 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Mark Eiteman | -- | 3266 | 2022-11-17 23:41:06 | | | |
| 2 | Lindsay Dong | Meta information modification | 3266 | 2022-11-21 02:20:49 | | |
Video Upload Options
Limiting an essential nutrient has a profound impact on microbial growth. The notion of growth under limited conditions was first described using simple Monod kinetics proposed in the 1940s. Different operational modes (chemostat, fed-batch processes) were soon developed to address questions related to microbial physiology and cell maintenance and to enhance product formation. With more recent developments of metabolic engineering and systems biology, as well as high-throughput approaches, the focus of current engineers and applied microbiologists has shifted from these fundamental biochemical processes.
1. Introduction
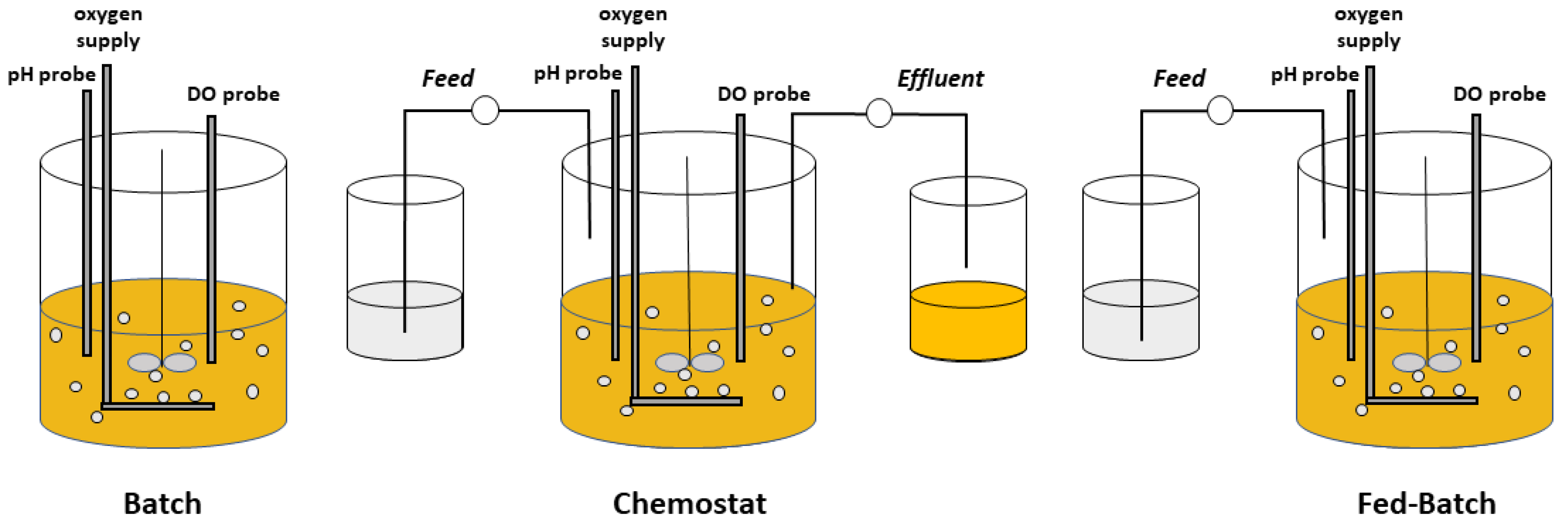
2. Chemostat Processes
3. Fed-Batch Processes
4. Nutrient Limitation Compared to Nutrient Starvation
5. Physiological Effects of Non-Carbon Nutrient Limitation
5.1. Nitrogen
Nitrogen occurs throughout cells, in proteins, nucleotides, and many metabolites. Nitrogen as ammonium is assimilated in E. coli and most bacteria and yeast into glutamine and glutamate [49], which are the primary intracellular nitrogen donors. In bacteria, 88% of the cellular nitrogen is derived from glutamate, while 12% is derived from glutamine [50]. Glutamate is the most abundant metabolite in E. coli, accounting for about 40% of the total metabolite concentration [51]. In many bacteria such as E. coli, two ammonium-assimilating pathways are available, a NADPH-dependent glutamate dehydrogenase and a high-affinity glutamate synthase (glutamine oxoglutarate aminotransferase, GOGAT)/glutamine synthase. Glutamate dehydrogenases generally have high values of KM for ammonium, so that during N-limited growth, glutamine synthase expression is elevated to maintain sufficient glutamate [52]. Dynamic N starvation in E. coli and S. cerevisiae growing on glucose depletes glutamine and to a lesser extent glutamate while α-ketoglutarate increases markedly and can even be excreted [47][53]. Accumulation of α-ketoglutarate occurs in cyanobacteria also, a signal which upregulates nitrogen assimilation via the global regulator NtcA and other regulators [54][55]. In yeast, the concentration of tryptophan, which relies on glutamine for its synthesis, decreases, while phenylalanine and tyrosine, which rely on glutamate for nitrogen, do not change, resulting in an accumulation of phenylpyruvate and phenylethanol, a quorum-sensing signal [56]. Accumulated α-ketoglutarate in E. coli noncompetitively and cooperatively inhibits EI of the PTS [57], citrate synthase [58], and PEP synthetase [59]. Furthermore, sudden nitrogen availability in N-starved wild-type E. coli induces a decrease in α-ketoglutarate and rapid increase in glucose uptake rate, while in a PTS-deficient strain with elevated galactose permease, glucose uptake is insensitive to N-availability [57]. In general, S. cerevisiae N-limitation leads to depletion of intracellular amino acids, particularly at low dilution rates [53]. Because intracellular glutamine and arginine concentration correlates strongly with dilution rate, these compounds likely control growth in N-limited S. cerevisiae [53]. Synechocystis also accumulate α-ketoglutarate [60] under N-starvation as well as glycogen, which is associated with the induction of the glgX gene [61][62][63]. E. coli has a high protein turnover under N-limited conditions compared to C-limited or P-limited conditions [64].
5.2. Phosphorus
5.3. Sulfur
Sulfur is present in the amino acids methionine and cysteine, and important metabolites such as S-adenosylmethionine, Coenzyme A (CoA), and lipoic acid. In most media, S is supplied as the sulfate ion, and thus, S-limitation is often studied specifically as sulfate-limitation.
5.4. Magnesium
5.5. Iron
Consistent with the theme of cells conserving limited resources, Fe-limitation causes cells to reduce their reliance on pathways which contain significant Fe, such as the tricarboxylic acid cycle (aconitase, fumarate hydratase, succinate dehydrogenase) and the proton-pumping components of the electron transport chain [101][102][103]. This realignment of metabolism has physiological consequences. For example, under Fe-limiting aerobic conditions, E. coli generates acetate at a yield of 0.25 g/g at a dilution rate of 0.4 h−1, but accumulates predominantly lactate (yield of 0.60 g/g) at 0.1 h−1 [101]. Elevated lactate under the most severe steady-state Fe-limitation provides cells a means to oxidize NADH when the Fe-requiring Nuo complex is curtailed, while an increased glucose uptake rate serves to meet ATP demand [101]. Lactate formation is also observed for Staphylococcus aureus exposed to Fe-starvation [104]. The reduction in activity of TCA cycle enzymes and accumulation of NADH encourages the formation of ethyl acetate by Kluyveromyces marxianus [105] and Candida utilis [106]. Because of the restricted capacity of the electron transport chain subject to Fe-limited conditions, cells essentially behave like they are encountering anaerobic conditions. For example, E. coli accumulates acetate and formate under Fe-limited steady-state conditions, attains a 60% greater glycolytic flux, and reduces by a factor of 5 the fraction of glucose entering the TCA cycle [107]. More generally, Fe-limitation impacts oxygenation and redox state because numerous electron-carrying enzymes contain Fe-S clusters, and many Fe-containing enzymes or pathways are constrained under Fe-limitation. For example, the nitrogenase enzyme system contains significant Fe [108], and Fe-limitation thus severely reduces nitrogen fixation by Azotobacter vinelandii [109]. Notably, lactic acid bacteria, which lack cytochrome and show generally high tolerance to peroxide, do not require iron for growth at all [110][111].
References
- Gaden, E.L., Jr. Bioengineering and fermentation. Appl. Microbiol. 1960, 8, 123–131.
- Maxon, W.D. Continuous fermentation: A discussion of its principles and applications. Appl. Microbiol. 1955, 3, 110–120.
- Yoshida, F.; Yamane, T.; Nakamoto, K.-I. Fed-batch hydrocarbon fermentation with colloidal emulsion feed. Biotechnol. Bioeng. 1973, 15, 257–270.
- Pirt, S.J. The theory of fed batch culture with reference to the penicillin fermentation. J. Appl. Chem. Biotechnol. 1974, 24, 415–424.
- Fox, M.S. A device for growing bacterial populations under steady state conditions. J. Gen. Physiol. 1955, 39, 261–266.
- Maxon, W.D. Continuous fermentation. Ind. Eng. Chem. 1960, 52, 64–65.
- Paalme, T.; Kahru, A.; Elken, R.; Vanatalu, K.; Tiisma, K.; Vilu, R. The computer-controlled continuous culture of Escherichia coli with a smooth change in dilution rate (A-stat). J. Microbiol. Meth. 1995, 24, 145–153.
- Schmideder, A.; Weuster-Botz, D. High-performance recombinant protein production with Escherichia coli in continuously operated cascades of stirred-tank reactors. J. Ind. Microbiol. Biotechnol. 2017, 44, 1021–1029.
- Adamberg, K.; Valgepea, K.; Vilu, R. Advanced continuous cultivation methods for systems microbiology. Microbiology 2015, 161, 1707–1719.
- Novick, A.; Szilard, L. Experiments with the chemostat on spontaneous mutations on bacteria. Proc. Natl. Acad. Sci. USA 1950, 36, 708–719.
- Ziv, N.; Brandt, N.J.; Gresham, D. The Use of Chemostats in Microbial Systems Biology. J. Vis. Exp. 2013, 80, e50168.
- Lameiras, F.; Heijnen, J.J.; van Gulik, W.M. Development of tools for quantitative intracellular metabolomics of Aspergillus niger chemostat cultures. Metabolomics 2015, 11, 1253–1264.
- Egli, T. Microbial growth and physiology: A call for better craftsmanship. Front. Microbiol. 2015, 6, 287.
- Herbert, D.; Elsworth, R.; Telling, R.C. The continuous culture of bacteria; a theoretical and experimental study. J. Gen. Microbiol. 1956, 14, 601–622.
- Powell, E.O. Criteria for the growth of contaminants and mutants in continuous culture. J. Gen. Microbiol. 1958, 18, 259–268.
- Rosenzweig, R.F.; Sharp, R.R.; Treves, D.S.; Adams, J. Microbial evolution in a simple unstructured environment: Genetic differentiation in Escherichia coli. Genetics 1994, 137, 903–917.
- Weikert, C.; Sauer, U.; Bailey, J.E. Use of a glycerol-limited, long-term chemostat for isolation of Escherichia coli mutants with improved physiological properties. Microbiology 1997, 143, 1567–1574.
- Bull, A.T. The renaissance of continuous culture in the post-genomics age. J. Ind. Microbiol. Biotechnol. 2010, 37, 993–1021.
- Wick, L.M.; Weilenmann, H.; Egli, T. The apparent clock-like evolution of Escherichia coli in glucose-limited chemostats is reproducible at large but not at small population sizes and can be explained with Monod kinetics. Microbiology 2002, 148, 2889–2902.
- Jansen, M.L.A.; Diderich, J.A.; Mashego, M.; Hassane, A.; de Winde, J.H.; Pascale, D.-L.; Pronk, J.T. Prolonged selection in aerobic, glucose-limited chemostat cultures of Saccharomyces cerevisiae causes a partial loss of glycolytic capacity. Microbiology 2005, 151, 1657–1669.
- Gresham, D.; Hong, J. The functional basis of adaptive evolution in chemostats. FEMS Microbiol. Rev. 2015, 39, 2–16.
- Brickwedde, A.; van den Broek, M.; Geertman, J.-M.A.; Magalhães, F.; Kuijpers, N.G.A.; Gibson, B.; Pronk, J.T.; Daran, J.-M.G. Evolutional engineering in chemostat cultures for improved maltotriose fermentation kinetics in Saccharomyces pastorianus lager brewing yeast. Front. Microbiol. 2017, 8, 1690.
- van Maris, A.J.A.; Geertman, J.-M.A.; Vermeulen, A.; Groothuizen, M.K.; Winkler, A.A.; Piper, M.D.W.; van Dijken, J.P.; Pronk, J.T. Directed evolution of pyruvate decarboxylase-negative Saccharomyces cerevisiae, yielding a C2-independent, glucose-tolerant, and pyruvate-hyperproducing yeast. Appl. Environ. Microbiol. 2004, 70, 159–166.
- Arensdorf, J.J.; Loomis, A.K.; DiGarazia, P.M.; Monticello, D.J.; Piendos, P.T. Chemostat approach for the directed evolution of biodesulfurization gain-of-function mutants. Appl. Environ. Microbiol. 2002, 68, 691–698.
- Maharjan, R.P.; Ferenci, T.A. shifting mutational landscape in 6 nutritional states: Stress-induced mutagenesis as a series of distinct stress input-mutation output relationships. PLoS Biol. 2017, 15, e2001477.
- Schreiber, F.; Littmann, S.; Lavik, G.; Escrig, S.; Meibom, A.; Kuypers, M.M.M.; Ackermann, M. Phenotypic heterogeneity driven by nutrient limitation promotes growth in fluctuating environments. Nat. Microbiol. 2016, 1, 16055.
- Cooney, C.L.; Wang, D.I. Transient response of Enterobacter aerogenes under a dual nutrient limitation in a chemostat. Biotechnol. Bioeng. 1978, 18, 189–198.
- Egli, T. On multiple-nutrient-limited growth of microorganisms, with special reference to dual limitation by carbon and nitrogen substrates. Antonie Van Leeuwenhoek 1991, 60, 225–234.
- Egli, T.; Zinn, M. The concept of multiple-nutrient-limited growth of microorganisms and its application in biotechnological processes. Biotechnol. Adv. 2003, 22, 35–43.
- Zinn, M.; Witholt, B.; Egli, T. Dual nutrient limited growth: Models, experimental observations and applications. J. Biotechnol. 2004, 113, 263–279.
- Lis, A.V.; Schneider, K.; Weber, J.; Keasling, J.D.; Jensen, M.K.; Klein, T. Exploring small-scale chemostats to scale up microbial processes: 3-hydroxypropionic acid production in S. cerevisiae. Microb. Cell Factories 2019, 18, 50.
- Yamanè, T.; Shimizu, S. Fed-batch techniques in microbial processes. In Bioprocess Parameter Control Advances in Biochemical Engineering/Biotechnology; Springer: Berlin/Heidelberg, Germany, 1984; Volume 30, pp. 147–194.
- Trilli, A.; Michelini, V.; Mantovani, V.; Pirt, S.J. Estimation of productivities in repeated fed-batch cephalosporin fermentation. J. Appl. Chem. Biotechnol. 1977, 27, 219–224.
- Yano, T.; Kurokawa, M.; Nishizawa, Y. Optimum substrate feed rate in fed-batch culture with the DO-stat method. J. Fermentation Bioeng. 1991, 71, 345–349.
- Hu, Z.-C.; Zheng, Y.-G.; Shen, Y.-C. Dissolved-oxygen-stat fed-batch fermentation of 1,3-dihydroxyacetone from glycerol by Gluconobacter oxydans ZJB09112. Biotechnol. Bioprocess Eng. 2010, 15, 651–656.
- Jørgensen, H.; Olsson, L.; Rønnow, B.; Palmqvist, E.A. Fed-batch cultivation of baker’s yeast followed by nitrogen or carbon starvation: Effects on fermentative capacity and content of trehalose and glycogen. Appl. Microbiol. Biotechnol. 2002, 59, 310–317.
- Shang, L.; Jiang, M.; Chang, H.N. Poly(3-hydroxybutyrate) synthesis in fed-batch culture of Ralstonia eutropha with phosphate limitation under different glucose concentrations. Biotechnol. Lett. 2003, 25, 1415–1419.
- Shen, X.-F.; Liu, J.-J.; Chauhan, A.S.; Hu, H.; Ma, L.-L.; Lam, P.K.S.; Zeng, R.J. Combining nitrogen starvation with sufficient phosphorus supply for enhanced biodiesel productivity of Chlorella vulgaris fed on acetate. Algal Res. 2016, 17, 261–267.
- Kim, H.M.; Chae, T.U.; Choi, S.Y.; Kim, W.J.; Lee, S.Y. Engineering of an oleaginous bacterium for the production of fatty acids and fuels. Nat. Chem. Biol. 2019, 15, 721–729.
- Nayak, M.; Suh, W.I.; Chang, Y.K.; Lee, B. Exploration of two-stage cultivation strategies using nitrogen starvation to maximize the lipid productivity in Chlorella sp. HS2. Bioresour. Technol. 2019, 276, 110–118.
- Pirt, S.J. Fed-batch culture of microbes. Ann. N. Y. Acad. Sci. 1979, 326, 119–125.
- Dunn, I.J.; Mor, J.-R. Variable-volume continuous cultivation. Biotechnol. Bioeng. 1975, 17, 1805–1822.
- Förberg, C.; Häggström, L. Effects of cultural conditions on the production of phenylalanine from a plasmid-harboring E. coli strain. Appl. Microbiol. Biotechnol. 1987, 26, 136–140.
- Werner-Washburne, M.; Braun, E.L.; Crawford, M.E.; Peck, V.M. Stationary phase in Saccharomyces cerevisiae. Mol. Microbiol. 1996, 19, 1159–1166.
- Gasch, A.P.; Werner-Washburne, M. The genomics of yeast responses to environmental stress and starvation. Funct. Integr. Genom. 2002, 2, 181–192.
- Peterson, C.N.; Mandel, M.J.; Silhavy, T.J. Escherichia coli starvation diets: Essential nutrients weigh in distinctly. J. Bacteriol. 2005, 187, 7549–7553.
- Brauner, M.J.; Yuan, J.; Bennett, B.D.; Lu, W.; Kimball, E.; Botstein, D.; Rabinowitz, J.D. Conservation of the metabolomic response to starvation across two divergent microbes. Proc. Natl. Acad. Sci. USA 2006, 103, 19302–19307.
- Minihane, B.J.; Brown, D.E. Fed-batch culture technology. Biotechnol. Adv. 1986, 4, 207–218.
- Reitzer, L. Nitrogen assimilation and global regulation in Escherichia coli. Annu. Rev. Microbiol. 2003, 57, 155–176.
- Hu, P.; Leighton, T.; Ishkhanova, G.; Kustu, S. Sensing of nitrogen limitation by Bacillus subtilis: Comparison to enteric bacteria. J. Bacteriol. 1999, 181, 5042–5050.
- Bennett, B.D.; Kimball, E.H.; Gao, M.; Osterhout, R.; van Dien, S.J.; Rabinowitz, J.D. Absolute metabolite concentrations and implied enzyme active site occupancy in Escherichia coli. Nat. Chem. Biol. 2009, 5, 593–599.
- Reitzer, L.J. Ammonia assimilation and the biosynthesis of glutamine, glutamate, aspartate, asparagine, L-alanine, and D-alanine. In Escherichia coli and Salmonella: Cellular and Molecular Biology; Neidhardt, F.C., Curtiss, R., III, Ingraham, J.L., Lin, E.C.C., Low, K.B., Magasanik, B., Eds.; American Society for Microbiology Press: Washington, DC, USA, 1996; pp. 391–407.
- Boer, V.M.; Crutchfield, C.A.; Bradley, P.H.; Botstein, D.; Rabinowitz, J.D. Growth-limiting intracellular metabolites in yeast growing under diverse nutrient limitations. Mol. Biol. Cell. 2010, 21, 198–211.
- Muro-Pastor, M.I.; Reyes, J.C.; Florencio, F.J. Ammonium assimilation in cyanobacteria. Photosynth. Res. 2005, 83, 135–150.
- Espinosa, J.; Rodriguez-Mateos, F.; Salinas, P.; Lanza, V.F.; Dixon, R.; de la Cruz, F.; Contreras, A. PipX, the coactivator of NtcA, is a global regulator in cyanobacteria. Proc. Natl. Acad. Sci. USA 2014, 111, e2423–e2430.
- Chen, H.; Fink, G.R. Feedback control of morphogenesis in fungi by aromatic alcohols. Genes Devel. 2006, 20, 1150–1161.
- Doucette, C.D.; Schwab, D.J.; Wingreen, N.S.; Rabinowitz, J.D. α-ketoglutarate coordinates carbon and nitrogen utilization via enzyme I inhibition. Nat. Chem. Biol. 2011, 7, 894–901.
- Pereira, D.S.; Donald, L.J.; Hosfield, D.J.; Duckworth, H.W. Active site mutants of Escherichia coli citrate synthase. J. Biol. Chem. 1994, 269, 412–417.
- Chulavatnatol, M.; Atkinson, D.E. Phosphoenolpyruvate synthetase from Escherichia coli: Effects of adenylate energy charge and modifier concentrations. J. Biol. Chem. 1973, 248, 2712–2715.
- Muro-Pastor, M.I.; Reyes, J.C.; Florencio, F.J. Cyanobacteria perceive nitrogen status by sensing intracellular 2-oxoglutarate levels. J. Biol. Chem. 2001, 276, 38320–38328.
- De Marsac, N.T.; Castets, A.-M.; Cohen-Bazire, G. Wavelength modulation of phycoerythrin synthesis in Synechocystis sp. 6701. J. Bacteriol. 1980, 142, 310–314.
- Osanai, T.; Imamura, S.; Asayama, M.; Shirai, M.; Suzuki, I.; Murata, N.; Tanaka, K. Nitrogen induction of sugar catabolic gene expression in Synechocystis sp. PCC 6803. DNA Res. 2006, 13, 185–195.
- Yoo, S.-H.; Keppel, C.; Spalding, M.; Jane, J.-L. Effects of growth condition on the structure of glycogen produced in cyanobacterium Synechocystis sp. PCC6803. Intl. J. Biol. Mol. 2007, 40, 498–504.
- Gupta, M.; Johnson, A.; Cruz, E.; Costa, E.; Guest, R.; Li, S.H.-J.; Hart, E.M.; Nguyen, T.; Stadlmeier, M.; Bratton, B.P.; et al. Global protein-turnover quantification in Escherichia coli reveals cytoplasmic recycling under nitrogen-limitation. bioRxiv 2022.
- Devine, K.M. Activation of the PhoPR-mediated response to phosphate limitation is regulated by wall teichoic acid metabolism in Bacillus subtilis. Front. Microbiol. 2018, 9, 2678.
- Ellwood, D.C.; Tempest, D.W. Teichoic acid or teichuronic acid in the walls of Bacillus subtilis var. niger, grown in a chemostat. Biochem. J. 1967, 104, 69.
- Ellwood, D.C.; Tempest, D.W. Control of teichoic acid and teichuronic acid biosynthesis in chemostat cultures of Bacillus subtilis var. niger. Biochem. J. 1969, 111, 1–5.
- Anderson, A.J.; Green, R.S.; Archibald, A.R. Wall composition and phage-binding properties of Bacillus subtilis W23 grown in chemostat culture in media containing varied concentrations of phosphate. FEMS Microbiol. Lett. 1978, 4, 129–132.
- Lang, W.K.; Glassey, K.; Archibald, A.R. Influence of phosphate supply on teichoic acid and teichuronic acid content of Bacillus subtilis cell walls. J. Bacteriol. 1982, 151, 367–375.
- Wouters, J.T.M.; Buysman, P.J. Production of some exocellular enzymes by Bacillus licheniformis 749/C in chemostat cultures. FEMS Microbiol. Lett. 1977, 1, 109–112.
- Boer, V.M.; de Winde, J.H.; Pronk, J.T.; Piper, M.D. The genome-wide transcriptional responses of Saccharomyces cerevisiae grown on glucose in aerobic chemostat cultures limited for carbon, nitrogen, phosphorous, or sulfur. J. Biol. Chem. 2003, 278, 3265–3274.
- Li, S.H.-J.; Li, Z.; Park, J.O.; King, C.G.; Rabinowitz, J.D.; Wingreen, N.S.; Gitai, Z. Escherichia coli translation strategies differ across carbon, nitrogen and phosphorous limitation conditions. Nat. Microbiol. 2018, 3, 939–947.
- Alton, T.H.; Koch, A.L. Unused protein synthetic capacity of Escherichia coli grown in phosphate-limited chemostats. J. Mol. Biol. 1974, 86, 1–9.
- Gyaneshwar, P.; Paliy, O.; McAuliffe, J.; Popham, D.L.; Jordan, M.I.; Kustu, S. Sulfur and nitrogen limitation in Escherichia coli K-12: Specific homeostatic response. J. Bacteriol. 2005, 187, 1074–1090.
- Merchant, S.S.; Helmann, J.D. Elemental economy: Microbial strategies for optimizing growth in the face of nutrient limitation. Adv. Microb. Physiol. 2012, 60, 91–210.
- Cuhel, R.L.; Taylor, C.D.; Jannasch, H.W. Assimilatory sulfur metabolism in marine microorganisms: Sulfur metabolism, growth, and protein synthesis of Pseudomonas halodurans and Alteromonas luteo-violaceus during sulfate limitation. Arch. Microbiol. 1981, 130, 1–7.
- Mazel, D.; Marlière, P. Adaptive eradication of methionine and cysteine from cyanobacterial light-harvesting proteins. Nature 1989, 341, 245–248.
- Robinson, A.; Tempest, D.W. Phenotypic variability of the envelope proteins of Klebsiella aerogenes. J. Gen. Microbiol. 1973, 78, 361–370.
- Pardee, A.B. Purification and properties of a sulfate-binding protein from Salmonella typhimurium. J. Biol. Chem. 1966, 241, 5886–5892.
- Beil, S.; Kertesz, M.A.; Leisinger, T.; Cook, A.M. The assimilation of sulfur from multiple sources and its correlation with expression of the sulfate-starvation-induced stimulon in Pseudomonas putida S-313. Microbiology 1996, 142, 1989–1995.
- Neijssel, O.M.; Tempest, D.W. The regulation of carbohydrate metabolism in Klebsiella aerogenes NCTC 418 organisms, growing in chemostat culture. Arch. Microbiol. 1975, 106, 251–258.
- Koch, A.L.; Deppe, C.S. In vivo assay of protein synthesizing capacity of Escherichia coli from slowly growing chemostat cultures. J. Mol. Biol. 1971, 55, 549–562.
- Chubukov, V.; Sauer, U. Environmental dependence of stationary-phase metabolism in Bacillus subtilis and Escherichia coli. Appl. Environ. Microbiol. 2014, 80, 2901–2909.
- Stouthamer, A.H.; Bettenhaussen, C.W. Determination of the efficiency of oxidative phosphorylation in continuous cultures of Aerobacter aerogenes. Arch. Microbiol. 1975, 102, 187–192.
- Zinebi, S.; Raval, G.; Petitdemange, H. Effect of oxygenation and sulfate concentrations on pyruvate and lactate formation in Klebsiella oxytoca ZS growing in chemostat culture. Curr. Microbiol. 1994, 29, 79–85.
- Chew, L.C.K.; Tacon, W.C.A.; Cole, J.A. Effect of growth conditions on the rate of loss of the plasmid pAT153 from continuous cultures of Escherichia coli HB101. FEMS Microbiol. 1988, 56, 101–104.
- Masuda, A.; Toya, Y.; Shimizu, H. Metabolic impact of nutrient starvation in mevalonate-producing Escherichia coli. Bioresour. Technol. 2017, 245, 1634–1640.
- Li, S.; Jendresen, C.B.; Nielsen, A.T. Increasing production yield of tyrosine and mevalonate through inhibition of biomass formation. Process Biochem. 2016, 51, 1992–2000.
- Willrodt, C.; Hoschek, A.; Bühler, B.; Schmid, A.; Julsing, M.K. Decoupling production from growth by magnesium sulphate limitation boosts de novo limonene production. Biotechnol. Bioeng. 2016, 113, 1305–1314.
- Wu, S.; Zhao, X.; Shen, H.; Wang, Q.; Zhao, Z.K. Microbial lipid production by Rhodosporidium toruloides under sulfate-limited conditions. Bioresour. Technol. 2011, 102, 1803–1807.
- Tissières, A.; Watson, J.D. Ribonucleoprotein particles from Escherichia coli. Nature 1958, 182, 778–780.
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656.
- Hartwig, A. Role of magnesium in genomic stability. Mutat. Res 2001, 475, 113–121.
- Christensen, D.G.; Orr, J.G.; Rao, V.G.; Wolfe, A.L. Increasing growth yield and decreasing acetylation in Escherichia coli by optimizing the carbon to magnesium ratio in peptide-based medium. Appl. Environ. Microbiol. 2017, 83, e03034-16.
- Warsi, O.M.; Andersson, D.I.; Dykhuizen, D.E. Different adaptive strategies in E. coli populations evolving under macronutrient limitation and metal ion limitation. BMC Evol. Biol. 2018, 18, 72.
- Fiil, A.; Branton, D. Changes in the plasma membrane of Escherichia coli during magnesium starvation. J. Bacteriol. 1969, 98, 1320–1327.
- Tempest, D.W.; Hunter, J.R.; Sykes, J. Magnesium-limited growth of Aerobacter aerogenes in a chemostat. J. Gen. Microbiol. 1965, 39, 355–366.
- Brock, T.D. Effects of magnesium ion deficiency on Escherichia coli and possible relation to the mode of action of novobiocin. J. Bacteriol. 1962, 84, 679–682.
- Dean, A.C.R.; Rogers, P.L. The cell size and macromolecular composition of Aerobacter aerogenes in various systems of continuous culture. Biochim. Biophys. Acta 1962, 148, 267–279.
- Smit, G.; Straver, M.H.; Lugtenberg, B.J.; Kijne, J.W. Flocculence of Saccharomyces cerevisiae cells is induced by nutrient limitation, with cell surface hydrophobicity as a major determinant. Appl. Environ. Microbiol. 1992, 58, 3709–3714.
- Folsom, J.P.; Parker, A.E.; Carlson, R.P. Physiological and proteomic analysis of Escherichia coli iron-limited chemostat growth. J. Bacteriol. 2014, 196, 2748–2761.
- Parente, A.F.A.; Bailão, A.M.; Borges, C.L.; Parente, J.A.; Magalhães, A.D.; Ricart, C.A.O.; Soares, C.M.A. Proteomic Analysis Reveals That Iron Availability Alters the Metabolic Status of the Pathogenic Fungus Paracoccidioides brasiliensis. PLoS ONE 2011, 6, e22810.
- McHugh, J.P.; Rodríguez-Quiñones, F.; Abdul-Tehrani, H.; Svistunenko, D.A.; Poole, R.K.; Cooper, C.E.; Andrews, S.C. Global iron-dependent gene regulation in Escherichia coli: A new mechanism for iron homeostasis. J. Biol. Chem. 2003, 278, 29478–29486.
- Friedman, D.B.; Stauff, D.L.; Pishchany, G.; Whitwell, C.W.; Torres, V.J.; Skaar, E.P. Staphylococcus aureus redirects central metabolism to increase iron availability. PloS Path. 2006, 2, 777–789.
- Löser, C.; Urit, T.; Stukert, A.; Bley, T. Formation of ethyl acetate from whey by Kluyveromyces marxianus on a pilot scale. J. Biotechnol. 2013, 163, 17–23.
- Armstrong, D.W.; Yamazaki, H. Effect of iron and EDTA on ethyl acetate accumulation in Candida utilis. Biotechnol. Lett. 1984, 6, 819–824.
- Baez, A.; Sharma, A.K.; Bryukhanov, A.; Anderson, E.D.; Rudack, L.; Olivares-Hernandez, R.; Quan, D.; Shiloach, J. Iron availability enhances the cellular energetics of aerobic Escherichia coli cultures while upregulating anaerobic respiratory chains. New Biotechnol. 2022, 71, 11–20.
- Georgiadis, M.M.; Komiya, H.; Chakrabarti, P.; Woo, D.; Kornuc, J.J.; Rees, D.C. Crystallographic structure of the nitrogenase iron protein from Azotobacter vinelandii. Science 1992, 257, 1653–1659.
- Fekete, F.A.; Spence, J.T.; Emery, T. Siderophores produced by nitrogen-fixing Azotobacter vinelandii OP in iron-limited continuous culture. Appl. Environ. Microbiol. 1983, 46, 1297–1300.
- MacLeod, R.A.; Snell, E.E. Some mineral requirements of the lactic acid bacteria. J. Biol. Chem. 1947, 170, 351–365.
- Archibald, F. Lactobacillus plantarum, an organism not requiring iron. FEMS Microbiol. Lett. 1983, 19, 29–32.

