The biochar biofilter proved to be 96% efficient at cleaning HCH and its transformation products from drainage water, a significant improvement over classic biofilter that remove, on average, 68% of HCH. Although iron- and sulfur-oxidizing bacteria, such as Gallionella and Sulfuricurvum, were dominant in the biochar bed outflows, they were absent in sediments, which were rich in Simplicispira, Rhodoluna, Rhodoferax, and Flavobacterium. The presence of functional genes involved in the biodegradation of HCH isomers and their byproducts was confirmed in both systems. The high effectiveness of the biochar biofilter displayed in this study should further encourage the use of biochar in water treatment solutions, e.g., for temporary water purification installations during the construction of other long-term wastewater treatment technologies, or even as final solutions at contaminated sites.
1. Introduction
Hexachlorocyclohexane (HCH) isomers (especially lindane (γ-1, 2, 3, 4, 5, 6-hexachlorocyclohexane)) are highly persistent toxic compounds, and three of which are included on the Stockholm convention list
[1]. Despite their known toxicity, HCH and its transformation products are virtually ubiquitous in the environment, with background concentrations at around ppb levels in a wide range of matrices
[2][3][4][5][6]. In the 1960s and 1970s, when the toxicity of these substances became apparent, large amounts of waste HCH isomers were disposed of into unsecured subsurface dumps (see
[7] for an inventory of HCH sites from 15 EU countries). Nevertheless, around 382,000 t of technical HCH, and 81,000 t of γ-HCH, were used for agricultural purposes in Europe between 1970 and 1996
[8].
A range of treatment technologies have been suggested for HCH-contaminated areas, and numerous studies focused on the chemical reduction and degradation of HCH isomers have been undertaken
[9][10][11][12][13]. Over the last decade, however, increasing attention has been paid to the use of carbon-rich materials, such as charcoal, bio-coal, and activated carbon, for stabilizing organic pollutants in sediments and soils in situ
[14][15][16]. Biochar, a charcoal produced from plant biomass, is an inexpensive and renewable adsorbent that has been used for a variety of applications, including soil conditioning and remediation, carbon sequestration, water treatment and the absorption of a variety of pollutants
[17]. However, although biochar is rapidly gaining in popularity, only a limited number of studies have been published on the use of biochar for minimizing the bioavailability of pollutants, most of which have been laboratory based
[18][19][20][21], and most concentrating on the factors affecting its ability to remove organic pollutants
[22][23]. One exception was a recent study that sought to restore sediments contaminated by γ-HCH and hexachlorobenzene through amendment with carbon-rich materials, with the aim of sequestering the contaminants and rendering them biologically unavailable
[24]. To date, however, nothing has been published on the use of biochar in constructed wetland systems for biodegradation and removal of pollutants, and particularly HCH isomers, at contaminated dumpsites. Theoretically, HCH accumulating in such biochar biofilters could undergo degradation by indigenous bacteria; however, to the best of our knowledge, nothing is known about the development of microbial consortia in biochar biofilter systems for HCH treatment. By comparison, detailed laboratory studies on aerobic
[25] and anaerobic
[26] microbial degradation of HCH isomers have been undertaken since the 1960s and a number of anaerobic bacteria capable of degrading HCH isomers have been identified, including
Clostridium sphenoids,
Clostridium butyricum,
Clostridium pasteurianum,
Citrobacter freundii,
Desulfovibrio gigas,
Desulfovibrio africanus,
Desulfococcus multivorans,
Dehalobacter sp., and
Clostridium rectum [26], and strains of the Sphingomonadaceae family, capable of aerobic degradation of HCH, have been recorded as dominant at HCH dumpsites worldwide
[27][28][29][30]. The primary reaction during HCH isomer (e.g., lindane) degradation is dehydrochlorination, and functional genes in the genus Sphingobium encoding the key enzymes have now been identified as HCH dehydrochlorinase (LinA), haloalkane dehalogenase (LinB, LinB-RT), and reductive dechlorinase (LinD), with
Sphingomonas paucimobilis UT26 suggested as a possible lindane degradation pathway
[31][32][33]. The dehydrochlorination reactions of γ-, α-, δ-, and β-HCH isomers catalyzed by LinA have also been described
[34]. Most studies have recorded microbial biomass as increasing following biochar addition, with significant changes in microbial community composition and enzyme activity that may explain the biogeochemical effects of biochar. Nevertheless, very little is known about the mechanisms through which biochar affects microbial abundance and community composition. Changes in microbial community composition or activity induced by biochar may affect not only nutrient cycles and plant growth but also the cycling of soil organic matter
[35][36][37].
2. General Characteristics of Inflow Water
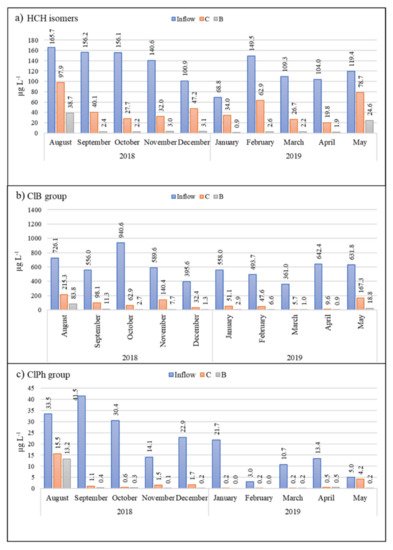
The inflow drainage water had a long-term average pH value of 7.3, with a high conductivity (1350 µS cm−1), high sulfate concentration (650 mg L−1), high total Fe concentration (20 mg L−1), and low F−, NO2−, and NO3− concentrations (below LOD). Average concentrations of HCH, ClB, and ClPh in inflow water were 635.5, 13, and 115.5 µg L−1, respectively (Figure 1).
Figure 1. Monthly measurements of (a) HCH isomers (b) ClB, and (c) ClPh in polluted inflow water (blue), the control biofilter (C, orange), and the biochar biofilter (B, gray) over the course of the experiment.
3. Pollutant Removal Efficiency
After passing through the biochar test bed, 90% of the HCH and other target pollutants had been removed from the drainage water (Figure 1). This already high efficiency was further increased to 96% after the initial high flow rate (15 L min−1) had decreased to 5 L min−1. In comparison, the classically constructed control biofilter with the clay/soil substrate removed around 80% of the HCH (Figure 1).
During analysis, the chemical compounds in outlet water samples of both biofilters were divided into three main groups, i.e., ClB, ClPh, and HCH. Approximately 98% of the total concentration of the ClB group comprised 1.4-diClB > chlorobenzene > 1.3-diClB > 1.2.4-triClB > 1.2-diClB > 1.3.5-triClB, with benzene, 1.2.3-triClB, 1.2.4.5+1.2.3.5-tetraClB, 1.2.3.4-tetraClB, penta-ClB, and hexa-ClB found at <1% of total concentration. In the ClPh group, approximately 89% of total concentration was represented by 4-ClPh > 2-ClPh; 2.4+2.5-diClPh > 3-ClPh > 3.5-diClPh. Finally, the average percentage content of each HCH isomer in the HCH group was δ-HCH (76.1%), ε-HCH (10%), α-HCH (6.7%), γ-HCH (5.1%), and β-HCH (2.1%).
Although we recorded no significant difference (p > 0.05) in outlet water ClB and ClPh concentrations between the control and biochar biofilters, there was a significant difference in HCH concentration (p < 0.01), with an average difference in removal efficiency of 45.3% between the control and biochar beds (see Figure 1), clearly indicating biochar’s greater ability to remove HCH.
In both the control bed and biochar bed, the degradation of pollutants is most likely to have been the result of microbial degradation, with biochar clearly being the more successful of the two methods. Previous studies have shown that the organic pollutant removal efficiency (sorption rate) depends a great deal on its origin (harvest waste, wood, rice husk, sugar industry wastes, etc.), in addition to its surface area, porosity, hydrophobicity, polarity, and pyrolysis temperature
[22][23]. It has been shown, for example, that biochars obtained at temperatures > 500 °C become less polar and more aromatic than those obtained at 300 °C due to the loss of O- and H-containing functional groups
[23]. Because the biochar used in this study was prepared at 570 °C, this would suggest that it had a lower polarity surface and was more aromatic, resulting in decreased sorption.
3.1. HCH Isomers
In the first month of the experiment, the total concentration of HCH in the biochar biofilter decreased by 76.6%, compared with 40.9%in the control (
Figure 1). In both biofilters, a minor decrease was observed in β-HCH and ε-HCH for ten months. These two isomers are most hydrophobic of all the HCH isomers analyzed (
log Kow 3.85 and 4.14, respectively), possibly explaining their low removal rate; however, their degradation rate may also have played a role. A previous study described the HCH isomer degradation rate as γ-HCH > α-HCH > δ-HCH > β-HCH
[38], which partly confirms our own data. However, these authors did not analyze ε-HCH degradation times; hence, we would suggest an augmented sequence of γ-HCH > α-HCH > δ-HCH > β-HCH > ε-HCH. Removal of β-HCH and ε-HCH in the biochar bed reached 91.8% by the fourth month, and continued to increase as time went on, whereas removal of the same two isomers in the control bed was the lowest than all other isomers, reaching just 60.9% and 20.1%, respectively, after ten months. Thus, the decrease in these compounds repeated the tendency of ClB.
Interestingly, the decline in total HCH concentration in the control bed showed a non-linear trend, with percentage removal increasing from 40.9% to 82.2% over the first three months, then decreasing from 82.2% to 50.6% over the next four months, then increasing again from 50.6% to 81% over the next four months (Figure 1). This wave-like dynamic appeared to be dependent on monthly temperature differences, with temperatures ranging from 24.0 to 13.0 °C in the first quarter of the experiment, from 6.0 to 0.7 °C over the second quarter, and from −1.5 to 5.5 °C over the third quarter. In comparison, the concentration of α-HCH decreased by more than 99.7% in the biochar bed over the first ten months (except August), β-HCH by 88.5%, and γ-HCH by 98.4%, or they were not detected at all (α-HCH in 6 months, β-HCH and γ-HCH in 5 months).
3.2. Chlorobenzenes
In the ClB group, the following dynamics was observed: starting from August the efficiency was by 99.5% and 96.6% in control and biochar biofilter, respectively. Moreover, ClB was not detectable in all outflow water samples during the following ten months. The explanation of this pattern could be that chlorobenzene is less hydrophobic (
log Kow 2.84) and volatile within the ClB group. During the following months the most significant decrease in concentrations in all biofilters was for 1.2-diClB (
log Kow 3.43) and 1.2.4-triClB (
log Kow 4.02), whereas a minor decrease was detected for 1.3-diClB (
log Kow is 3.53) and 1.3.5-triClB (
log Kow 4.19). Adsorption of organic compounds by biochar depends not only on the level of hydrophobicity but also on their polarity. Furthermore, there are two different mechanisms for the adsorption of two different types of compounds (polar and hydrophobic). Although the biochar applied to the restoration of the aquatic system was produced at 570 °C and became less polar and more aromatic, polar chemical compounds such as 1.2-diClB (0.91)
[39] and 1.2.4-triClB (0.98)
[40] were better adsorbed by biochar. At the same time, the adsorption and removal of the less polar compounds such as 1.3-diClB (0.85)
[41] and 1.3.5-triClB (0.7)
[42] were relatively smaller. The first month of the experiment (21 August 2018) showed a decrease in the total concentration of ClB in the control biofilter by 70.4%, and in B by 88.5% (
Figure 1). Pairwise comparison evidenced the difference between two biofilters, and the
p-value was <0.001. Unlike the biochar biofilter, the decrease in ClB in the control biofilter exceeded 90% only after four months (December 2018), and at the end of the experiment was 98.5%. This peculiarity can be explained by the drop in average monthly temperature from 24.0 to 3.8 °C and decreasing outflow rate (from 6.2 to 2.3 L min
−1). By comparison, in the biochar biofilter, the decrease was more than 96% by the second month and at the end of the experiment almost reached 100% (99.9%).
Throughout the experiment, the behavior of 1.3.5-triClB in the control biofilter was interesting and might be explained by the fact that this compound is the most hydrophobic within the group (log Kow is 4.19):
The decrease in 1.3.5-triClB concentration remained the smallest during the whole period. Thus, it reduced the decrease percentage of the total concentration of ClB;
-
From August to November, the decrease was oscillatory (58.2%, 28%, 57.4%, and 41%, respectively);
-
From December (8.3%), its removal consistently increased and in April reached 84.4%.
Therefore, soil and clay substrate are not suitable for natural attenuation of high concentrations of 1.3.5-triClB. The biochar biofilter showed better efficiency 98.2% throughout the ten-month experiment.
3.3. Chlorophenols
Five dominant compounds (2-ClPh, 3-ClPh, and 2.4+2.5-ClPh) represent the group of detected ClPh. Total concentration of ClPh in control biofilter decreased within the first month by 53.9% and by 60.8% in biochar wetland (
Figure 2). During the following ten months, the decrease in ClPh total concentration was not lower than 89.7% in the control and 96.4% in the biochar wetland. The smallest decrease was observed for 4-ClPh, which was the only detected ClPh from September in biochar wetland, with the exception in January and February—in these months, it was not detected at all. This tendency can be also explained by the polarity/polarizability level (π) and volatility of the compounds: 4-ClPh is a less polar contaminant within the group (0.72), whereas the π values of 2-ClPh, 3-ClPh, and 2.4+2.5-ClPh are 0.82, 0.77, and 0.87, respectively
[43][44].
4. Microbial Community Structure and Function
The microbial community was analyzed in samples from inflow, control, and biochar biofilters in September 2018 and November 2019. Sediments for molecular biology analyses were taken from both biofilters.
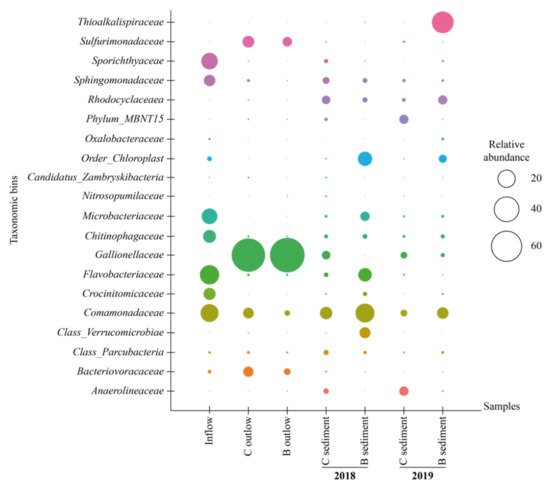
The relative representation of microorganisms at the family level is shown in Figure 3.
Figure 3. Relative abundance (>5%) of microorganisms (family level) in water and sediment samples. B = biochar biofilter, C = control biofilter.
The dominant families in inflow water were Flavobacteriaceae, together with Sporithyaceae and Comamonadaceae. The population of Flavobacterium reached 26.1% of the total biomass in the inflow, whereas in outflow water from the control biofilter its percentage decreased to 0.3% and from the biochar biofilter it was as low as 0.1%. Flavobacterium may be well adapted to higher HCH concentrations due to its possible ability to degrade high concentrations of tech-HCH (25 mg kg
−1) in a microbial consortium containing Pseudomonas spp., Burkholderia, and Vibrio
[45].
The
Gallionellaceae family dominated water outflow samples of control and biochar biofilters (71.2% and 84.6%, respectively) in September 2018, most probably due to the high level of Fe
2+ in the inflow water. Gallionella, Geobacter, and Sulfuricurvum appeared only in outflows due to a decrease in the pollutant’s total concentration. The genus Gallionella is characterized by its chemolithotrophic growth with ferrous iron as a donor of electrons. Chemical dichloroelimination of γ-HCH is relatively pH insensitive and can be mediated by iron sulfides (FeS) and activated carbon to generate di- and trichlorobenzene isomers as end products. Liu et al.
[46] reported that the reducing agent FeS enhanced dichloroelimination of γHCH at a concentration of 10 g L
−1.
The abundance of microorganisms in the control biofilter was very diverse in sediment and water samples (September 2018). In the sediment sample, aryl-halorespiring facultative anaerobic myxobacterium—Anaeromyxobacter was detected, whereas in the water the dominant genus was Sideroxydans, which is able to oxidize Fe
2+ [47].
The abundance of the microbial community changed dramatically after one year. The presence of Galionella increased from 0.9% to 5% in sediment samples in the control biofilter. For the biochar biofilter, Gallionella was not detected in sediment samples in 2018, whereas in 2019, it increased to 1.2%. The abundance of the remaining microbial populations was different between the years. Sediment of the biochar biofilter hosted more Rhodocyclaceae (7.1%) than that of the control biofilter (1.5%). Dadwhal et al.
[48] isolated Rhodocyclaceae from HCH-contaminated soil and suggested that members of this family may be involved in the removal of HCH.
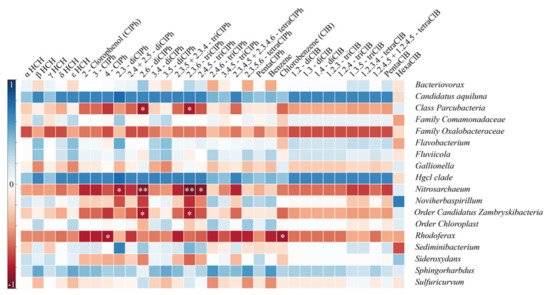
Spearman correlation analysis demonstrated a negative correlation between several families and chemical compounds (
Figure 3). Parcubacteria and ZhambryskiiBacteria families negatively correlated with 2.6-diClPh and 2.3.6-triClPh (
p < 0.05), i.e., while these two compounds’ concentrations increase, the abundance of Parcubacteria and ZhambryskiiBacteria families decreases. Nitrosarchaeum had a stronger negative correlation with the above compounds (
p < 0.05), whereas its correlation with 2.3-diClPh and 2.4.5-triClPh was significant but weaker (
p < 0.05). The Rhodoferax family was negatively correlated with ClB and 4-ClPh (
p < 0.05). Thus, all detected correlations were negative and three of five families correlated with the 2.6-diClPh and 2.3.6-triClPh. Interestingly, three bacterial groups positively (although not significantly) correlated with all targeted pollutants.
Candidatus Aquiluna (Microbacteriaceae) has been reported from a wide range of aquatic habitats from freshwaters to hypersaline waters
[49]. Hgcl clade (Actinobacteria), was associated with soil and eutrophic waters and is able to use organic compounds for their metabolism and, most importantly,
Sphingorhabdus is able to metabolize HCH.
Figure 3. Spearman correlations for taxa to chemical compounds ** p < 0.01, * p < 0.05.
5. Functional Genes Involved in HCH Biodegradation
The functional genes
linA, linB, linB-RT, and
linD encoding enzymes involved in aerobic and anaerobic γ-HCH biodegradation were detected in all sediment samples except those taken from the freshly installed biochar biofilter in 2018 (
Table 1), suggesting that the microbial community was still not well developed one month after installation. In comparison, the highest levels of
U16SRT, linA, LinB-RT, and
LinD genes were detected in sediment from the control bed in 2018. However, it was the opposite in samples from the same biofilter in 2019, except for the
U16SRT gene. There is clear experimental evidence that LinA dehydrochlorizes γ-, α- and δ-HCH, whereas LinB hydrolytically dechlorines β- and δ-HCH in all strains studied
[25]. Within HCH isomers, the γ-HCH (lindane) displaying the fastest degradation rate.
Table 1. Relative abundance of genes indicating total bacterial biomass (16S rDNA), dehydrochlorinase (linA), haloalkane dehalogenase (linB, linB-RT), and reductive dechlorinase (linD) in the control biofilter (C, average of duplicate samples) and biochar sediments (B, average of duplicate samples). The color scale indicates the relative quantity of a given marker: red (+++) highest, orange (++) high, and yellow (+) intermediate quantity, ND = not detected or below the LOQ.
| |
|
Gene |
| Biofilter |
U16SRT |
linA |
linB |
linB-RT |
linD |
| 2018 |
C |
+++ |
+++ |
++ |
+++ |
+++ |
| B |
+ |
ND |
ND |
ND |
ND |
| 2019 |
C |
+++ |
++ |
++ |
++ |
++ |
| B |
+++ |
+++ |
+++ |
+++ |
+++ |
The linB gene was detected in large quantities in all samples except in early biochar samples from 2018. The largest quantity of this gene was in sediment samples from biochar biofilter in 2019. The largest quantity of the linB-RT gene was found in sediment samples from the control biofilter in 2018 and sediment from biochar biofilter in 2019. In 2018, linD genes were most abundant in sediment samples taken from the control biofilter, but in 2019 the quantity was decreased, unlike in biochar biofilter, for which its quantity was greatest.
linD encoding reductive dechlorinase was first isolated from
Sphingobium japonica UT26
[50]. Dehalogenation of HCH isomers and byproducts is also a significant reaction. LinB and linB-RT encode haloalkane dehalogenase and catalyzes the HCH hydrolytic dechlorination of 1.4-TCDN to 2.5-DDOL
[51].
Overall, in 2019, lin genes were found in all samples, and the maximum quantity was found in samples taken from the biochar biofilter in 2019. Therefore, the removal of HCH isomers and their byproducts was not driven by sorption on biochar, but by biodegradation.
 +1 credit
+1 credit