurable investigations performed in the last few decades, there are still no efficient disease-modifying treatments for AD [11]. Indeed, many clinical trials targeting Aβ and tau have failed, and the disease still lacks a cure [12]. More recently, other signaling pathways have also been investigated as pathogenic in AD, leading to the formulation of the infectious hypothesis, the inflammation hypothesis, the cholinergic hypothesis, the glutamatergic hypothesis, and many others (summarized here [13][14]). Various mitochondrial defects, such as decreased bioenergetics and ATP synthesis, increased ROS (reactive oxygen species) production, and altered mitochondrial transport and dynamics, have been reported to contribute to synaptic dysfunction and neuronal cell death in both SAD and FAD [15], leading to the formulation of the mitochondrial cascade hypothesis [16].
An increasing body of evidence points out the early and crucial role of cellular Ca2+ handling dysregulation in AD pathogenesis [17]. Interestingly, Ca2+ is a key regulator of several mitochondrial functions, such as ATP production, and brain cells rely mostly on OXPHOS to match their energy demands. Moreover, mitochondrial Ca2+ alterations may affect the functionality of other brain cells involved in memory formation and consolidation. Indeed, a growing number of studies have revealed that memory formation is not only a matter of neuronal interactions, but it also crucially involves glial cells [18]. As a result, the classical synaptic framework, consisting only of pre- and post-synaptic compartments, has been gradually expanded to include astrocytes [19] and microglia [20]. The involvement of these cell types in AD pathogenesis has also been confirmed by genome-wide association studies (GWAS). These studies describe SAD-associated genetic polymorphisms in genes involved in microglia, astrocytes and neuronal functions [21][22][23][24], as well as genes encoding mitochondria complexes or proteins involved in energy metabolism [25][26][27][28].
Thus, starting from the recent concept of quad-partite synapse and its importance in the determination of brain functions, researchers will here summarize the evidence of mitochondrial Ca2+ alterations and their consequences on cell bioenergetics in AD, focusing on neurons, astrocytes and microglia. Researchers' overview will span different experimental disease models, focusing in particular on in vivo studies, but including also those based on Induced Pluripotent Stem Cells (iPSCs) that, by filling the gap due to the large evolutionary distance between mice and humans [29], represent a novel and promising tool to model human neurodegenerative diseases [30].
2. The Physiology of Brain Mitochondria: Ca2+ and Bioenergetics
Mitochondria are dynamic organelles able to change size, shape and position in a few seconds. They can move along the cytoskeleton to reach specific subcellular areas, or they undergo fission and fusion to constantly remodel their network and match local energy needs [31]. The organelles are equipped with a double membrane, the internal (IMM) and outer (OMM) mitochondrial membranes, which define the intermembrane space (IMS) and the mitochondrial matrix. The latter presents Ca2+ buffering capacity and participates in several Ca2+-mediated signaling pathways. The IMM is impermeable, even to small molecules, and hosts the electron transport chain (ETC) proteins devoted to OXPHOS. To maximize ATP synthesis, the IMM has many in-folding processes that span the mitochondrial matrix, where tricarboxylic acid (TCA, also called the Krebs cycle) occurs. The OMM is permeable to molecules up to 5 kDa because of the presence of the porin VDAC (Voltage-Dependent Anion Channel) [32]. This means that the IMS small molecule composition is similar to that of the cytosol, whereas molecules with a molecular weight > 5 kDa need specific transporters. The OMM is also the membrane involved in interactions with other organelles. Among them, the ER is one of the most important and its domains closely interacting with the OMM are called Mitochondria-Associated Membranes (MAMs). MAMs regulate numerous cellular processes, such as ER-mitochondria Ca2+ shuttling, lipid synthesis, inflammatory response, autophagy and apoptosis. These highly specialized subcellular domains are emerging as powerful signaling platforms and have been the subject of extensive studies in recent years [33]. Of note, these domains of close apposition between the ER and mitochondria result in alterations in several neurodegenerative diseases, including AD (see below and Figure 1). Although the molecular identity of proteins involved in organelle tethering is still elusive, a cell-specific molecular composition of MAMs is emerging. Recently, a neuronal-specific protein, PDZD8, has been shown to be essential for maintaining ER-mitochondria juxtaposition and neuronal Ca2+ homeostasis [34]. Moreover, Ooi et al. showed that in stress-induced hypertension rats, activation of the Sigma-1 receptor, a chaperone that localizes at MAMs [35], reduces microglia M1 polarization and neuroinflammation through MAM and mitochondrial activity modulation [36]. In astrocytes, MAMs are enriched at the endfeet, i.e., in the astrocyte processes in close contact with vessels [37], but no astrocytic-specific MAM proteins are known nowadays.
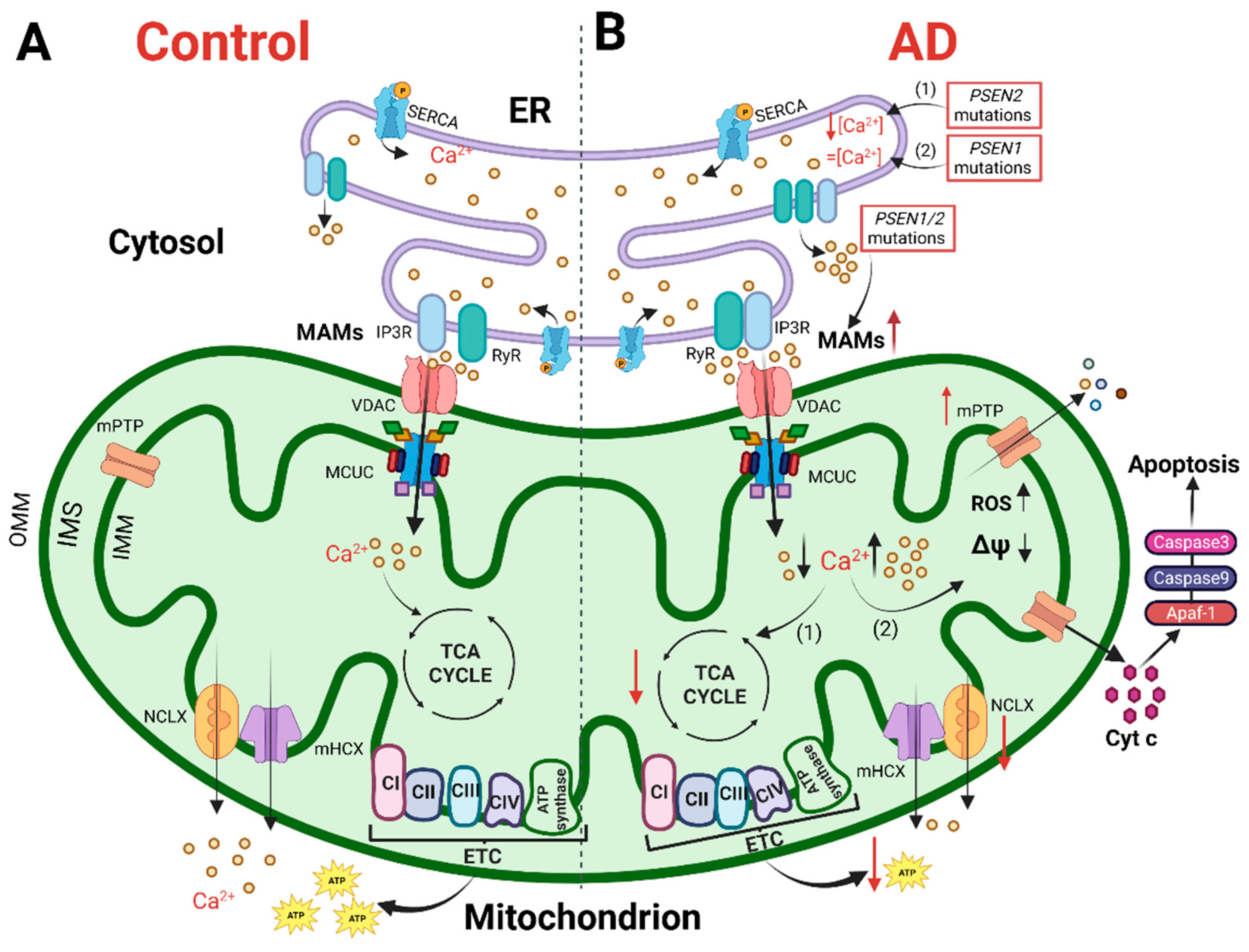
Figure 1. The cartoon shows mitochondrial Ca2+ handling and Ca2+-dependent bioenergetic functions in both physiological conditions (left) and AD (right). (A) In physiological conditions, the Ca2+ cross-talk between ER and mitochondria at MAMs, mediated by Inositol 1,4,5-trisphosphate receptors (IP3R) and ryanodine receptors (RyR) on the ER and VDAC and mitochondrial Ca2+ uniporter (MCU) complex (MCUC) on mitochondria, regulates mitochondrial Ca2+ entry, sustaining the TCA cycle and ATP synthesis. (B) In AD, various Ca2+ alterations have been reported. (1) Mutations in PSEN2 cause a reduction in the ER Ca2+ content, whereas (2) mutations in PSEN1 do not alter the ER Ca2+ content. Mutations in PSEN1/2, by increasing the open probability of ER Ca2+ releasing channels, cause an excessive release of Ca2+ from this store. Both a mitochondrial Ca2+ overload and blunted mitochondrial Ca2+ uptake ability have been described. The first stimulates a sustained production of ROS, the opening of mitochondrial permeability transition pore (mPTP) and the release of cytochrome c activating the apoptotic pathway. The second causes a decrease in ATP production, leading to a bioenergetic crisis. This original figure was created by the authors using “BioRender.com” (https://biorender.com, accessed on 7 November 2022).
3. Mitochondria in Alzheimer’s Disease: Ca2+ Signaling and Bioenergetics
More than 30 years ago, Khachaturian first described a Ca2+ homeostasis alteration in different AD cells [38][39]. From this original observation, the so-called “Ca2+ cascade hypothesis for AD” has been proposed [40], in which neurons from aged and diseased brains experience cytosolic Ca2+ overload upon depolarization [41]. Furthermore, mitochondrial Ca2+ alterations, as well as dysfunctional mitochondria, have been reported in various models of AD, including postmortem patient-derived samples, leading to the formulation of the “mitochondrial cascade hypothesis for AD” [42]. Based on this latter, excessive mitochondrial Ca2+ uptake triggers an increase in ROS production, ATP synthesis inhibition, mPTP opening, release of cytochrome c, activation of caspases and apoptosis (revised in [43], Figure 1B). However, contrasting data have also been produced, and this scheme is not completely accepted. Nowadays, there are still few works investigating the role of mitochondria in the pathogenesis of AD in physiologically relevant experimental samples, i.e., ex vivo and in vivo. Furthermore, one of the main issues to be addressed for both the Ca2+ and mitochondrial cascade hypotheses is to define whether Ca2+ or mitochondrial dysfunctions have a primary and causative role in AD pathogenesis. Indeed, to gain insight into new potential therapeutic targets, it is crucial to determine whether one of these defects is sufficient per se to cause AD. Since the mitochondrial ability to take up Ca2+ strongly depends on cytosolic Ca2+ dynamics, both parameters must be assessed in the same model. Indeed, many global Ca2+ alterations have been reported so far in various AD models.
4. Mitochondria Ca2+ Signaling and Bioenergetics as Therapeutic Targets
Since mitochondrial Ca2+ signaling is pivotal in determining many brain cell functions, its therapeutic targeting can be exploited to avoid neurodegeneration and cell death. The recent discovery of the molecular components of MCUC expands the possibility to tune mitochondrial Ca2+ handling. Nowadays, many drugs targeting MCUC are available, but most of them are not able to cross the blood–brain barrier or they are prone to multiple side effects. Concerning mitochondrial Ca2+ uptake, the historical inhibitor and cell-impermeant ruthenium red have been modified, obtaining a more permeable and powerful version called Ru265 [44]. High-throughput screening analyses are unveiling novel inhibitors of MCUC, such as Mitoxantrone [45], DS16570511 [46], MCU-i4 and MCU-i11 [47]. Recently, amorolfine and benzethonium, positive and negative MCUC modulators, respectively, have been identified by high-throughput screening on a US Food and Drug Administration (FDA)-approved drug library [48].
Natural plant flavonoids have been reported to activate MCU. Kaempferol is one of the compounds reported to exert a protective effect on mitochondrial functions by activating MCU [49]. Furthermore, kaempferol has been shown to improve the metabolism-secretion coupling of β-pancreatic cells [50]. SB202190, an inhibitor of p38 mitogen-activated protein (MAP) kinase, has also been reported to reversibly stimulate mitochondrial Ca2+ uptake [49]. Antioxidants have been reported to positively modulate mitochondrial Ca2+ homeostasis, by stabilizing the cellular redox state or targeting mitochondrial ROS (reviewed in [51]).
As for mitochondrial Ca2+ efflux, the NCLX inhibitor CGP-37157 is widely used despite its lack of specificity [52].
Furthermore, different therapeutic possibilities are expanding over time, showing promising tools for modulating mitochondrial function (reviewed here [53]). Gene therapy can be exploited to manipulate specific genes expressed only in a cell type, targeting either neurons, astrocytes or microglia, as well as to specifically edit mitochondrial DNA (mtDNA) without altering genomic DNA. Finally, diet and exercise can enhance/modulate mitochondrial function. As an example, ketogenic diets have been reported to rescue neuronal Ca2+ handling dysfunction, by overcoming the mitochondria hypometabolism caused by impaired pyruvate import [54].
References
- Sokoloff, L. Energetics of functional activation in neural tissues. Neurochem. Res. 1999, 24, 321–329.
- Giorgi, C.; Missiroli, S.; Patergnani, S.; Duszynski, J.; Wieckowski, M.R.; Pinton, P. Mitochondria-associated membranes: Composition, molecular mechanisms, and physiopathological implications. Antioxid. Redox Signal. 2015, 22, 995–1019.
- Filadi, R.; Greotti, E.; Pizzo, P. Highlighting the endoplasmic reticulum-mitochondria connection: Focus on Mitofusin 2. Pharmacol. Res. 2018, 128, 42–51.
- Alzheimer’s Association. 2020 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2020, 16, 391–460.
- Niu, H.; Álvarez-Álvarez, I.; Guillén-Grima, F.; Aguinaga-Ontoso, I. Prevalence and incidence of Alzheimer’s disease in Europe: A meta-analysis. Neurologia 2017, 32, 523–532.
- Cuyvers, E.; Sleegers, K. Genetic variations underlying Alzheimer’s disease: Evidence from genome-wide association studies and beyond. Lancet Neurol. 2016, 15, 857–868.
- Altomari, N.; Bruno, F.; Laganà, V.; Smirne, N.; Colao, R.; Curcio, S.; Di Lorenzo, R.; Frangipane, F.; Maletta, R.; Puccio, G.; et al. A Comparison of Behavioral and Psychological Symptoms of Dementia (BPSD) and BPSD Sub-Syndromes in Early-Onset and Late-Onset Alzheimer’s Disease. J. Alzheimer’s Dis. 2022, 85, 691–699.
- Scheltens, P.; Blennow, K.; Breteler, M.M.B.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. Lancet 2016, 388, 505–517.
- Hardy, J.A.; Higgins, G.A. Alzheimer’s disease: The amyloid cascade hypothesis. Science 1992, 256, 184–185.
- Kosik, K.S.; Joachim, C.L.; Selkoe, D.J. Microtubule-associated protein tau (tau) is a major antigenic component of paired helical filaments in Alzheimer disease. Proc. Natl. Acad. Sci. USA 1986, 83, 4044–4048.
- Khan, A.; Corbett, A.; Ballard, C. Emerging treatments for Alzheimer’s disease for non-amyloid and non-tau targets. Expert Rev. Neurother. 2017, 17, 683–695.
- Poudel, P.; Park, S. Recent Advances in the Treatment of Alzheimer’s Disease Using Nanoparticle-Based Drug Delivery Systems. Pharmaceutics 2022, 14, 835.
- Du, X.; Wang, X.; Geng, M. Alzheimer’s disease hypothesis and related therapies. Transl. Neurodegener. 2018, 7, 2.
- Bruno, F.; Malvaso, A.; Canterini, S.; Bruni, A.C. Antimicrobial Peptides (AMPs) in the Pathogenesis of Alzheimer’s Disease: Implications for Diagnosis and Treatment. Antibiotics 2022, 11, 726.
- Bell, S.M.; Barnes, K.; De Marco, M.; Shaw, P.J.; Ferraiuolo, L.; Blackburn, D.J.; Venneri, A.; Mortiboys, H. Mitochondrial Dysfunction in Alzheimer’s Disease: A Biomarker of the Future? Biomedicines 2021, 9, 63.
- Weidling, I.W.; Swerdlow, R.H. Mitochondria in Alzheimer’s disease and their potential role in Alzheimer’s proteostasis. Exp. Neurol. 2020, 330, 113321.
- Callens, M.; Loncke, J.; Bultynck, G. Dysregulated Ca2+ Homeostasis as a Central Theme in Neurodegeneration: Lessons from Alzheimer’s Disease and Wolfram Syndrome. Cells 2022, 11, 1963.
- Santello, M.; Toni, N.; Volterra, A. Astrocyte function from information processing to cognition and cognitive impairment. Nat. Neurosci. 2019, 22, 154–166.
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite synapses: Glia, the unacknowledged partner. Trends Neurosci. 1999, 22, 208–215.
- Schafer, D.P.; Lehrman, E.K.; Stevens, B. The “quad-partite” synapse: Microglia-synapse interactions in the developing and mature CNS. Glia 2013, 61, 24–36.
- Kunkle, B.W.; Grenier-Boley, B.; Sims, R.; Bis, J.C.; Damotte, V.; Naj, A.C.; Boland, A.; Vronskaya, M.; van der Lee, S.J.; Amlie-Wolf, A.; et al. Genetic meta-analysis of diagnosed Alzheimer’s disease identifies new risk loci and implicates Aβ, tau, immunity and lipid processing. Nat. Genet. 2019, 51, 414–430.
- Gatz, M.; Reynolds, C.A.; Fratiglioni, L.; Johansson, B.; Mortimer, J.A.; Berg, S.; Fiske, A.; Pedersen, N.L. Role of genes and environments for explaining Alzheimer disease. Arch. Gen. Psychiatry 2006, 63, 168–174.
- Zhang, Q.; Sidorenko, J.; Couvy-Duchesne, B.; Marioni, R.E.; Wright, M.J.; Goate, A.M.; Marcora, E.; Huang, K.L.; Porter, T.; Laws, S.M.; et al. Risk prediction of late-onset Alzheimer’s disease implies an oligogenic architecture. Nat. Commun. 2020, 11, 4799.
- Wightman, D.P.; Jansen, I.E.; Savage, J.E.; Shadrin, A.A.; Bahrami, S.; Holland, D.; Rongve, A.; Børte, S.; Winsvold, B.S.; Drange, O.K.; et al. A genome-wide association study with 1,126,563 individuals identifies new risk loci for Alzheimer’s disease. Nat. Genet. 2021, 53, 1276–1282.
- Wang, W.; Zhao, F.; Ma, X.; Perry, G.; Zhu, X. Mitochondria dysfunction in the pathogenesis of Alzheimer’s disease: Recent advances. Mol. Neurodegener. 2020, 15, 30.
- Cavalcante, G.C.; Brito, L.M.; Schaan, A.P.; Ribeiro-Dos-santos, Â.; de Araújo, G.S. Mitochondrial Genetics Reinforces Multiple Layers of Interaction in Alzheimer’s Disease. Biomedicines 2022, 10, 880.
- Flaquer, A.; Baumbach, C.; Kriebel, J.; Meitinger, T.; Peters, A.; Waldenberger, M.; Grallert, H.; Strauch, K. Mitochondrial genetic variants identified to be associated with BMI in adults. PLoS ONE 2014, 9, e105116.
- Rumshisky, A.; Ghassemi, M.; Naumann, T.; Szolovits, P.; Castro, V.M.; McCoy, T.H.; Perlis, R.H. Predicting early psychiatric readmission with natural language processing of narrative discharge summaries. Transl. Psychiatry 2016, 6, e921.
- Schröder, H.; Moser, N.; Huggenberger, S. Neuroanatomy of the Mouse; Springer International Publishing: Cham, Switzerland, 2020.
- de Rus Jacquet, A.; Denis, H.L.; Cicchetti, F.; Alpaugh, M. Current and future applications of induced pluripotent stem cell-based models to study pathological proteins in neurodegenerative disorders. Mol. Psychiatry 2021, 26, 2685–2706.
- Wai, T.; Langer, T. Mitochondrial Dynamics and Metabolic Regulation. Trends Endocrinol. Metab. 2016, 27, 105–117.
- Messina, A.; Reina, S.; Guarino, F.; De Pinto, V. VDAC isoforms in mammals. Biochim. Biophys. Acta 2012, 1818, 1466–1476.
- Perrone, M.; Caroccia, N.; Genovese, I.; Missiroli, S.; Modesti, L.; Pedriali, G.; Vezzani, B.; Vitto, V.A.M.; Antenori, M.; Lebiedzinska-Arciszewska, M.; et al. The role of mitochondria-associated membranes in cellular homeostasis and diseases. In International Review of Cell and Molecular Biology; Elsevier Publighing: Amsterdam, The Netherlands, 2020; ISBN 9780128197448.
- Hirabayashi, Y.; Kwon, S.-K.; Paek, H.; Pernice, W.M.; Paul, M.A.; Lee, J.; Erfani, P.; Raczkowski, A.; Petrey, D.S.; Pon, L.A.; et al. ER-mitochondria tethering by PDZD8 regulates Ca2+ dynamics in mammalian neurons. Science 2017, 358, 623–630.
- Hayashi, T.; Su, T.P. Sigma-1 receptor chaperones at the ER-mitochondrion interface regulate Ca(2+) signaling and cell survival. Cell 2007, 131, 596–610.
- Ooi, K.; Hu, L.; Feng, Y.; Han, C.; Ren, X.; Qian, X.; Huang, H.; Chen, S.; Shi, Q.; Lin, H.; et al. Sigma-1 Receptor Activation Suppresses Microglia M1 Polarization via Regulating Endoplasmic Reticulum–Mitochondria Contact and Mitochondrial Functions in Stress-Induced Hypertension Rats. Mol. Neurobiol. 2021, 58, 6625–6646.
- Göbel, J.; Engelhardt, E.; Pelzer, P.; Sakthivelu, V.; Jahn, H.M.; Jevtic, M.; Folz-Donahue, K.; Kukat, C.; Schauss, A.; Frese, C.K.; et al. Mitochondria-Endoplasmic Reticulum Contacts in Reactive Astrocytes Promote Vascular Remodeling. Cell Metab. 2020, 31, 791–808.e8.
- Khachaturian, Z.S. Hypothesis on the regulation of cytosol calcium concentration and the aging brain. Neurobiol. Aging 1987, 8, 345–346.
- Khachaturian, Z.S. Calcium, membranes, aging, and Alzheimer’s disease. Introduction and overview. Ann. N. Y. Acad. Sci. 1989, 568, 1–4.
- LaFerla, F.M. Calcium dyshomeostasis and intracellular signalling in Alzheimer’s disease. Nat. Rev. Neurosci. 2002, 3, 862–872.
- Landfield, P.W.; Pitler, T.A. Prolonged Ca2+-dependent afterhyperpolarizations in hippocampal neurons of aged rats. Science 1984, 226, 1089–1092.
- Swerdlow, R.H.; Khan, S.M. A “mitochondrial cascade hypothesis” for sporadic Alzheimer’s disease. Med. Hypotheses 2004, 63, 8–20.
- Filadi, R.; Greotti, E. The yin and yang of mitochondrial Ca2+ signaling in cell physiology and pathology. Cell Calcium 2021, 93, 102321.
- Woods, J.J.; Nemani, N.; Shanmughapriya, S.; Kumar, A.; Zhang, M.; Nathan, S.R.; Thomas, M.; Carvalho, E.; Ramachandran, K.; Srikantan, S.; et al. A Selective and Cell-Permeable Mitochondrial Calcium Uniporter (MCU) Inhibitor Preserves Mitochondrial Bioenergetics after Hypoxia/Reoxygenation Injury. ACS Cent. Sci. 2019, 5, 153–166.
- Arduino, D.M.; Wettmarshausen, J.; Vais, H.; Navas-Navarro, P.; Cheng, Y.; Leimpek, A.; Ma, Z.; Delrio-Lorenzo, A.; Giordano, A.; Garcia-Perez, C.; et al. Systematic Identification of MCU Modulators by Orthogonal Interspecies Chemical Screening. Mol. Cell 2017, 67, 711–723 e7.
- Kon, N.; Murakoshi, M.; Isobe, A.; Kagechika, K.; Miyoshi, N.; Nagayama, T. DS16570511 is a small-molecule inhibitor of the mitochondrial calcium uniporter. Cell Death Discov. 2017, 3, 17045.
- Di Marco, G.; Vallese, F.; Jourde, B.; Bergsdorf, C.; Sturlese, M.; De Mario, A.; Techer-Etienne, V.; Haasen, D.; Oberhauser, B.; Schleeger, S.; et al. A High-Throughput Screening Identifies MICU1 Targeting Compounds. Cell Rep. 2020, 30, 2321–2331.e6.
- De Mario, A.; Tosatto, A.; Hill, J.M.; Kriston-Vizi, J.; Ketteler, R.; Vecellio Reane, D.; Cortopassi, G.; Szabadkai, G.; Rizzuto, R.; Mammucari, C. Identification and functional validation of FDA-approved positive and negative modulators of the mitochondrial calcium uniporter. Cell Rep. 2021, 35, 109275.
- Montero, M.; Lobaton, C.D.; Hernandez-Sanmiguel, E.; Santodomingo, J.; Vay, L.; Moreno, A.; Alvarez, J. Direct activation of the mitochondrial calcium uniporter by natural plant flavonoids. Biochem. J. 2004, 384, 19–24.
- Bermont, F.; Hermant, A.; Benninga, R.; Chabert, C.; Jacot, G.; Santo-Domingo, J.; Kraus, M.R.C.; Feige, J.N.; De Marchi, U. Targeting Mitochondrial Calcium Uptake with the Natural Flavonol Kaempferol, to Promote Metabolism/Secretion Coupling in Pancreatic β-cells. Nutrients 2020, 12, 538.
- Rodríguez, L.R.; Lapeña-Luzón, T.; Benetó, N.; Beltran-Beltran, V.; Pallardó, F.V.; Gonzalez-Cabo, P.; Navarro, J.A. Therapeutic Strategies Targeting Mitochondrial Calcium Signaling: A New Hope for Neurological Diseases? Antioxidants 2022, 11, 165.
- Neumann, J.T.; Diaz-Sylvester, P.L.; Fleischer, S.; Copello, J.A. CGP-37157 inhibits the sarcoplasmic reticulum Ca2+ ATPase and activates ryanodine receptor channels in striated muscle. Mol. Pharmacol. 2011, 79, 141–147.
- Onyango, I.G. Modulation of mitochondrial bioenergetics as a therapeutic strategy in Alzheimer’s disease. Neural Regen. Res. 2018, 13, 19–25.
- De La Rossa, A.; Laporte, M.H.; Astori, S.; Marissal, T.; Montessuit, S.; Sheshadri, P.; Ramos-Fernández, E.; Mendez, P.; Khani, A.; Quairiaux, C.; et al. Paradoxical neuronal hyperexcitability in a mouse model of mitochondrial pyruvate import deficiency. Elife 2022, 11, e72595.
More
Information
Subjects:
Cell Biology; Physiology; Pathology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.2K
Revisions:
2 times
(View History)
Update Date:
09 Dec 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No