Partial pressure of oxygen (pO2) in the kidney is maintained at a relatively stable level by a unique and complex functional interplay between renal blood flow, glomerular filtration rate (GFR), oxygen consumption, and arteriovenous oxygen shunting. The vulnerability of this interaction renders the kidney vulnerable to hypoxic injury leading to different renal diseases. Hypoxia has long been recognized as an important factor in the pathogenesis of acute kidney injury (AKI), especially renal ischemia/reperfusion injury. Accumulating evidence suggests that hypoxia also plays an important role in the pathogenesis and progression of chronic kidney disease (CKD) and CKD-related complications, such as anemia, cardiovascular events, and sarcopenia. In addition, renal cancer is linked to deregulation of hypoxia pathways. Renal utilizes various molecular pathways to respond and adapt to changes in renal oxygenation. Particularly, hypoxia-inducible factor (HIF) (including HIF-1, 2, 3) has been shown to be activated in renal disease and plays a major role in the protective response to hypoxia. HIF-1 is a heterodimer that is composed of an oxygen-regulated HIF-1α subunit and a constitutively expressed HIF-1β subunit. In renal diseases, the critical characteristic of HIF-1α is protective, but it also has a negative effect, such as in sarcopenia. This review summarizes the mechanisms of HIF-1α regulation in renal disease.
- hypoxia-inducible factor-1α
- renal cancer
- renal ischemia/reperfusion injury
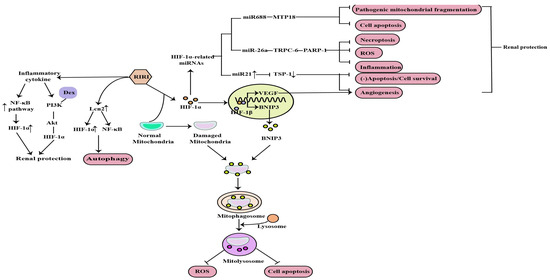
1. The Role of HIF-1α in RIRI
2. The Role of HIF-1α in Diabetic Nephropathy
DN is one of the most common models of CKD and the leading cause of ESRD. Earlier studies have demonstrated that hypoxia is an early event in the development and progression of experimental DN, and an increased HIF-1α expression in diabetic kidneys compared to the kidneys of control rats and normal human kidneys [8][9][10][37,38,39] (Figure 23). Ferroptosis is a recently discovered form of iron-dependent cell death. Heme is a main source of synthetic iron, and heme oxygenase (HO)-1 metabolizes heme into biliverdin/bilirubin, carbon monoxide, and ferrous iron. HO-1 can be induced by a variety of cues, including inflammatory mediators, oxidants, and physical or chemical stimuli, and HO-1 is one of the HIF target genes [11][40]. Chronic hypoxia due to renal ischemia induces the increase of HIF-1α in renal tubules of diabetic models, with the elevated HO-1 level [12][13][41,42]. Feng et al. indicated that ferroptosis might enhance DN and damage renal tubules in DN models through the HIF-1α/HO-1 pathway [14][43]. However, a study in 2020 demonstrated that HIF-1α improved mitochondrial dysfunction and restricted mitochondria-dependent apoptosis in the tubular cells of DN via the HO-1 pathway. In addition, the HIF-1α/HO-1 pathway is the pivotal pathway mediating tubular cell mitochondrial dynamics in DN [15][18].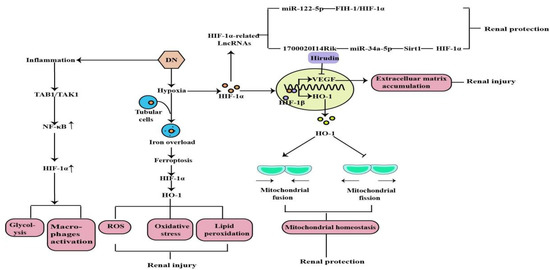
3. The Role of HIF-1α in Chronic Kidney Disease-Related Complications
3.1. Renal Anemia
Anemia is a common complication of CKD, mainly due to injured kidneys failing to produce sufficient amounts of EPO, which regulates red blood cell production. Hypoxia serves as the major stimulus of EPO production. Iron is essential in erythropoiesis. Iron deficiency also occurs in CKD patients due to the inadequate provision or absorption of dietary iron (Figure 34). HIF-1α is mainly induced in renal tubular and glomerular epithelial cells and in papillary interstitial cells, whereas HIF-2α is expressed in endothelial cells and fibroblasts upon hypoxic stimulation, suggesting that HIF-2α is the main regulator of EPO production [21][49]. Under hypoxic conditions, HIF-2α regulates EPO expression in combination with hypoxia response elements on the EPO gene in the kidney and liver [22][50]. Voit et al. revealed that EPO production in the kidney can also be regulated by HIF-1α, which is degraded under normoxic conditions by HIF-prolyl hydroxylase (HIF-PHD) [23][19]. The discovery of prolyl hydroxylase domain (PHD) enzymes as regulators of hypoxia-inducible factor (HIF)-dependent erythropoiesis has led to the development of novel therapeutic agents for renal anemia [24][51]. Roxadustat, the first small-molecule PHI, can lead to increased EPO production, better iron absorption, and amelioration of anemia in CKD [23][19]. In addition, HIF-2α also regulates iron metabolism by stimulating duodenal cytochrome B (DCYTB) and divalent metal transporter-1 (DMT1) expression.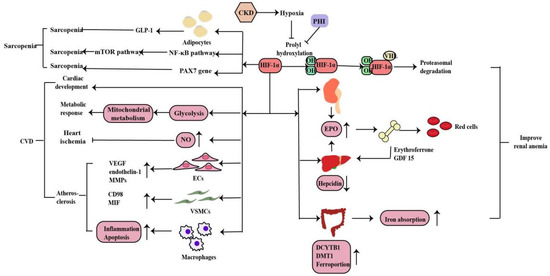
3.2. Cardiovascular Disease
CVD is the main cause of death in patients with CKD. Hypoxia is also a promoting factor of myocardial infarction, cardiac remodeling, atherosclerosis and peripheral arterial disease in CKD patients. Under hypoxia, HIF signaling affects the development, metabolic response, ischemia and atherosclerosis in heart disease in myriad ways (Figure 4). Animal experiments have shown that the HIF-1α deletion has a wide range of cardiac abnormalities, including septation and trabeculation, and is affected by specific alleles and the mouse genetic background [25][52]. Thus, HIF-regulated transcription is necessary for important cardiac developmental events. Cellular metabolism is inextricably linked with cardiac contractility and HIF plays a crucial role in shaping the metabolic response in the heart. HIF-1α promotes glycolysis during hypoxia and influences mitochondrial metabolism to proper cardiometabolic function [26][20]. As a vasodilator, NO plays a vital role in the regulation of vascular tone through the regulating cGMP in smooth muscle cells, S-nitrosylation of target proteins, the activation of sarco/endoplasmic reticulum calcium ATPase and the production of cyclic inosine monophosphate. HIF-1α can activate inducible nitric oxide synthase (iNOS) gene expression by increasing NO synthesis to limit ischemic damage in the heart [27][53]. The effects of HIF-1α in the three most important cell types of atherosclerosis--macrophages, vascular smooth muscle cells (VSMCs), and endothelial cells (ECs) are essential. Whereas HIF-1α directly induces VEGF, endothelin-1, and matrix metalloproteinases (MMPs) in endothelial cells to facilitate angiogenesis, its effect on vascular smooth muscle cells is to induce proliferation of these cells in the atheroma by up-regulating factors such as CD98 and macrophage migration inhibitory factor (MIF). HIF-1α also modulates the function of diseased macrophage foam cells by making the cells more inflammatory and apoptotic while simultaneously inhibiting their ability to metabolize lipids [26][20].3.3. Sarcopenia
Sarcopenia is one of the complications of CKD patients. Hypoxia induces oxygen delivery reduction to the muscles, which is more obvious in anemia with CKD. Mechanistically, HIF-1α deletion stimulates GLP-1 secretion in human adipocytes, which contributes to sarcopenia in hypoxia[54]. What’s more, hypoxia induces muscle wasting by activating the HIF-1α and NF-kB catabolic pathways and inhibiting the anabolic mammalian target of rapamycin (mTOR) pathway[55,56]. Cirillo et al. revealed that a decrease in the expression of HIF-1α and its target genes in sarcopenia, as well as of PAX7, the major stem cell marker of satellite cells, whereas the atrophy marker MURF1 was increased. And a pharmacological activation of HIF-1α and its target genes caused a reduction in skeletal muscle atrophy and activation of PAX7 gene expression. Thus, HIF-1α plays a role in sarcopenia and is involved in satellite cell homeostasis but need further research[57](Figure [28](Figure 44).4. The Role of HIF-1α in Renal CancerThe role of HIF-1α in renal cancer
Hypoxia is common in many types of solid tumors, in which tumor cells rapidly proliferate and form large solid tumor masses, resulting in the obstruction and compression of the blood vessels around these masses. These abnormal vessels usually do not function properly, causing poor O2 supply to the central tumor region [29][58]. Renal cancer is a common urological malignancy with limited therapeutic options for metastatic disease. Most clear cell renal cell carcinomas (ccRCC) are associated with loss of von Hippel-Lindau tumor suppressor (pVHL) function and deregulation of the hypoxia pathway. The VHL tumor suppressor is inactivated in a majority of ccRCC tumors. VHL is the substrate recognition component of an E3 ubiquitin ligase complex containing the elongins B and C, Cullin-2, and Rbx1 that targets the hydroxylation, oxygen-sensitive α-subunits of HIFs for ubiquitination and degradation by the 26S proteasome. The VHL loss results in the constitutive activation of HIF targets, including the pro-angiogenic factors VEGF and platelet derived growth factor (PDGF) [30][59]. Activated HIF-1 plays a crucial role in the tumor cell adaptive response to oxygen changes by transcriptionally activating more than 100 downstream genes that regulate important biological processes required for tumor survival and progression. Further studies on the HIF would help in developing cancer therapies[60,61](Figure [31][32] (Figure 45).
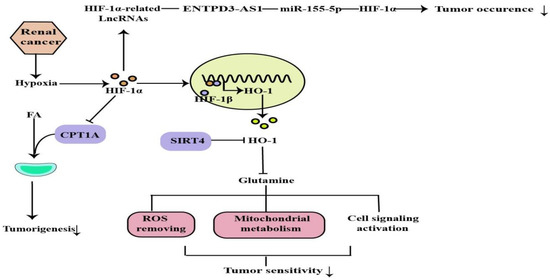
Figure 4. ROS removing, mitochondrial metabolism, cell signaling activation are regulated by HIF-1α, which can regulate tumor occurrence and sensitivity through the above mechanisms.