 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Dong Zhang | -- | 2204 | 2022-03-30 12:28:00 | | | |
| 2 | Dong Zhang | + 3960 word(s) | 6164 | 2022-03-30 12:41:08 | | | | |
| 3 | Dong Zhang | Meta information modification | 6164 | 2022-03-30 12:42:14 | | | | |
| 4 | Dong Zhang | Meta information modification | 6164 | 2022-03-30 12:43:07 | | | | |
| 5 | Amina Yu | -72 word(s) | 6092 | 2022-03-31 03:49:34 | | | | |
| 6 | Amina Yu | -4 word(s) | 6088 | 2022-03-31 04:10:39 | | | | |
| 7 | Amina Yu | -67 word(s) | 6021 | 2022-03-31 05:38:50 | | | | |
| 8 | Amina Yu | -35 word(s) | 5986 | 2022-03-31 06:24:40 | | |
Video Upload Options
Adventitious root (AR) formation is required for the vegetative propagation of economically important horticultural crops, such as apples. Asexual propagation is commonly utilized for breeding programs because of its short life cycle, true-to-typeness, and high efficiency. The lack of AR formation from stem segments is a barrier to segment survival.
1. Multiple Hormonal Pathways Mediate Adventitious Rooting
After separation from the donor plants, stem cuttings critically modify hormone homeostasis in the detached shoots [1]. Different growth regulators were examined to increase the rooting ability of stem cuttings. In the last few decades, it has been conducted on the formation of ARs in apples. Different endogenous hormones play a key role in forming ARs in apples. Auxin, as a master controller, promotes AR induction and initiation stages while inhibiting the AR emergence stage. Auxin crosstalk with melatonin (MT) also promotes the AR induction stage. Cytokinin (CK) works antagonistically to auxin. Gibberellic acid 3 (GA3) also plays a negative role in the initial stages. Moreover, ethylene (ET) and jasmonic acid (JA) can inhibit the induction stage, and their roles in other stages are unclear. Abscisic acid (ABA) is a negative regulator of AR formation at all stages.
1.1. Auxin: A Master Regulator for ARs
Auxin is involved in various physiological events, including vascular differentiation, cell expansion, lateral roots (LRs), and ARs. More auxin is needed to induce ARs, but this is not necessary for the AR emergence stage [2]. Auxin treatment was shown to boost assimilate translocation from the leaf and sugar content at the root growth site [3]. Indole-3-acetic acid (IAA) is perhaps the most prevalent natural auxin, but Indole-3-butyric acid (IBA) is the most frequently utilized exogenous auxin for increasing ARs in most species, particularly in difficult-to-root genotypes. Recently, it was conducted on M 116 apple clonal rootstock to identify the effects of different concentrations of IBA and naphthaleneacetic acid (NAA) (IBA: T1, 1500; T2, 2000; T3, 2500; T4, 3000; T5, 3500; T6, 4000 ppm; NAA: T7, 500; T8, 1000; T9, 1500 ppm) compared with T10, control plants during adventitious rooting under mist chamber conditions. It was suggested that increasing auxin levels correlated positively with rooting success. IBA treatments significantly improved rooting. T5 had the highest rooting percentage (57.12%), AR numbers per cutting (7.33), AR length (34.85 cm), and AR diameter (5.27 mm) when compared to other treatments and control [4]. Furthermore, the Jork9 was also treated with auxins (IAA, IBA, and NAA). The cultures were kept in darkness during the initial time of rooting treatment. During this phase, the AR initials were made, then cultures were shifted to light. NAA and IAA or IBA treatments obtained the lowest ARs (8) and the highest ARs (15), respectively. The maximal AR numbers were recorded throughout a broad range of IAA levels (10–100 µM), although at only one level of IBA or NAA (10 µM and 3 µM, respectively) [1]. It was indicated that the duration of exposure to auxins mainly controlled the AR formation. The cuttings are not highly responsive to auxin within the first 24 h after being collected. It is believed that during this lag time, cells dedifferentiate and become capable of responding to the rhizogenic signal, auxin. The cells that give rise to the AR primordia are often seen between the vascular bundles and store starch over the first 24 h. The rhizogenic activity of auxin in the induction stage then commits previously activated cells to create AR primordia for up to 72 h. During this time, auxin pulses stimulate the highest AR numbers. Auxin is no longer necessary after 96 h, and auxin levels advantageous for establishing root meristemoids are restrictive during this period [5].
IBA promotes AR formation in M.9 and M.26 apple rootstocks, but higher AR formation was seen in M.26, which is due to the high amount of free IAA in the stem basal part compared to M.9. Furthermore, the conjugated IAA level was higher in M.9 as compared to M.26, suggesting that the difference between AR responses in both rootstocks may be associated with free IAA content in stem basal parts [6]. During the whole process of adventitious rooting, endogenous auxin, or applied externally, plays a critical function at each step. In the AR initial phases, a high level of endogenous auxin is generally associated with a high rooting rate [7]. When the auxin is applied exogenously to increase root formation, it influences the endogenous auxin concentration, which generally reaches a maximum after wounding [8]. Still, the auxin peak was not detected [9]. The endogenous level of auxin induces the AR primordia formation, and the quantity of primordia formation is raised together with the elevation of the IAA level [10]. In apples, IBA was found to be highly important for inducing adventitious rooting in M.26 apple rootstock. The other hormones seem to be indirectly involved in the control of ARs by their interaction with the auxin, where the endogenous level of IAA was observed to be higher at the early stage and induces primordia. Then, the level of IAA decreases towards a later stage [11]. These variations in the IAA levels suggest that auxin plays a critical function at the early stages and may not be crucial after the AR primordia have been established. IBA-treated cuttings of Malus prunifolia var. ringo produced more ARs than cuttings treated with the auxin transport inhibitor NPA, which completely inhibited AR formation. This shows that IBA plays a role in AR formation [12]. In M9-T337 apple stem cutting, IBA inhibited AR elongation at later stages of development by decreasing cell length and by decreasing the expression of genes related to cell elongation [13]. Moreover, transcriptome sequencing situations on IBA-treated T337 cuttings showed that auxin, both endogenous and exogenous, regulates AR formation through homologous signaling pathways to some degree. AR formation is largely controlled via the auxin signaling pathway. Furthermore, various hormone, wounding, as well as sugar signaling pathways work together with the auxin signaling pathway to regulate adventitious rooting in T337 [14]. In M.9 apple rootstock, IAA stimulates the process of AR formation by the upregulation of PIN-FORMED (PIN). Auxin promotes the multiplication and extension of AR founder cells via starch grain hydrolysis, resulting in endomembrane system multiplication, lenticel dehiscence, and AR emergence. This effect was inhibited by the NPA application [15]. Nevertheless, in stem cuttings of Malus species, the IBA promotes more root formation than the IAA. At the same time, it was changed only at deficient IAA levels, implying that the IBA was operational or controlled IAA activity [16].
1.2. Cytokinin: A Required Inhibitor
1.3. Ethylene: A Positive or Negative Regulator for ARs
ET plays a crucial function in regulating ARs in many species. Many experiments were conducted to identify their roles in root formation. The findings of these experiments were extremely flexible for specific species. ET behaved as activators or inhibitors and did not influence the formation of ARs. Because of earlier findings, auxin affects ET synthesis [57]. Subsequently, many efforts have been made to identify how auxin interacts with ET during AR formation. High ET level tissues amplified the responsiveness of root developing tissues in response to endogenous IAA. The role of ET was known in M9-T337 apple stem cutting during AR formation. The AgNO3 (ethylene inhibitor) reduced the appearance of ET and promoted the AR’s emergence and development. However, the ET precursor, 1 aminocyclopropane-1-carboxylic acid (ACC), was added to the MS medium, where it may convert into ET, inhibiting AR emergence and decreasing AR length in M9-T337 [13]. Harbage and Stimart [58] found that ET was not involved in AR formation in apple micro-cuttings of Gala and Triple Red Delicious. It was found that IBA-induced ET formation was reduced by aminoethoxyvinylglycine (AVG), although the AR number continued to be IBA-dependent. ACC restored the inhibitory effect of IBA+AVG on rooting, while ACC separately had little impact on the AR number. Unlike 2,3,5-triiodobenzoic acid (TIBA) and N-1-naphthylphthalamic acid (NPA), which impede polar auxin transport, ET production is increased without increasing the AR number [58]. ET inhibits or promotes the process of root development depending on the stage of the process. It has a stimulatory role at the initial stage but reflects an inhibitory role at later stages of ARs’ development [5]. Root development can also be inhibited by ET, mainly by limiting cell expansion, but this does not affect root meristem activation [59].
1.4. Abscisic Acid: A Negative Regulator for ARs
1.5. Jasmonic Acid: A Positive or Negative Regulator for ARs
1.6. Melatonin: A Positive Regulator
MT is thought to be necessary for the creation and growth of ARs in apples. [37]. In plants, MT served as a vital regulatory signal [77] and was essential for root formation, stress response, explants, and shoots [78][79][80]. It has indicated that the exogenous treatment of MT stimulated AR formation in cuttings of Malus prunifolia, where MT mainly affects the AR induction stage by IAA homeostasis. WOX11 was induced by MT, and apple plants overexpressing MdWOX11 developed more ARs than the GL-3 WT plant, suggesting that MdWOX11 promotes ARs by MT signaling [37]. It has been advocated that exogenous MT at low concentrations could increase the endogenous content of IAA, and it is supposed that this stimulating MT effect on growth and development might be triggered by this rise in IAA levels [81]. On the other hand, it has suggested that the effect of MT on root formation and differentiation is IAA independent [82]. The IAA content was increased at the AR induction stage after the application of MT; however, it was reduced at the AR initiation stage and emergence stage in apple [37].
1.7. Gibberellic acid and brassinosteroids: a positive or negative regulator for ARs
The specific roles of GA and BR are still largely unknown in the regulation of ARs. The effect of GA3 was known in M9 cv. Jork stem discs. First, the discs were cultured in darkness for 24 h on a root-inducing medium containing 24.6 μM IBA. Afterward, the discs were shifted to light exposure and cultured on a hormone-free medium and a medium containing 10 μM GA3 for different time points of adventitious rooting. The results suggest that GA3 treatments limit AR formation from the initial to final stages of AR formation [83]. Moreover, the concentration of GA3 was significantly increased at the initiation of ARs, indicating that GA3 plays an important role in forming apple ARs [12]. However, some shreds of evidence show that BR participated in AR formation. The availability of BRs triggers dual effects on the formation of ARs: the enhancing effect at low levels and the inhibitory effect at high levels [84]. Their high concentration inhibited AR formation in apple rootstock [12]. However, the above information is not enough to identify the specific role of GA and BR in forming ARs.
2. Role of Phenolic Compound in the Regulation of ARs
3. Role of Sugars in the Regulation of ARs
4. Role of Polyamines in the Regulation of ARs
5. Role of Nutrients in the Life of ARs
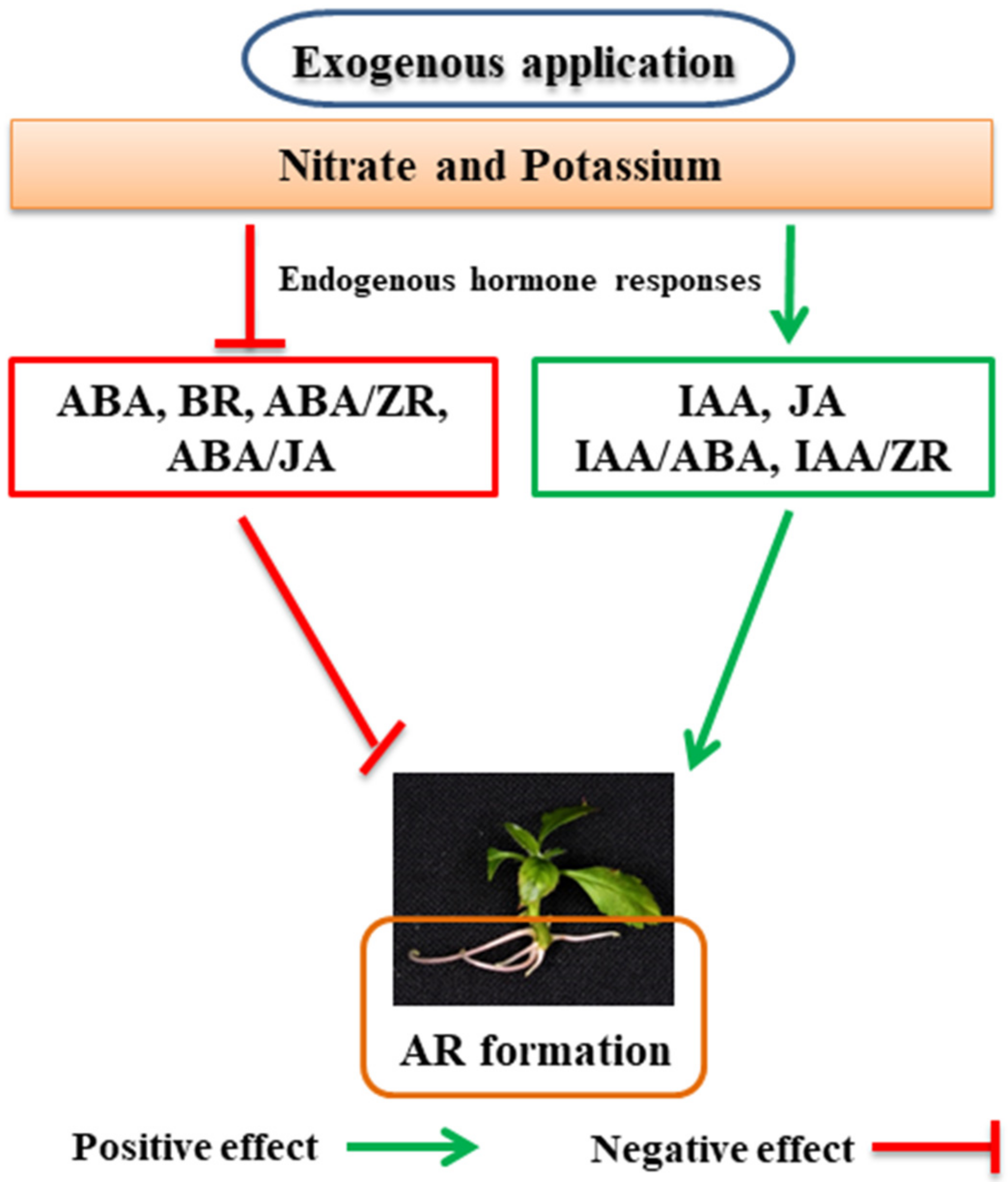
5.1. Role of Nitrogen in the Formation of ARs
5.2. Role of Potassium in the Formation of ARs
References
- De Klerk, G.-J.; Brugge, J.T.; Marinova, S. Effectiveness of indoleacetic acid, indolebutyric acid and naphthaleneacetic acid during adventitious root formation in vitro in Malus ‘Jork 9’. Plant Cell Tissue Organ. Cult. 1997, 49, 39–44.
- Druege, U.; Franken, P.; Hajirezaei, M.R. Plant hormone homeostasis, signaling, and function during adventitious root formation in cuttings. Front. Plant Sci. 2016, 7, 381.
- Agulló-Antón, M.Á.; Sánchez-Bravo, J.; Acosta, M.; Druege, U. Auxins or sugars: What makes the difference in the adventitious rooting of stored carnation cuttings? J. Plant Growth Regul. 2011, 30, 100–113.
- Patial, S.; Chandel, J.; Sharma, N.; Verma, P. Influence of Auxin on Rooting in Hardwood Cuttings of Apple (×Borkh.) Clonal Rootstock′M 116′under Malus domestica Mist Chamber Conditions. Indian J. Ecol. 2021, 48, 429–433.
- de Klerk, G.-J.; van der Krieken, W.; de Jong, J.C. Review the formation of adventitious roots: New concepts, new possibilities. Vitr. Cell. Dev. Biol.-Plant 1999, 35, 189–199.
- Alvarez, R.; Nissen, S.J.; Sutter, E.G. Relationship between indole-3-acetic acid levels in apple (Malus pumila Mill) rootstocks cultured in vitro and adventitious root formation in the presence of indole-3-butyric acid. Plant Physiol. 1989, 89, 439–443.
- Caboni, E.; Tonelli, M.; Lauri, P.; Iacovacci, P.; Kevers, C.; Damiano, C.; Gaspar, T. Biochemical aspects of almond microcuttings related to in vitro rooting ability. Biol. Plant 1997, 39, 91–97.
- Gatineau, F.; Fouché, J.G.; Kevers, C.; Hausman, J.-F.; Gaspar, T. Quantitative variations of indolyl compounds including IAA, IAA-aspartate and serotonin in walnut microcuttings during root induction. Biol. Plant 1997, 39, 131–137.
- Label, P.; Sotta, B.; Miginiac, E. Endogenous levels of abscisic acid and indole-3-acetic acid during in vitro rooting of wild cherry explants produced by micropropagation. Plant Growth Regul. 1989, 8, 325–333.
- Li, X.; He, Y.; Tang, Z. Effects of IAA and stimulated microgravity on formation of adventitious roots of Chinese cabbage. Shi Yan Sheng Wu Xue Bao 2000, 33, 179.
- Meng, Y.; Xing, L.; Li, K.; Wei, Y.; Wang, H.; Mao, J.; Dong, F.; Ma, D.; Zhang, Z.; Han, M. Genome-wide identification, characterization and expression analysis of novel long non-coding RNAs that mediate IBA-induced adventitious root formation in apple rootstocks. Plant Growth Regul. 2019, 87, 287–302.
- Wang, H.; Tahir, M.M.; Nawaz, M.A.; Mao, J.; Li, K.; Wei, Y.; Ma, D.; Lu, X.; Zhao, C.; Zhang, D. Spermidine application affects the adventitious root formation and root morphology of apple rootstock by altering the hormonal profile and regulating the gene expression pattern. Sci. Hortic. 2020, 266, 109310.
- Bai, T.; Dong, Z.; Zheng, X.; Song, S.; Jiao, J.; Wang, M.; Song, C. Auxin and its interaction with ethylene control adventitious root formation and development in apple rootstock. Front. Plant Sci. 2020, 11, 574881.
- Li, K.; Liang, Y.; Xing, L.; Mao, J.; Liu, Z.; Dong, F.; Meng, Y.; Han, M.; Zhao, C.; Bao, L. Transcriptome analysis reveals multiple hormones, wounding and sugar signaling pathways mediate adventitious root formation in apple rootstock. Int. J. Mol. Sci. 2018, 19, 2201.
- Guan, L.; Li, Y.; Huang, K.; Cheng, Z.-M.M. Auxin regulation and MdPIN expression during adventitious root initiation in apple cuttings. Hortic. Res. 2020, 7, 143.
- Van der Krieken, W.; Breteler, H.; Visser, M. Uptake and metabolism of indolebutyric acid during root formation on Malus microcuttings. Acta Bot. Neerl. 1992, 41, 435–442.
- Meng, Y.; Mao, J.; Tahir, M.M.; Wang, H.; Wei, Y.; Zhao, C.; Li, K.; Ma, D.; Zhao, C.; Zhang, D. Mdm-miR160 participates in auxin-induced adventitious root formation of apple rootstock. Sci. Hortic. 2020, 270, 109442.
- Lei, C.; Fan, S.; Li, K.; Meng, Y.; Mao, J.; Han, M.; Zhao, C.; Bao, L.; Zhang, D. iTRAQ-based proteomic analysis reveals potential regulation networks of IBA-induced adventitious root formation in apple. Int. J. Mol. Sci. 2018, 19, 667.
- Gray, W.M.; Kepinski, S.; Rouse, D.; Leyser, O.; Estelle, M. Auxin regulates SCF TIR1-dependent degradation of AUX/IAA proteins. Nature 2001, 414, 271–276.
- Okushima, Y.; Overvoorde, P.J.; Arima, K.; Alonso, J.M.; Chan, A.; Chang, C.; Ecker, J.R.; Hughes, B.; Lui, A.; Nguyen, D. Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: Unique and overlapping functions of ARF7 and ARF19. Plant Cell 2005, 17, 444–463.
- Wilmoth, J.C.; Wang, S.; Tiwari, S.B.; Joshi, A.D.; Hagen, G.; Guilfoyle, T.J.; Alonso, J.M.; Ecker, J.R.; Reed, J.W. NPH4/ARF7 and ARF19 promote leaf expansion and auxin-induced lateral root formation. Plant J. 2005, 43, 118–130.
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 2007, 19, 118–130.
- Mao, J.; Niu, C.; Li, K.; Mobeen Tahir, M.; Khan, A.; Wang, H.; Li, S.; Liang, Y.; Li, G.; Yang, Z. Exogenous 6-benzyladenine application affects root morphology by altering hormone status and gene expression of developing lateral roots in Malus hupehensis. Plant Biol. 2020, 22, 1150–1159.
- Wang, J.-W.; Wang, L.-J.; Mao, Y.-B.; Cai, W.-J.; Xue, H.-W.; Chen, X.-Y. Control of root cap formation by microRNA-targeted auxin response factors in Arabidopsis. Plant Cell 2005, 17, 2204–2216.
- Liu, Z.; Li, J.; Wang, L.; Li, Q.; Lu, Q.; Yu, Y.; Li, S.; Bai, M.Y.; Hu, Y.; Xiang, F. Repression of callus initiation by the mi RNA-directed interaction of auxin–cytokinin in Arabidopsis thaliana. Plant J. 2016, 87, 391–402.
- Li, K.; Liu, Z.; Xing, L.; Wei, Y.; Mao, J.; Meng, Y.; Bao, L.; Han, M.; Zhao, C.; Zhang, D. miRNAs associated with auxin signaling, stress response, and cellular activities mediate adventitious root formation in apple rootstocks. Plant Physiol. Biochem. 2019, 139, 66–81.
- Sorin, C.; Bussell, J.D.; Camus, I.; Ljung, K.; Kowalczyk, M.; Geiss, G.; McKhann, H.; Garcion, C.; Vaucheret, H.; Sandberg, G. Auxin and light control of adventitious rooting in Arabidopsis require ARGONAUTE1. Plant Cell 2005, 17, 1343–1359.
- Tian, C.E.; Muto, H.; Higuchi, K.; Matamura, T.; Tatematsu, K.; Koshiba, T.; Yamamoto, K.T. Disruption and overexpression of auxin response factor 8 gene of Arabidopsis affect hypocotyl elongation and root growth habit, indicating its possible involvement in auxin homeostasis in light condition. Plant J. 2004, 40, 333–343.
- Meng, Y.; Huang, F.; Shi, Q.; Cao, J.; Chen, D.; Zhang, J.; Ni, J.; Wu, P.; Chen, M. Genome-wide survey of rice microRNAs and microRNA–target pairs in the root of a novel auxin-resistant mutant. Planta 2009, 230, 883–898.
- Marin, E.; Jouannet, V.; Herz, A.; Lokerse, A.S.; Weijers, D.; Vaucheret, H.; Nussaume, L.; Crespi, M.D.; Maizel, A. miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. Plant Cell 2010, 22, 1104–1117.
- Xu, X.; Li, X.; Hu, X.; Wu, T.; Wang, Y.; Xu, X.; Zhang, X.; Han, Z. High miR156 expression is required for auxin-induced adventitious root formation via MxSPL26 independent of PINs and ARFs in Malus xiaojinensis. Front. Plant Sci. 2017, 8, 1059.
- Levy, A.; Szwerdszarf, D.; Abu-Abied, M.; Mordehaev, I.; Yaniv, Y.; Riov, J.; Arazi, T.; Sadot, E. Profiling microRNAs in Eucalyptus grandis reveals no mutual relationship between alterations in miR156 and miR172 expression and adventitious root induction during development. BMC Genom. 2014, 15, 524.
- Grieneisen, V.A.; Xu, J.; Marée, A.F.; Hogeweg, P.; Scheres, B. Auxin transport is sufficient to generate a maximum and gradient guiding root growth. Nature 2007, 449, 1008–1013.
- Ioio, R.D.; Nakamura, K.; Moubayidin, L.; Perilli, S.; Taniguchi, M.; Morita, M.T.; Aoyama, T.; Costantino, P.; Sabatini, S. A genetic framework for the control of cell division and differentiation in the root meristem. Science 2008, 322, 1380–1384.
- Bennett, M.J.; Marchant, A.; Green, H.G.; May, S.T.; Ward, S.P.; Millner, P.A.; Walker, A.R.; Schulz, B.; Feldmann, K.A. Arabidopsis AUX1 gene: A permease-like regulator of root gravitropism. Science 1996, 273, 948–950.
- Marchant, A.; Bhalerao, R.; Casimiro, I.; Eklöf, J.; Casero, P.J.; Bennett, M.; Sandberg, G. AUX1 promotes lateral root formation by facilitating indole-3-acetic acid distribution between sink and source tissues in the Arabidopsis seedling. Plant Cell 2002, 14, 589–597.
- Mao, J.; Niu, C.; Li, K.; Chen, S.; Tahir, M.M.; Han, M.; Zhang, D. Melatonin promotes adventitious root formation in apple by promoting the function of MdWOX11. BMC Plant Biol. 2020, 20, 536.
- Liu, B.; Wang, L.; Zhang, J.; Li, J.; Zheng, H.; Chen, J.; Lu, M. WUSCHEL-related Homeobox genes in Populus tomentosa: Diversified expression patterns and a functional similarity in adventitious root formation. BMC Genom. 2014, 15, 296.
- Tian, H.; Jia, Y.; Niu, T.; Yu, Q.; Ding, Z. The key players of the primary root growth and development also function in lateral roots in Arabidopsis. Plant Cell Rep. 2014, 33, 745–753.
- Hu, X.; Xu, L. Transcription factors WOX11/12 directly activate WOX5/7 to promote root primordia initiation and organogenesis. Plant Physiol. 2016, 172, 2363–2373.
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517.
- Sozzani, R.; Cui, H.; Moreno-Risueno, M.; Busch, W.; Van Norman, J.; Vernoux, T.; Brady, S.; Dewitte, W.; Murray, J.A.H.; Benfey, P. Spatiotemporal regulation of cell-cycle genes by SHORTROOT links patterning and growth. Nature 2010, 466, 128–132.
- Liu, J.; Sheng, L.; Xu, Y.; Li, J.; Yang, Z.; Huang, H.; Xu, L. WOX11 and 12 are involved in the first-step cell fate transition during de novo root organogenesis in Arabidopsis. Plant Cell 2014, 26, 1081–1093.
- Mao, J.; Zhang, D.; Meng, Y.; Li, K.; Wang, H.; Han, M. Inhibition of adventitious root development in apple rootstocks by cytokinin is based on its suppression of adventitious root primordia formation. Physiol. Plant 2019, 166, 663–676.
- Mähönen, A.P.; Higuchi, M.; Törmäkangas, K.; Miyawaki, K.; Pischke, M.S.; Sussman, M.R.; Helariutta, Y.; Kakimoto, T. Cytokinins regulate a bidirectional phosphorelay network in Arabidopsis. Curr. Biol. 2006, 16, 1116–1122.
- Atkinson, J.A.; Rasmussen, A.; Traini, R.; Voß, U.; Sturrock, C.; Mooney, S.J.; Wells, D.M.; Bennett, M.J. Branching out in roots: Uncovering form, function, and regulation. Plant Physiol. 2014, 166, 538–550.
- Ramírez-Carvajal, G.A.; Morse, A.M.; Dervinis, C.; Davis, J.M. The cytokinin type-B response regulator PtRR13 is a negative regulator of adventitious root development in Populus. Plant Physiol. 2009, 150, 759–771.
- Jinxiang, W.; Xiaolong, Y.; Ruichi, P. Relationship between adventitious root formation and plant hormones. Plant Physiol. Commun. 2005, 41, 133–142.
- Amiri, E.M.; Elahinia, A. Optimization of medium composition for apple rootstocks. Afr. J. Biotechnol. 2011, 10, 3594–3601.
- Hutchison, C.E.; Li, J.; Argueso, C.; Gonzalez, M.; Lee, E.; Lewis, M.W.; Maxwell, B.B.; Perdue, T.D.; Schaller, G.E.; Alonso, J.M. The Arabidopsis histidine phosphotransfer proteins are redundant positive regulators of cytokinin signaling. Plant Cell 2006, 18, 3073–3087.
- Chen, L.; Tong, J.; Xiao, L.; Ruan, Y.; Liu, J.; Zeng, M.; Huang, H.; Wang, J.-W.; Xu, L. YUCCA-mediated auxin biogenesis is required for cell fate transition occurring during de novo root organogenesis in Arabidopsis. J. Exp. Bot. 2016, 67, 4273–4284.
- Zhao, Y.; Cheng, S.; Song, Y.; Huang, Y.; Zhou, S.; Liu, X.; Zhou, D.-X. The interaction between rice ERF3 and WOX11 promotes crown root development by regulating gene expression involved in cytokinin signaling. Plant Cell 2015, 27, 2469–2483.
- Chandler, J.W.; Werr, W. Cytokinin–auxin crosstalk in cell type specification. Trends Plant Sci. 2015, 20, 291–300.
- Zhao, D.; Wang, Y.; Feng, C.; Wei, Y.; Peng, X.; Guo, X.; Guo, X.; Zhai, Z.; Li, J.; Shen, X. Overexpression of MsGH3. 5 inhibits shoot and root development through the auxin and cytokinin pathways in apple plants. Plant J. 2020, 103, 166–183.
- Da Costa, C.T.; De Almeida, M.R.; Ruedell, C.M.; Schwambach, J.; Maraschin, F.D.S.; Fett-Neto, A.G. When stress and development go hand in hand: Main hormonal controls of adventitious rooting in cuttings. Front. Plant Sci. 2013, 4, 133.
- De Klerk, G.-J.; Hanecakova, J.; Jasik, J. The role of cytokinins in rooting of stem slices cut from apple microcuttings. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2001, 135, 79–84.
- Zimmerman, P.W. Several chemical growth substances which cause initiation of roots and other responses in plants. Contrib. Boyce Thompson Inst. 1935, 7, 209–229.
- Harbage, J.F.; Stimart, D.P. Ethylene does not promote adventitious root initiation on apple microcuttings. J. Am. Soc. Hortic. Sci. 1996, 121, 880–885.
- Růžička, K.; Ljung, K.; Vanneste, S.; Podhorská, R.; Beeckman, T.; Friml, J.; Benková, E. Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell 2007, 19, 2197–2212.
- Harris, J.M. Abscisic acid: Hidden architect of root system structure. Plants 2015, 4, 548–572.
- Petricka, J.J.; Benfey, P.N. Root layers: Complex regulation of developmental patterning. Curr. Opin. Genet. Dev. 2008, 18, 354–361.
- Heidstra, R.; Sabatini, S. Plant and animal stem cells: Similar yet different. Nat. Rev. Mol. Cell Biol. 2014, 15, 301–312.
- Zhang, H.; Han, W.; De Smet, I.; Talboys, P.; Loya, R.; Hassan, A.; Rong, H.; Jürgens, G.; Paul Knox, J.; Wang, M.H. ABA promotes quiescence of the quiescent centre and suppresses stem cell differentiation in the Arabidopsis primary root meristem. Plant J. 2010, 64, 764–774.
- Sriskandarajah, S. Induction of Adventitious Roots in Some Scion Cultivars of Apple (Malus pumila Mill); University of Sydney: Sydney, Australia, 1984.
- Pelese, F.; Megnegneau, B.; Sotta, B.; Sossountzov, L.; Caboche, M.; Miginiac, E. Hormonal characterization of a nonrooting naphthalene-acetic acid tolerant tobacco mutant by an immunoenzymic method. Plant Physiol. 1989, 89, 86–92.
- De Smet, I.; Signora, L.; Beeckman, T.; Inzé, D.; Foyer, C.H.; Zhang, H. An abscisic acid-sensitive checkpoint in lateral root development of Arabidopsis. Plant J. 2003, 33, 543–555.
- Noiton, D.; Vine, J.H.; Mullins, M.G. Endogenous indole-3-acetic acid and abscisic acid in apple microcuttings in relation to adventitious root formation. Plant Growth Regul. 1992, 11, 63–67.
- Zhang, X.; Tahir, M.M.; Li, S.; Tang, T.; Mao, J.; Li, K.; Shao, Y.; Yang, W.; Niu, J.; Zhang, D. Effect of exogenous abscisic acid (ABA) on the morphology, phytohormones, and related gene expression of developing lateral roots in ‘Qingzhen 1’apple plants. Plant Cell Tissue Organ. Cult. (PCTOC) 2022, 148, 23–34.
- Zhang, X.; Tahir, M.M.; Li, S.; Mao, J.; Nawaz, M.A.; Liu, Y.; Li, K.; Xing, L.; Niu, J.; Zhang, D. Transcriptome analysis reveals the inhibitory nature of high nitrate during adventitious roots formation in the apple rootstock. Physiol. Plant 2021, 173, 867–882.
- Li, S.; Tahir, M.M.; Wu, T.; Xie, L.; Zhang, X.; Mao, J.; Ayyoub, A.; Xing, L.; Zhang, D.; Shao, Y. Transcriptome Analysis Reveals Multiple Genes and Complex Hormonal-Mediated Interactions with PEG during Adventitious Root Formation in Apple. Int. J. Mol. Sci. 2022, 23, 976.
- Wasternack, C.; Song, S. Jasmonates: Biosynthesis, metabolism, and signaling by proteins activating and repressing transcription. J. Exp. Bot. 2017, 68, 1303–1321.
- Schilmiller, A.L.; Howe, G.A. Systemic signaling in the wound response. Curr. Opin. Plant Biol. 2005, 8, 369–377.
- Ahkami, A.H.; Lischewski, S.; Haensch, K.T.; Porfirova, S.; Hofmann, J.; Rolletschek, H.; Melzer, M.; Franken, P.; Hause, B.; Druege, U. Molecular physiology of adventitious root formation in Petunia hybrida cuttings: Involvement of wound response and primary metabolism. New Phytol. 2009, 181, 613–625.
- Lischweski, S.; Muchow, A.; Guthörl, D.; Hause, B. Jasmonates act positively in adventitious root formation in petunia cuttings. BMC Plant Biol. 2015, 15, 229.
- Światek, A.; Lenjou, M.; Van Bockstaele, D.; Inzé, D.; Van Onckelen, H. Differential effect of jasmonic acid and abscisic acid on cell cycle progression in tobacco BY-2 cells. Plant Physiol. 2002, 128, 201–211.
- Pauwels, L.; Morreel, K.; De Witte, E.; Lammertyn, F.; Van Montagu, M.; Boerjan, W.; Inzé, D.; Goossens, A. Mapping methyl jasmonate-mediated transcriptional reprogramming of metabolism and cell cycle progression in cultured Arabidopsis cells. Proc. Natl. Acad. Sci. USA 2008, 105, 1380–1385.
- Park, W.J. Melatonin as an endogenous plant regulatory signal: Debates and perspectives. J. Plant Biol. 2011, 54, 143–149.
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin promotes adventitious-and lateral root regeneration in etiolated hypocotyls of Lupinus albus L. J. Pineal Res. 2007, 42, 147–152.
- Sarropoulou, V.; Dimassi-Theriou, K.; Therios, I.; Koukourikou-Petridou, M. Melatonin enhances root regeneration, photosynthetic pigments, biomass, total carbohydrates and proline content in the cherry rootstock PHL-C (Prunus avium × Prunus cerasus). Plant Physiol. Biochem. 2012, 61, 162–168.
- Li, C.; Liang, B.; Chang, C.; Wei, Z.; Zhou, S.; Ma, F. Exogenous melatonin improved potassium content in Malus under different stress conditions. J. Pineal Res. 2016, 61, 218–229.
- Chen, Q.; Qi, W.-b.; Reiter, R.J.; Wei, W.; Wang, B.-m. Exogenously applied melatonin stimulates root growth and raises endogenous indoleacetic acid in roots of etiolated seedlings of Brassica juncea. J. Plant Physiol. 2009, 166, 324–328.
- Pelagio-Flores, R.; Muñoz-Parra, E.; Ortiz-Castro, R.; López-Bucio, J. Melatonin regulates Arabidopsis root system architecture likely acting independently of auxin signaling. J. Pineal Res. 2012, 53, 279–288.
- Pawlicki, N.; Welander, M. The effects of benzyladenine and gibberellic acid on adventitious root formation in apple stem discs. Agronomie 1992, 12, 783–788.
- Müssig, C.; Shin, G.-H.; Altmann, T. Brassinosteroids promote root growth in Arabidopsis. Plant Physiol. 2003, 133, 1261–1271.
- De Klerk, G.-J.; Guan, H.; Huisman, P.; Marinova, S. Effects of phenolic compounds on adventitious root formation and oxidative decarboxylation of applied indoleacetic acid in Malus ‘Jork 9’. Plant Growth Regul. 2011, 63, 175–185.
- James, D.J.; Thurbon, I.J. Phenolic compounds and other factors controlling rhizogenesis in vitro in the apple rootstocks M. 9 and M. 26. Z. Für Pflanzenphysiol. 1981, 105, 11–20.
- Gorter, C.J. Further Experiments on Auxin-Synergists. Physiol. Plant 1962, 15, 88–95.
- Jaleel, C.A.; Riadh, K.; Gopi, R.; Manivannan, P.; Ines, J.; Al-Juburi, H.J.; Chang-Xing, Z.; Hong-Bo, S.; Panneerselvam, R. Antioxidant defense responses: Physiological plasticity in higher plants under abiotic constraints. Acta Physiol. Plant 2009, 31, 427–436.
- Murphy, A.; Peer, W.A.; Taiz, L. Regulation of auxin transport by aminopeptidases and endogenous flavonoids. Planta 2000, 211, 315–324.
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J. Integr. Plant Biol. 2010, 52, 98–111.
- Jasik, J.; De Klerk, G.-J. Anatomical and ultrastructural examination of adventitious root formation in stem slices of apple. Biol. Plant 1997, 39, 79–90.
- Pawlicki, N.; Welander, M. Influence of carbohydrate source, auxin concentration and time of exposure on adventitious rooting of the apple rootstock Jork 9. Plant Sci. 1995, 106, 167–176.
- Calamar, A.; De Klerk, G.-J. Effect of sucrose on adventitious root regeneration in apple. Plant Cell Tissue Organ. Cult. 2002, 70, 207–212.
- Kromer, K.; Gamian, A. Analysis of soluble carbohydrates, proteins and lipids in shoots of M 7 apple rootstock cultured in vitro during regeneration of adventitious roots. J. Plant Physiol. 2000, 156, 775–782.
- Tahir, M.M.; Chen, S.; Ma, X.; Li, S.; Zhang, X.; Shao, Y.; Shalmani, A.; Zhao, C.; Bao, L.; Zhang, D. Transcriptome analysis reveals the promotive effect of potassium by hormones and sugar signaling pathways during adventitious roots formation in the apple rootstock. Plant Physiol. Biochem. 2021, 165, 123–136.
- Wang, Y.-Y.; Hsu, P.-K.; Tsay, Y.-F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467.
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157.
- Tahir, M.M.; Wang, H.; Ahmad, B.; Liu, Y.; Fan, S.; Li, K.; Lei, C.; Shah, K.; Li, S.; Zhang, D. Identification and characterization of NRT gene family reveals their critical response to nitrate regulation during adventitious root formation and development in apple rootstock. Sci. Hortic. 2021, 275, 109642.
- O′Brien, J.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutiérrez, R.A. Nitrate transport, sensing, and responses in plants. Mol. Plant 2016, 9, 837–856.
- Tahir, M.M.; Zhang, X.; Shah, K.; Hayat, F.; Li, S.; Mao, J.; Liu, Y.; Shao, Y.; Zhang, D. Nitrate application affects root morphology by altering hormonal status and gene expression patterns in B9 apple rootstock nursery plants. Fruit Res. 2021, 1, 1–11.
- López-Bucio, J.; Cruz-Ramırez, A.; Herrera-Estrella, L. The role of nutrient availability in regulating root architecture. Curr. Opin. Plant Biol. 2003, 6, 280–287.
- Crawford, N.M.; Glass, A.D. Molecular and physiological aspects of nitrate uptake in plants. Trends Plant Sci. 1998, 3, 389–395.
- Zhang, H.; Jennings, A.; Barlow, P.W.; Forde, B.G. Dual pathways for regulation of root branching by nitrate. Proc. Natl. Acad. Sci. USA 1999, 96, 6529–6534.
- Tahir, M.M.; Li, S.; Mao, J.; Liu, Y.; Li, K.; Zhang, X.; Lu, X.; Ma, X.; Zhao, C.; Zhang, D. High nitrate inhibited adventitious roots formation in apple rootstock by altering hormonal contents and miRNAs expression profiles. Sci. Hortic. 2021, 286, 110230.
- Tahir, M.M.; Lu, Z.; Wang, C.; Shah, K.; Li, S.; Zhang, X.; Mao, J.; Liu, Y.; Shalmani, A.; Li, K. Nitrate Application Induces Adventitious Root Growth by Regulating Gene Expression Patterns in Apple Rootstocks. J. Plant Growth Regul. 2021, 1–12.
- Sriskandarajah, S.; Skirvin, R.; Abu-Qaoud, H. The effect of some macronutrients on adventitious root development on scion apple cultivars in vitro. Plant Cell Tissue Organ. Cult. 1990, 21, 185–189.
- Bhat, K. Nutrient inflows into apple roots. Plant Soil 1983, 71, 371–380.
- Hilo, A.; Shahinnia, F.; Druege, U.; Franken, P.; Melzer, M.; Rutten, T.; von Wirén, N.; Hajirezaei, M.-R. A specific role of iron in promoting meristematic cell division during adventitious root formation. J. Exp. Bot. 2017, 68, 4233–4247.
- Véry, A.-A.; Nieves-Cordones, M.; Daly, M.; Khan, I.; Fizames, C.; Sentenac, H. Molecular biology of K+ transport across the plant cell membrane: What do we learn from comparison between plant species? J. Plant Physiol. 2014, 171, 748–769.
- Römheld, V.; Kirkby, E.A. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180.
- Yamasaki, A.; Yano, T. In Effect of Supplemental Application of Fertilizers on Flower Bud Initiation and Development of Strawberry; Possible Role of Nitrogen. In Proceedings of the VI International Strawberry Symposium 842, Huelva, Spain, 3 March 2008; pp. 765–768.
- Hartz, T.; Johnstone, P.; Francis, D.; Miyao, E. Processing tomato yield and fruit quality improved with potassium fertigation. HortScience 2005, 40, 1862–1867.
- Zhao, Z.; Li, G.; Huang, G. Promotive effect of potassium on adventitious root formation in some plants. Plant Sci. 1991, 79, 47–50.
- Kannan, C.G.; Perumalsamy, P.; Thangavelu, M. Influences of potassium chloride fertilization on mycorrhizal formation in a tropical alfisol. Commun. Soil Sci. Plant Anal. 2017, 48, 524–538.
- Wang, Y.; Wu, W.-H. Potassium transport and signaling in higher plants. Annu. Rev. Plant Biol. 2013, 64, 451–476.

