 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Huixia Liu | -- | 1828 | 2022-11-01 09:58:01 | | | |
| 2 | Huixia Liu | + 542 word(s) | 2370 | 2022-11-01 11:27:00 | | | | |
| 3 | Catherine Yang | + 2 word(s) | 2372 | 2022-11-02 01:54:08 | | |
Video Upload Options
Partial pressure of oxygen (pO2) in the kidney is maintained at a relatively stable level by a unique and complex functional interplay between renal blood flow, glomerular filtration rate (GFR), oxygen consumption, and arteriovenous oxygen shunting. The vulnerability of this interaction renders the kidney vulnerable to hypoxic injury leading to different renal diseases. Hypoxia has long been recognized as an important factor in the pathogenesis of acute kidney injury (AKI), especially renal ischemia/reperfusion injury. Accumulating evidence suggests that hypoxia also plays an important role in the pathogenesis and progression of chronic kidney disease (CKD) and CKD-related complications, such as anemia, cardiovascular events, and sarcopenia. In addition, renal cancer is linked to deregulation of hypoxia pathways. Renal utilizes various molecular pathways to respond and adapt to changes in renal oxygenation. Particularly, hypoxia-inducible factor (HIF) (including HIF-1, 2, 3) has been shown to be activated in renal disease and plays a major role in the protective response to hypoxia. HIF-1 is a heterodimer that is composed of an oxygen-regulated HIF-1α subunit and a constitutively expressed HIF-1β subunit. In renal diseases, the critical characteristic of HIF-1α is protective, but it also has a negative effect, such as in sarcopenia. This review summarizes the mechanisms of HIF-1α regulation in renal disease.
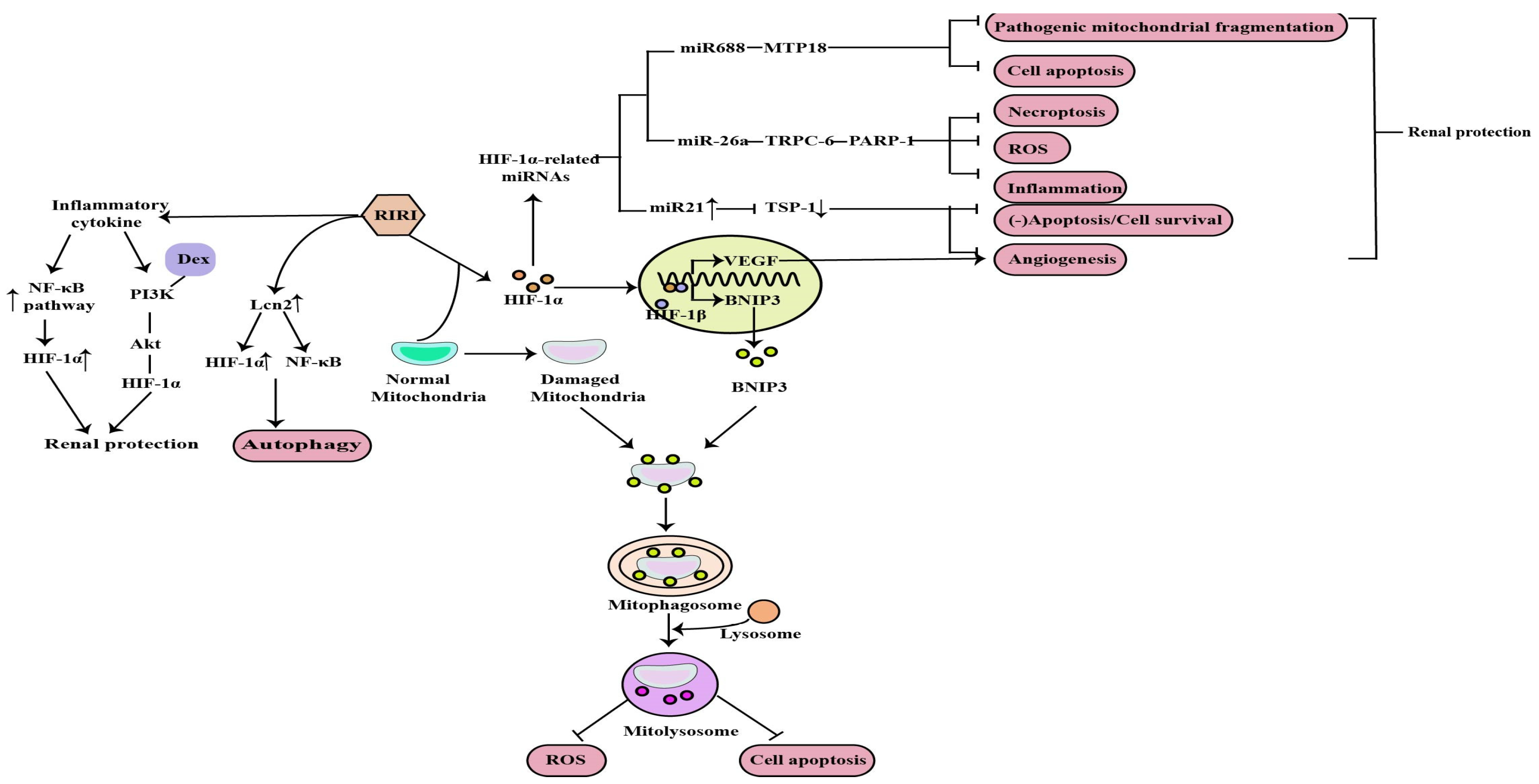
1. The Role of HIF-1α in RIRI
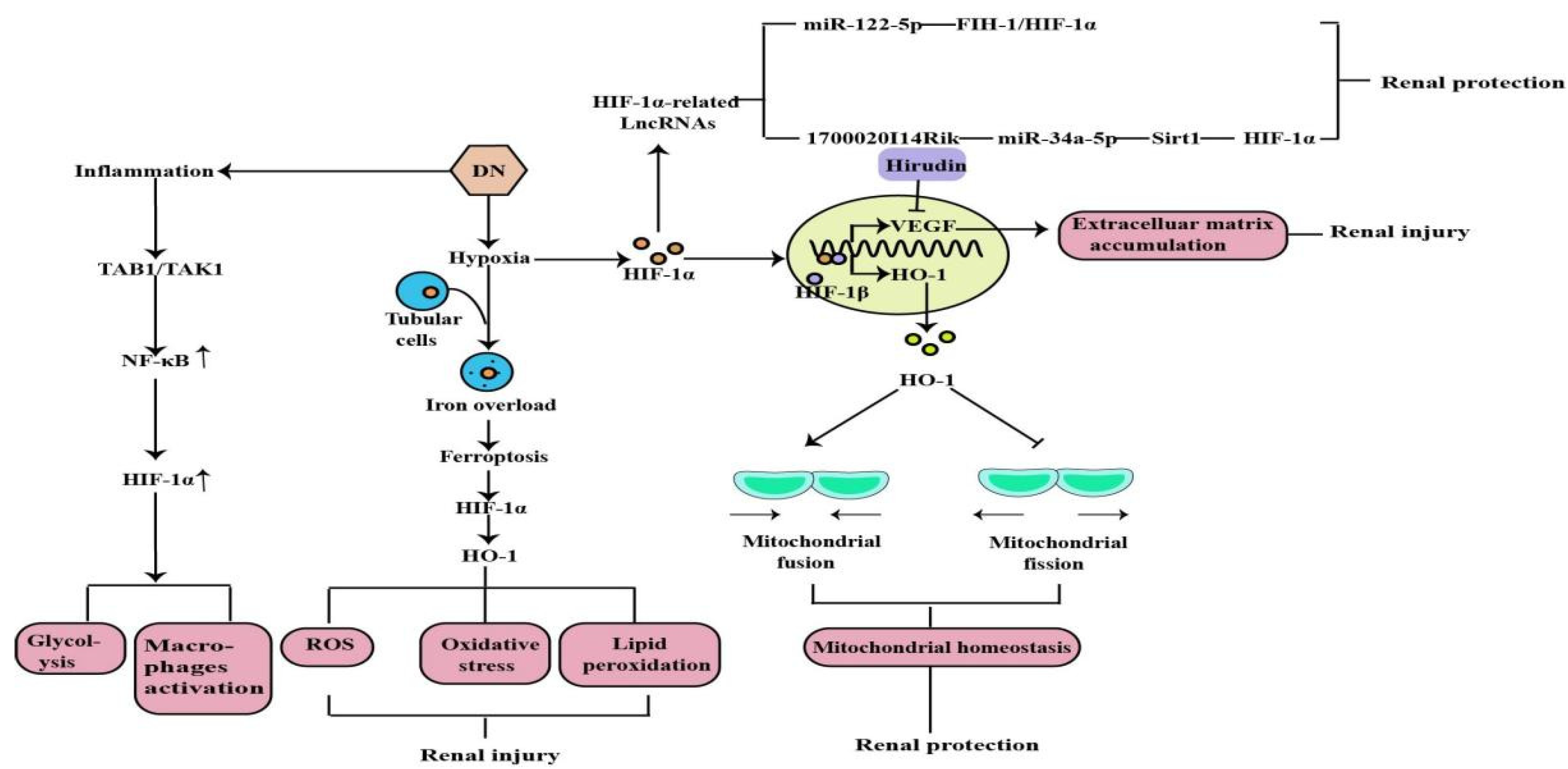
2. The Role of HIF-1α in Diabetic Nephropathy
3. The Role of HIF-1α in Chronic Kidney Disease-Related Complications
3.1. Renal Anemia

3.2. Cardiovascular Disease
3.3. Sarcopenia
4. The Role of HIF-1α in Renal Cancer
Hypoxia is common in many types of solid tumors, in which tumor cells rapidly proliferate and form large solid tumor masses, resulting in the obstruction and compression of the blood vessels around these masses. These abnormal vessels usually do not function properly, causing poor O2 supply to the central tumor region [29]. Renal cancer is a common urological malignancy with limited therapeutic options for metastatic disease. Most clear cell renal cell carcinomas (ccRCC) are associated with loss of von Hippel-Lindau tumor suppressor (pVHL) function and deregulation of the hypoxia pathway. The VHL tumor suppressor is inactivated in a majority of ccRCC tumors. VHL is the substrate recognition component of an E3 ubiquitin ligase complex containing the elongins B and C, Cullin-2, and Rbx1 that targets the hydroxylation, oxygen-sensitive α-subunits of HIFs for ubiquitination and degradation by the 26S proteasome. The VHL loss results in the constitutive activation of HIF targets, including the pro-angiogenic factors VEGF and platelet derived growth factor (PDGF) [30]. Activated HIF-1 plays a crucial role in the tumor cell adaptive response to oxygen changes by transcriptionally activating more than 100 downstream genes that regulate important biological processes required for tumor survival and progression. Further studies on the HIF would help in developing cancer therapies [31][32] (Figure 4).

Figure 4. ROS removing, mitochondrial metabolism, cell signaling activation are regulated by HIF-1α, which can regulate tumor occurrence and sensitivity through the above mechanisms.
References
- Guo, A.; Wang, W.; Shi, H.; Wang, J.; Liu, T. Identification of Hub Genes and Pathways in a Rat Model of Renal Ischemia-Reperfusion Injury Using Bioinformatics Analysis of the Gene Expression Omnibus (GEO) Dataset and Integration of Gene Expression Profiles. Med. Sci. Monit. 2019, 25, 8403–8411.
- Qiu, S.; Chen, X.; Pang, Y.; Zhang, Z. Lipocalin-2 protects against renal ischemia/reperfusion injury in mice through autophagy activation mediated by HIF1α and NF-κb crosstalk. Biomed. Pharmacother. 2018, 108, 244–253.
- Jaberi, S.A.; Cohen, A.; D’Souza, C.; Abdulrazzaq, Y.M.; Ojha, S.; Bastaki, S.; Adeghate, E.A. Lipocalin-2: Structure, function, distribution and role in metabolic disorders. Biomed. Pharmacother. 2021, 142, 112002.
- Shen, B.; Mei, M.; Pu, Y.; Zhang, H.; Liu, H.; Tang, M.; Pan, Q.; He, Y.; Wu, X.; Zhao, H. Necrostatin-1 Attenuates Renal Ischemia and Reperfusion Injury via Meditation of HIF-1α/mir-26a/TRPC6/PARP1 Signaling. Mol. Ther. Nucleic Acids 2019, 17, 701–713.
- Wei, H.; Xu, Z.; Chen, L.; Wei, Q.; Huang, Z.; Liu, G.; Li, W.; Wang, J.; Tang, Q.; Pu, J. Long non-coding RNA PAARH promotes hepatocellular carcinoma progression and angiogenesis via upregulating HOTTIP and activating HIF-1α/VEGF signaling. Cell Death Dis. 2022, 13, 102.
- Liu, L.; Zhu, D.; Ding, W.; Zhang, T.; Ma, X.; Zou, J. MiRNA-21- HIF-1α-VEGF Axis is Associated with Myopic Choroidal Neovascularization in Guinea Pigs. Ophthalmic. Res. 2022, 65, 493–505.
- Xu, X.; Song, N.; Zhang, X.; Jiao, X.; Hu, J.; Liang, M.; Teng, J.; Ding, X. Renal Protection Mediated by Hypoxia Inducible Factor-1α Depends on Proangiogenesis Function of miR-21 by Targeting Thrombospondin 1. Transplantation 2017, 101, 1811–1819.
- Sugahara, M.; Tanaka, S.; Tanaka, T.; Saito, H.; Ishimoto, Y.; Wakashima, T.; Ueda, M.; Fukui, K.; Shimizu, A.; Inagi, R.; et al. Prolyl Hydroxylase Domain Inhibitor Protects against Metabolic Disorders and Associated Kidney Disease in Obese Type 2 Diabetic Mice. J. Am. Soc. Nephrol. 2020, 31, 560–577.
- Yamazaki, T.; Mimura, I.; Tanaka, T.; Nangaku, M. Treatment of Diabetic Kidney Disease: Current and Future. Diabetes Metab. J. 2021, 45, 11–26.
- Rosenberger, C.; Khamaisi, M.; Abassi, Z.; Shilo, V.; Weksler-Zangen, S.; Goldfarb, M.; Shina, A.; Zibertrest, F.; Eckardt, K.U.; Rosen, S.; et al. Adaptation to hypoxia in the diabetic rat kidney. Kidney Int. 2008, 73, 34–42.
- Chiang, S.K.; Chen, S.E.; Chang, L.C. A Dual Role of Heme Oxygenase-1 in Cancer Cells. Int. J. Mol. Sci. 2018, 20, 39.
- Song, L.; Wang, K.; Yin, J.; Yang, Y.; Li, B.; Zhang, D.; Wang, H.; Wang, W.; Zhan, W.; Guo, C.; et al. Traditional Chinese Medicine Fufang-Zhenzhu-Tiaozhi capsule prevents renal injury in diabetic minipigs with coronary heart disease. Chin. Med. 2022, 17, 102.
- Sun, H.K.; Lee, Y.M.; Han, K.H.; Kim, H.S.; Ahn, S.H.; Han, S.Y. Phosphodiesterase inhibitor improves renal tubulointerstitial hypoxia of the diabetic rat kidney. Korean J. Intern. Med. 2012, 27, 163–170.
- Feng, X.; Wang, S.; Sun, Z.; Dong, H.; Yu, H.; Huang, M.; Gao, X. Ferroptosis Enhanced Diabetic Renal Tubular Injury via HIF-1α/HO-1 Pathway in db/db Mice. Front. Endocrinol. 2021, 12, 626390.
- Jiang, N.; Zhao, H.; Han, Y.; Li, L.; Xiong, S.; Zeng, L.; Xiao, Y.; Wei, L.; Xiong, X.; Gao, P.; et al. HIF-1α ameliorates tubular injury in diabetic nephropathy via HO-1-mediated control of mitochondrial dynamics. Cell Prolif. 2020, 53, e12909.
- Cheng, L.; Qiu, X.; He, L.; Liu, L. MicroRNA-122-5p ameliorates tubular injury in diabetic nephropathy via FIH-1/HIF-1α pathway. Renal Fail. 2022, 44, 293–303.
- Chen, Y.; He, Y.; Zhou, H. The potential role of lncRNAs in diabetes and diabetic microvascular complications. Endocr. J. 2020, 67, 659–668.
- Li, A.; Peng, R.; Sun, Y.; Liu, H.; Peng, H.; Zhang, Z. LincRNA 1700020I14Rik alleviates cell proliferation and fibrosis in diabetic nephropathy via miR-34a-5p/Sirt1/HIF-1α signaling. Cell Death Dis. 2018, 9, 461.
- Zeng, H.; Qi, X.; Xu, X.; Wu, Y. TAB1 regulates glycolysis and activation of macrophages in diabetic nephropathy. Inflamm. Res. 2020, 69, 1215–1234.
- Pang, X.; Zhang, Y.; Shi, X.; Peng, Z.; Xing, Y.; Jiarui, H. Hirudin Reduces the Expression of Markers of the Extracellular Matrix in Renal Tubular Epithelial Cells in a Rat Model of Diabetic Kidney Disease Through the Hypoxia-Inducible Factor-1α (HIF-1α)/Vascular Endothelial Growth Factor (VEGF) Signaling Pathway. Med. Sci. Monit. 2020, 26, e921894.
- Rosenberger, C.; Mandriota, S.; Jürgensen, J.S.; Wiesener, M.S.; Hörstrup, J.H.; Frei, U.; Ratcliffe, P.J.; Maxwell, P.H.; Bachmann, S.; Eckardt, K.U. Expression of hypoxia-inducible factor-1alpha and -2alpha in hypoxic and ischemic rat kidneys. J. Am. Soc. Nephrol. 2002, 13, 1721–1732.
- Koury, M.J.; Haase, V.H. Anaemia in kidney disease: Harnessing hypoxia responses for therapy. Nat. Rev. Nephrol. 2015, 11, 394–410.
- Voit, R.A.; Sankaran, V.G. Stabilizing HIF to Ameliorate Anemia. Cell 2020, 180, 6.
- Li, Z.L.; Tu, Y.; Liu, B.C. Treatment of Renal Anemia with Roxadustat: Advantages and Achievement. Kidney Dis. 2020, 6, 65–73.
- Iyer, N.V.; Kotch, L.E.; Agani, F.; Leung, S.W.; Laughner, E.; Wenger, R.H.; Gassmann, M.; Gearhart, J.D.; Lawler, A.M.; Yu, A.Y.; et al. Cellular and developmental control of O2 homeostasis by hypoxia-inducible factor 1 alpha. Genes Dev. 1998, 12, 149–162.
- Knutson, A.K.; Williams, A.L.; Boisvert, W.A.; Shohet, R.V. HIF in the heart: Development, metabolism, ischemia, and atherosclerosis. J. Clin. Investig. 2021, 131, e137557.
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W. Vascular nitric oxide: Beyond eNOS. J. Pharmacol. Sci. 2015, 129, 83–94.
- Cirillo, F.; Mangiavini, L.; La Rocca, P.; Piccoli, M.; Ghiroldi, A.; Rota, P.; Tarantino, A.; Canciani, B.; Coviello, S.; Messina, C.; et al. Human Sarcopenic Myoblasts Can Be Rescued by Pharmacological Reactivation of HIF-1α. Int. J. Mol. Sci. 2022, 23, 7114.
- Li, F.; Aljahdali, I.A.M.; Zhang, R.; Nastiuk, K.L.; Krolewski, J.J.; Ling, X. Kidney cancer biomarkers and targets for therapeutics: Survivin (BIRC5), XIAP, MCL-1, HIF1α, HIF2α, NRF2, MDM2, MDM4, p53, KRAS and AKT in renal cell carcinoma. J. Exp. Clin. Cancer Res. 2021, 40, 254.
- Xiao, Y.; Thakkar, K.N.; Zhao, H.; Broughton, J.; Li, Y.; Seoane, J.A.; Diep, A.N.; Metzner, T.J.; von Eyben, R.; Dill, D.L.; et al. The m(6)A RNA demethylase FTO is a HIF-independent synthetic lethal partner with the VHL tumor suppressor. Proc. Natl. Acad. Sci. USA 2020, 117, 21441–21449.
- Ullah, A.; Leong, S.W.; Wang, J.; Wu, Q.; Ghauri, M.A.; Sarwar, A.; Su, Q.; Zhang, Y. Cephalomannine inhibits hypoxia-induced cellular function via the suppression of APEX1/HIF-1α interaction in lung cancer. Cell Death Dis. 2021, 12, 490.
- Ullah, A.; Ullah, N.; Nawaz, T.; Aziz, T. Molecular mechanisms of Sanguinarine in cancer prevention and treatment. Anti-Cancer Agents Med. Chem. 2022.

