Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Antonio Pineda-Lucena | + 2794 word(s) | 2794 | 2021-09-13 11:03:19 | | | |
| 2 | Camila Xu | Meta information modification | 2794 | 2021-09-24 08:40:39 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Pineda-Lucena, A. Multi-Omic Approaches to Breast Cancer. Encyclopedia. Available online: https://encyclopedia.pub/entry/14510 (accessed on 24 June 2026).
Pineda-Lucena A. Multi-Omic Approaches to Breast Cancer. Encyclopedia. Available at: https://encyclopedia.pub/entry/14510. Accessed June 24, 2026.
Pineda-Lucena, Antonio. "Multi-Omic Approaches to Breast Cancer" Encyclopedia, https://encyclopedia.pub/entry/14510 (accessed June 24, 2026).
Pineda-Lucena, A. (2021, September 24). Multi-Omic Approaches to Breast Cancer. In Encyclopedia. https://encyclopedia.pub/entry/14510
Pineda-Lucena, Antonio. "Multi-Omic Approaches to Breast Cancer." Encyclopedia. Web. 24 September, 2021.
Copy Citation
Breast cancer (BC) is the most frequently diagnosed tumor and the leading cause of cancer deaths in women worldwide.
metabolism
breast cancer
multi-omics
biomarkers
early diagnosis
1. Introduction
Breast cancer (BC) is the most frequently diagnosed tumor and the leading cause of cancer deaths in women worldwide [1]. The year 2020 saw an estimated 2.3 million new cases of BC (11.7% of all cancer cases), with 685,000 deaths worldwide [1]; however, advances in population screening and early treatment (among other factors) have supported a steady decrease in BC mortality [2][3]. Unfortunately, figures from the American Cancer Society place the five-year survival rate after diagnosis of metastatic BC at 27%, a low value considering the 99% five-year survival rate for localized disease [4]. Therefore, early BC detection represents a crucial step in reducing disease mortality [5]. BC screening currently relies on mammography, a non-invasive strategy primarily performed in women between 50 to 69 years of age that has prompted a reduction in BC mortality [6]. Nevertheless, this approach suffers from several limitations, including false-positive reporting and overdiagnosis [6][7][8][9][10]. Ultrasound, magnetic resonance imaging, and computed tomography can overcome such problems thanks to their high sensitivity; however, the elevated costs associated with these tools make this approach less accessible. Thus, we still lack alternative methods for the accurate, non-invasive, and low-cost diagnosis of early-stage BC.
BC is a highly heterogeneous disease from a molecular perspective and is primarily characterized by the overexpression of the HER2 growth factor, estrogen receptor (ER), and progesterone receptor (PR) and mutations in the BRCA1/2 genes, with the latter associated with a higher risk of developing BC [11]. Former classifications of BC tumors employed tumor size, histological grade, immunohistochemistry of ER/PR status, and the amplification of HER2. The addition of gene expression profiling to these molecular features has resulted in the classification currently used by the European Society for Medical Oncology as a clinical guideline for BC diagnosis, follow-up, and treatment [12][13]. This system classifies BC tumors into four major intrinsic molecular subgroups: luminal A (ER+ and/or PR+, HER2-, low Ki67), luminal B (ER+ and/or PR+, HER2+ or HER2- with high Ki67), basal-like (ER/PR-, HER2-), and HER2-enriched (ER/PR-, HER2+). Luminal A tumors (the low-grade group) are the most common BC subtype, comprising over 60–70% of all cases. Meanwhile, basal-like tumors, with an ~80% overlap with highly proliferative triple-negative breast cancer (TNBC) [14], exhibit aggressive behavior and suffer from poor prognosis [15]. Although ER+ tumors present lower recurrence rates within the first five years, over 50% of tumor recurrences occur after this time and cause most BC-related deaths [16][17]. Each BC subtype has a characteristic biological profile, prognosis, and treatment strategy [18][19][20][21][22], and several scoring systems aid prognosis and treatment decision-making processes. Unfortunately, systems based on different molecular features related to tumor biology, including histological type, grade, lymphovascular invasion, and marker status, do not accurately reflect BC subtype heterogeneity or specific patient subtypes [23]. Thus, enormous efforts have been devoted to classifying heterogeneous BC subtypes into molecular subtypes that guide treatment decisions [24][25][26][27][28].
2. Multi-Omics Studies of BC Prognosis
Metabolic deregulation can impact various molecular processes (e.g., cell proliferation, apoptosis, migration, and invasion) that contribute to tumor progression [29][30][31][32] and influence cancer patient survival (Figure 2) [33].
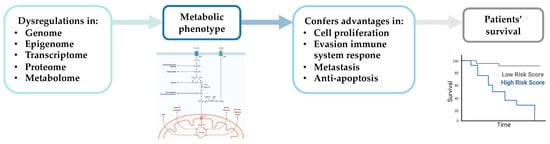
Figure 2. Schematic representation of the impact of metabolic changes on essential molecular processes associated with tumor progression and patient survival. Created with BioRender.com.
Several studies have reported associations between metabolic alterations and BC patient survival based on single omic analysis, including genomic [34], transcriptomic [35], proteomic [36], and metabolomic [37][38][39] studies in tissue [36][34][35] and serum [37][38][39] samples. However, additional studies based on the integration of data from multi-omics analyses have provided more accurate information regarding the molecules involved in metabolic rewiring associated with BC progression. Of the seven studies following a multi-omics approach to identify metabolic alterations associated with BC prognosis (Table 3), most relied on the analysis of tissue sample analysis and the integration of transcriptomic and metabolomic datasets. Overall, BC patient survival associated with the altered expression of enzymes involved in nucleotide, lipid, and amino acid metabolism.
Table 3. Multi-omics studies focused on identifying metabolic alterations associated with BC prognosis.
| Study | Sample | Omics Data | Major Findings * |
|---|---|---|---|
| Putluri et al. [40] | Cell lines + Tissue | M+T | ↑ RRM2 (pyrimidine metabolism) |
| Luo et al. [41] | Blood + Tissue | M+T | ↑ RRM2 (pyrimidine metabolism) and ↓ AMPD1 (de novo purine metabolism) |
| Iqbal et al. [42] | Tissue | M+T | ↑ CBX2 and ↓ CBX7 (glycolysis) |
| Camarda et al. [43] | Cell lines + Tissue | M+T | ↓ ACC2 (FAO) |
| Kang et al. [44] | Cell lines | L+T | ↓ ELOVL2 (lipid synthesis) |
| Terunuma et al. [45] | Tissue | M+T+P+E | ↑ 2HG, SAM and SAH ↑ IDH2 (glutamine metabolism) |
| Budczies et al. [46] | Tissue | M+T | ↓ ABAT, ↑ β-alanine (β-alanine metabolism) |
ABAT: 4-aminobutyrate aminotransferase, ACC2: acetyl-CoA carboxylase 2, AMPD1: adenosine monophosphate deaminase 1, CBX2: chromobox 2, CBX7: chromobox 7, E: epigenomics, ELOVL2: ELOVL fatty acid elongase 2, FAO: fatty acid oxidation, L: lipidomics, IDH2: isocitrate dehydrogenase (NADP(+)) 2, M: metabolomics, P: proteomics, RRM2: ribonucleotide reductase regulatory subunit M2, SAH: S-adenosylhomocysteine, SAM: S-adenosyl- methionine, T: transcriptomics, 2HG: 2-hydroxyglutarate. * Direction of metabolic alterations directly correlated with worse BC patients’ outcomes.
Previous studies have demonstrated that nucleotide biosynthesis plays a vital role in BC [47][48][49][50][51], and could represent a promising therapeutic strategy [52][53][54]. Notably, two of the multi-omics studies included in Table 3 established an inverse correlation between the expression levels of genes involved in de novo purine and pyrimidine syntheses and BC patient survival [41][40]. Putluri et al. performed an in silico analysis to evaluate the association between omics-based enrichment and patient survival, using ten independent gene expression data sets to select clinically relevant prognostic biomarkers [40]. Kaplan–Meyer curves revealed an association between increased expression of pyrimidine metabolism-related genes and shorter metastasis-free survival across all BC and within the subset of ER + tumors. RRM2, a critical gene in pyrimidine metabolism displaying elevated expression in aggressive BC [55], has prognostic relevance in BC [56][57] when combined with proliferation markers [58]. RRM2 expression distinguished good vs. poor survival within the entire BC patient group in this multi-omics study, including a significant proportion of luminal A subtype typically considered to have better survival outcomes [48]. Luo and colleagues integrated metabolomic and transcriptomic analysis to confirm the association between alterations in nucleotide metabolism and BC patient survival in a TCGA cohort of patients [41]. The authors observed a significant correlation between poor survival of BC patients and changes to the expression of RRM2 and adenosine monophosphate deaminase 1 (AMPD1), a key enzyme in de novo purine synthesis. These enzymes have been postulated as promising therapeutic targets in different tumor types, including BC [40][53][59][60][61][62].
Iqbal et al. established antagonistic roles of CBX2 and CBX7 in metabolic reprogramming of BC and an association with BC patient survival [42]. The authors described a significant correlation between higher CBX2 and lower CBX7 mRNA levels and worse BC prognosis, which agrees with previous findings that correlated the CBX2 or CBX7 expression with overall patient survival [63][64][65][66].
As for the alterations in lipid metabolism, Camarda and colleagues followed a targeted metabolomics approach and reported the dramatic upregulation of FAO intermediates in a MYC-driven model of TNBC [43]. To characterize a potential association between FAO gene expression and prognosis in TNBC, the authors performed a univariate analysis of 336 fatty acid metabolism genes on a patient cohort with long-term distant recurrence-free survival data. The analysis revealed that decreased ACACB (acetyl-CoA carboxylase 2, ACC2) expression levels associated with worse prognoses in all BC and TNBC patients. Subsequent studies also described significant associations between increased levels of ACC2 and better BC prognosis [67][68][69][70]. Kang et al. conducted a multi-layered lipidomics and transcriptomics analysis to describe the rewiring of the BC lipidome during malignant transformation [44]. Analyses in a spheroid-induced epithelial-mesenchymal transition (EMT) model demonstrated a dramatic reduction in the ratio of C22:6n3 (docosahexaenoic acid, DHA) to C22:5n3 in spheroid cells, similarly to the down-regulation of ELOVL2, a process associated with the induction of metastatic characteristics in BC cells. The authors examined the relationship between ELOVL2 expression and metastatic relapse-free in a BC cohort with a follow-up of ten years, resulting in the discovery of an association between lower ELOVL2 expression levels and shorter metastasis-free survival and higher tumor grade [44]. A recent study investigating the molecular mechanisms of tamoxifen resistance in BC confirmed these findings and described lower ELOVL2 expression in tamoxifen-resistant models and ELOVL2 downregulation in patients with tamoxifen resistance [71].
Additional studies have revealed a correlation between alterations in the levels of genes and metabolites involved in amino acid metabolism and BC prognosis. Terunuma and coworkers identified a subset of BC tumors accumulating high levels of 2-hydroxyglutarate (2HG). Further analyses revealed the presence of a subgroup of BC patients with significantly decreased survival characterized by an exceptionally high accumulation of 2HG, reduced DNA methylation at the isocitrate dehydrogenase (IDH2) locus, increased IDH2 expression, and increased levels of S-adenosyl- methionine (SAM) and S-adenosylhomocysteine (SAH) [45]. Previous studies reported an accumulation of the oncometabolite 2HG in different tumor types, including BC [72], glioma [73], and leukemia [74]. In another study, concerning amino acid metabolism, Budczies et al. described an association between alterations in the metabolism of β-alanine and shorter recurrence-free survival of BC patients [46]. Specifically, the authors demonstrated that lower expression levels of 4-aminobutyrate aminotransferase (ABAT), which negatively correlated with the concentration of β-alanine, indicated worse prognoses in BC patients. A similar study reported decreased ABAT expression in more aggressive BC subtypes, which correlated with an increased risk of metastasis and shorter overall, relapse-free, and distant metastasis-free survival [75]. Finally, Jansen et al. correlated ABAT downregulation with poor progression-free and metastasis-free survival in tamoxifen-treated patients [76].
3. Multi-Omics Studies and Novel BC Treatment Strategies
Omics-based technologies have also been used to identify novel therapeutic targets and monitor biological alterations related to BC metabolism following treatment [77][78][79][80][81][82][83][84][85]. The majority of studies relied on the application of metabolomics-based approaches in BC tissue [80][82] and serum [79][81][83] samples, although groups have evaluated transcriptomic [86][78] and proteomic [85] profiles in tissue samples. Various studies have described how the combination of omics approaches could characterize specific targets and foster the development of novel therapeutic strategies for specific subgroups of BC patients [87][88][89]. In particular, multi-omics studies have focused on identifying and validating metabolic enzymes as promising therapeutic strategies for the treatment of different BC tumors (Table 4). Figure 3 illustrates those metabolic-related genes proposed as potential therapeutic targets for treating BC patients in these studies. Overall, these findings suggest the therapeutic potential of inhibiting specific metabolic enzymes associated with glycolysis or involved in nucleotide, amino acid, and lipid metabolism in BC patients.
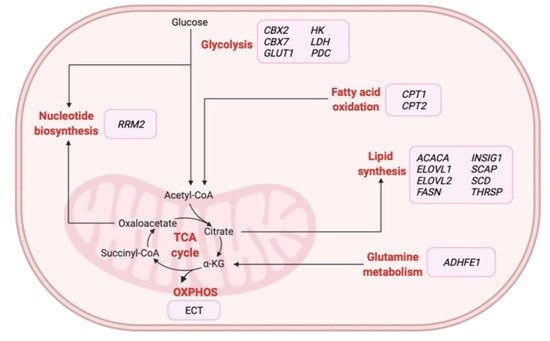
Figure 3. Overview of metabolic-related therapeutic targets for the treatment of BC patients identified from multi-omics-based studies. α-KG: alpha-ketoglutarate, ACACA: acetyl-CoA carboxylase alpha, ADHFE1: alcohol dehydrogenase iron containing 1, CBX2: chromobox 2, CBX7: chromobox 7, CPT1: carnitine palmitoyltransferase 1A, CPT2: carnitine palmitoyltransferase 2, ECT: electron transport chain, ELOVL1: ELOVL fatty acid elongase 1, ELOVL2: ELOVL fatty acid elongase 2, FASN: fatty acid synthase, GLUT1: glucose transporter 1, HK: hexokinase, INSIG1: insulin-induced gene 1, LDH: lactate dehydrogenase, OXPHOS: oxidative phosphorylation, PDC: pyruvate dehydrogenase kinase, RRM2: ribonucleotide reductase regulatory subunit M2, SCAP: SREBF chaperone, SCD: stearoyl-CoA desaturase, TCA: tricarboxylic acid, THRSP: thyroid hormone-responsive. Created with BioRender.com.
Table 4. Multi-omics studies focused on developing new therapeutic strategies for the treatment of BC.
| Study | Omics Data | BC Subtype | Potential Targets |
|---|---|---|---|
| Iqbal et al. [42] | M+T | TNBC and luminal-like | CBX2 and CBX7 |
| Gong et al. [90] | M+T | TNBC | FASN and LDH |
| Mahendralingam et al. [91] | P+T | Basal- and luminal-like | GLUT1, HK, LDH, and PDC |
| Putluri et al. [40] | M+T | Basal- and luminal-like | RRM2 |
| Terunuma et al. [45] | M+T+P+E | TNBC and basal-like | ADHFE1 |
| Hilvo et al. [92] | L+T | TNBC, luminal- and basal-like |
ACACA, ELOVL1, FASN, INSIG1, SCAP, SCD and THRSP |
| Kang et al. [44] | L+T | Luminal-like | ELOVL2 |
| Camarda et al. [43] | M+T | TNBC and HER2 + | CPT1 and CPT2 |
ACACA: acetyl-CoA carboxylase alpha, ADHFE1: alcohol dehydrogenase iron containing 1, CBX2: chromobox 2, CBX7: chromobox 7, CPT1: carnitine palmitoyltransferase 1A, CPT2: carnitine palmitoyltransferase 2, E: epigenomics, ELOVL1: ELOVL fatty acid elongase 1, ELOVL2: ELOVL fatty acid elongase 2, FASN: fatty acid synthase, GLUT1: glucose transporter 1, HK: hexokinase, IDH2: isocitrate dehydrogenase (NADP(+)) 2, INSIG1: insulin-induced gene 1, L: lipidomics, LDH: lactate dehydrogenase, M: metabolomics, P: proteomics, PDC: pyruvate dehydrogenase kinase, RRM2: ribonucleotide reductase regulatory subunit M2, SCAP: SREBF chaperone, SCD: stearoyl-CoA desaturase, T: transcriptomics, TNBC: triple negative breast cancer, THRSP: thyroid hormone-responsive.
Iqbal and coworkers demonstrated that silencing CBX2 and CBX7 exhibited inverse effects on glycolysis, ATP production, viability, and proliferation [59]. CBX7 overexpression provided comparable results to CBX2 knockdown, which included decreased biomass production and reduced cell viability and proliferation. These in vitro results agreed with the findings of the transcripto-metabolomic analyses performed on BC patients and validated the roles of CBX2 and CBX7 in metabolic reprogramming of BC, highlighting the potential of these targets for the development of therapeutic strategies in BC. Of note, additional studies have provided similar results for CBX2 and CBX7 in BC [66][93][94], pancreatic adenocarcinoma [64] and metastatic prostate cancer [95]. Gong and coworkers evaluated the sensitivity of different BC metabolic phenotypes to metabolic inhibitors targeting glycolysis or de novo fatty acid synthesis [90]. The glycolytic BC phenotype displayed greater sensitivity to glycolytic inhibitors (oxamate, lactate dehydrogenase (LDH) inhibitors, and 2-deoxy-D-glucose), while inhibitors of lipid synthesis (cerulin and FASN inhibitor) exhibited higher efficacy against the lipogenic phenotype. Significantly, in vivo LDH inhibition enhanced tumor response to anti-PD-1 immunotherapy in the BC glycolytic phenotype. Previous studies have shown that LDH inhibition can suppress glycolysis [96][97] and cell proliferation [98] in BC cell lines. Differences in sensitivity to pharmacological inhibitors targeting glycolysis or electron transport chain (ECT) subunits were evaluated in different metabolic MECs phenotypes by Mahendralingam and colleagues [91]. In this study, the glycolytic phenotype displayed greater sensitivity to inhibitors targeting glucose transporter 1 (GLUT1), hexokinase (HK), LDH, and pyruvate dehydrogenase kinase (PDC), which agrees with results reported by Gong et al. [90]. Furthermore, studies have demonstrated that HK inhibition prevents BC growth [99][100].
The metabolic enzyme RRM2 has also been proposed as a potential therapeutic target for the treatment of BC. Putluri et al. observed that inhibiting RRM2 in BC cells significantly decreased proliferation and the expression of cell cycle genes and sensitized cells to tamoxifen treatment [40]. In agreement with the potential relevance of RRM2, additional studies have reported a reduction in proliferation [53][62] and tamoxifen resistance [101] in BC cell lines following RRM2 inhibition. Furthermore, associations between RRM2 overexpression and deterioration in BC survival have been widely reported, strongly suggesting a role as a targeted therapy for BC [41][56][57].
A multi-omics-based study by Terunuma et al. identified alterations associated with glutamine metabolism in a subset of BC tumors [45]. The authors described a subset of BC tumors with high 2HG levels and a distinct DNA methylation pattern associated with worse prognoses. Overall, studies have underscored the critical role of epigenetic-metabolomic interplay in promoting tumorigenesis [102]; in particular, high 2-HG levels induce epigenetic reprogramming associated with progression in different tumors [103][104]. Interestingly, the silencing of IDH2 and alcohol dehydrogenase iron containing 1 (ADHFE1), two enzymes implicated in the mitochondria-associated α-ketoglutarate–dependent production of 2HG [74][105], prompted a marked reduction of endogenous 2HG in BC cells. Furthermore, ADHFE1 loss resulted in a moderate but significant inhibition of cell cycle kinetics and reduced migration and invasion, suggesting an oncogenic role for ADHFE1 in BC. In agreement, several studies have associated high ADHFE1 expression levels with increased synthesis of 2HG and worse patient prognosis in BC [106][107].
In the context of a multi-omics study focused on lipid metabolism, Hilvo et al. conducted gene silencing experiments on seven enzymes involved in phospholipid remodeling and de novo lipid synthesis [92]. The results established that the individual inhibition of multiple lipid metabolism-regulating genes reduced the growth and viability of BC cell lines, which agrees with studies reporting reduced cell migration, invasion, and tumor proliferation in BC [108][109][110] and other tumor types [111][112][113] following the inhibition of specific lipid metabolism-related enzymes. Interestingly, a more recent study described FASN, another enzyme involved in de novo lipid metabolism, as a promising therapeutic target for BC treatment [114].
The results of a multi-omics-based study conducted by Kang et al. revealed lipid composition alterations during the EMT in BC [44]. The inhibition of ELOVL2 increased malignant potential, higher migration rate, and elevated colony formation. Mechanistically, downregulation of ELOVL2 increased sterol regulatory element-binding transcription factor 1 (SREBP1) expression in BC cells and activated lipogenesis, a process associated with the promotion of malignant BC phenotypes. SREBP1 is a crucial regulator of fatty acid metabolism and plays a pivotal role in the transcriptional regulation of different lipogenic genes mediating lipid synthesis [115][116]. SREBP1 overexpression has been observed in different tumor types, including BC [92][117][118], and supports the malignant BC phenotype [110]. Finally, Camarda et al. suggested the inhibition of FAO as a novel therapeutic target in a MYC-driven model of TNBC [43]. Based on the characterization of the effects of small-molecule inhibition and knock-down of carnitine palmitoyltransferase 1A (CPT1) and carnitine palmitoyltransferase 2 (CPT2) in BC cell lines, the authors demonstrated that FAO plays an essential role in this BC model. Furthermore, in vivo experiments demonstrated that treatment with etomoxir, a CPT1 inhibitor, significantly attenuated tumor growth in various BC models. Additional studies have also demonstrated that individual knockdown of both CPT enzymes reduces FAO metabolism and cell proliferation in different tumors [119][120][121]. In addition, more recent studies suggested CPT1 as a potential BC tumor target [122][123].
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249.
- Berry, D.A.; Cronin, K.A.; Plevritis, S.K.; Fryback, D.G.; Clarke, L.; Zelen, M.; Mandelblatt, J.S.; Yakovlev, A.Y.; Habbema, J.D.F.; Feuer, E.J. Effect of screening and adjuvant therapy on mortality from breast cancer. N. Engl. J. Med. 2005, 353, 1784–1792.
- Plevritis, S.K.; Munoz, D.; Kurian, A.W.; Stout, N.K.; Alagoz, O.; Near, A.M.; Lee, S.J.; Van den Broek, J.J.; Huang, X.; Schechter, C.B.; et al. Association of screening and treatment with breast cancer mortality by molecular subtype in US women, 2000–2012. JAMA 2018, 319, 154–164.
- Survival Rates for Breast Cancer. Available online: https://www.cancer.org/cancer/breast-cancer/understanding-a-breast-cancer-diagnosis/breast-cancer-survival-rates.html (accessed on 23 June 2021).
- Li, J.; Guan, X.; Fan, Z.; Ching, L.-M.; Li, Y.; Wang, X.; Cao, W.-M.; Liu, D.-X. Non-invasive biomarkers for early detection of breast cancer. Cancers 2020, 12, 2767.
- Independent UK panel on breast cancer screening the benefits and harms of breast cancer screening: An independent review. Lancet 2012, 380, 1778–1786.
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Benbrahim-Tallaa, L.; Bouvard, V.; Bianchini, F.; Straif, K. International agency for research on cancer handbook working group breast-cancer screening—Viewpoint of the IARC working group. N. Engl. J. Med. 2015, 372, 2353–2358.
- Houssami, N. Overdiagnosis of breast cancer in population screening: Does it make breast screening worthless? Cancer Biol. Med. 2017, 14, 1–8.
- Nelson, H.D.; Pappas, M.; Cantor, A.; Griffin, J.; Daeges, M.; Humphrey, L. Harms of breast cancer screening: Systematic review to update the 2009 U.S. preventive services task force recommendation. Ann. Intern. Med. 2016, 164, 256.
- Carter, J.L.; Coletti, R.J.; Harris, R.P. Quantifying and monitoring overdiagnosis in cancer screening: A systematic review of methods. BMJ 2015, 350, g7773.
- Brewer, H.R.; Jones, M.E.; Schoemaker, M.J.; Ashworth, A.; Swerdlow, A.J. Family history and risk of breast cancer: An analysis accounting for family structure. Breast Cancer Res. Treat. 2017, 165, 193–200.
- Cardoso, F.; Kyriakides, S.; Ohno, S.; Penault-Llorca, F.; Poortmans, P.; Rubio, I.T.; Zackrisson, S.; Senkus, E. ESMO Guidelines Committee. Early breast cancer: ESMO clinical practice guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2019, 30, 1194–1220.
- Cardoso, F.; Senkus, E.; Costa, A.; Papadopoulos, E.; Aapro, M.; André, F.; Harbeck, N.; Aguilar Lopez, B.; Barrios, C.H.; Bergh, J.; et al. 4th ESO-ESMO international consensus guidelines for advanced breast cancer (ABC 4). Ann. Oncol. 2018, 29, 1634–1657.
- Network, T.C.G.A. Cancer genome atlas network comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70.
- Milioli, H.H.; Tishchenko, I.; Riveros, C.; Berretta, R.; Moscato, P. Basal-like breast cancer: Molecular profiles, clinical Features and survival outcomes. BMC Med. Genom. 2017, 10, 19.
- Anderson, W.F.; Chen, B.E.; Jatoi, I.; Rosenberg, P.S. Effects of estrogen receptor expression and histopathology on annual hazard rates of death from breast cancer. Breast Cancer Res. Treat. 2006, 100, 121–126.
- Goncalves, R.; Ma, C.; Luo, J.; Suman, V.; Ellis, M.J. Use of neoadjuvant data to design adjuvant endocrine therapy trials for breast cancer. Nat. Rev. Clin. Oncol. 2012, 9, 223–229.
- Rouzier, R.; Perou, C.M.; Symmans, W.F.; Ibrahim, N.; Cristofanilli, M.; Anderson, K.; Hess, K.R.; Stec, J.; Ayers, M.; Wagner, P.; et al. Breast cancer molecular subtypes respond differently to preoperative chemotherapy. Clin. Cancer Res. 2005, 11, 5678–5685.
- Sotiriou, C.; Neo, S.-Y.; McShane, L.M.; Korn, E.L.; Long, P.M.; Jazaeri, A.; Martiat, P.; Fox, S.B.; Harris, A.L.; Liu, E.T. Breast cancer classification and prognosis based on gene expression profiles from a population-based study. Proc. Natl. Acad. Sci. USA 2003, 100, 10393–10398.
- Van’t Veer, L.J.; Dai, H.; Van de Vijver, M.J.; He, Y.D.; Hart, A.A.M.; Mao, M.; Peterse, H.L.; Van der Kooy, K.; Marton, M.J.; Witteveen, A.T.; et al. Gene expression profiling predicts clinical outcome of breast cancer. Nature 2002, 415, 530–536.
- Carey, L.A.; Dees, E.C.; Sawyer, L.; Gatti, L.; Moore, D.T.; Collichio, F.; Ollila, D.W.; Sartor, C.I.; Graham, M.L.; Perou, C.M. The triple negative paradox: Primary tumor chemosensitivity of breast cancer subtypes. Clin. Cancer Res. 2007, 13, 2329–2334.
- Huang, E.; Cheng, S.H.; Dressman, H.; Pittman, J.; Tsou, M.H.; Horng, C.F.; Bild, A.; Iversen, E.S.; Liao, M.; Chen, C.M.; et al. Gene expression predictors of breast cancer outcomes. Lancet 2003, 361, 1590–1596.
- Phung, M.T.; Tin Tin, S.; Elwood, J.M. Prognostic models for breast cancer: A systematic review. BMC Cancer 2019, 19, 230.
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767.
- Burstein, M.D.; Tsimelzon, A.; Poage, G.M.; Covington, K.R.; Contreras, A.; Fuqua, S.A.W.; Savage, M.I.; Osborne, C.K.; Hilsenbeck, S.G.; Chang, J.C.; et al. Comprehensive genomic analysis identifies novel subtypes and targets of triple-negative breast cancer. Clin. Cancer Res. 2015, 21, 1688–1698.
- Bareche, Y.; Venet, D.; Ignatiadis, M.; Aftimos, P.; Piccart, M.; Rothe, F.; Sotiriou, C. Unravelling triple-negative breast cancer molecular heterogeneity using an integrative multiomic analysis. Ann. Oncol. 2018, 29, 895–902.
- Jiang, Y.-Z.; Ma, D.; Suo, C.; Shi, J.; Xue, M.; Hu, X.; Xiao, Y.; Yu, K.-D.; Liu, Y.-R.; Yu, Y.; et al. Genomic and transcriptomic landscape of triple-negative breast cancers: Subtypes and treatment strategies. Cancer Cell 2019, 35, 428–440.e5.
- Garrido-Castro, A.C.; Lin, N.U.; Polyak, K. Insights into molecular classifications of triple-negative breast cancer: Improving patient selection for treatment. Cancer Discov. 2019, 9, 176–198.
- Wang, L.; Zhang, S.; Wang, X. The Metabolic mechanisms of breast cancer metastasis. Front. Oncol. 2021, 10, 602416.
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 28.
- Wei, Q.; Qian, Y.; Yu, J.; Wong, C.C. Metabolic rewiring in the promotion of cancer metastasis: Mechanisms and therapeutic implications. Oncogene 2020, 39, 6139–6156.
- Kreuzaler, P.; Panina, Y.; Segal, J.; Yuneva, M. Adapt and conquer: Metabolic flexibility in cancer growth, invasion and evasion. Mol. Metab. 2020, 33, 83–101.
- Läsche, M.; Emons, G.; Gründker, C. Shedding new light on cancer metabolism: A metabolic tightrope between life and death. Front. Oncol. 2020, 10, 409.
- Li, C.; Li, X.; Li, G.; Sun, L.; Zhang, W.; Jiang, J.; Ge, Q. Identification of a prognosis-associated signature associated with energy metabolism in triple-negative breast cancer. Oncol. Rep. 2020, 44, 819–837.
- Feng, J.; Ren, J.; Yang, Q.; Liao, L.; Cui, L.; Gong, Y.; Sun, S. Metabolic gene signature for predicting breast cancer recurrence using transcriptome analysis. Future Oncol. 2021, 17, 71–80.
- Bernhardt, S.; Bayerlová, M.; Vetter, M.; Wachter, A.; Mitra, D.; Hanf, V.; Lantzsch, T.; Uleer, C.; Peschel, S.; John, J.; et al. Proteomic profiling of breast cancer metabolism identifies SHMT2 and ASCT2 as prognostic factors. Breast Cancer Res. 2017, 19, 112.
- Asiago, V.M.; Alvarado, L.Z.; Shanaiah, N.; Gowda, G.A.N.; Owusu-Sarfo, K.; Ballas, R.A.; Raftery, D. Early detection of recurrent breast cancer using metabolite profiling. Cancer Res. 2010, 70, 8309–8318.
- Oakman, C.; Tenori, L.; Claudino, W.M.; Cappadona, S.; Nepi, S.; Battaglia, A.; Bernini, P.; Zafarana, E.; Saccenti, E.; Fornier, M.; et al. Identification of a serum-detectable metabolomic fingerprint potentially correlated with the presence of micrometastatic disease in early breast cancer patients at varying risks of disease relapse by traditional prognostic methods. Ann. Oncol. 2011, 22, 1295–1301.
- Jobard, E.; Pontoizeau, C.; Blaise, B.J.; Bachelot, T.; Elena-Herrmann, B.; Trédan, O. A serum nuclear magnetic resonance-based metabolomic signature of advanced metastatic human breast cancer. Cancer Lett. 2014, 343, 33–41.
- Putluri, N.; Maity, S.; Kommagani, R.; Creighton, C.J.; Putluri, V.; Chen, F.; Nanda, S.; Bhowmik, S.K.; Terunuma, A.; Dorsey, T.; et al. Pathway-centric integrative analysis identifies RRM2 as a prognostic marker in breast cancer associated with poor survival and tamoxifen resistance. Neoplasia 2014, 16, 390–402.
- Luo, X.; Yu, H.; Song, Y.; Sun, T. Integration of metabolomic and transcriptomic data reveals metabolic pathway alteration in breast cancer and impact of related signature on survival. J. Cell. Physiol. 2019, 234, 13021–13031.
- Iqbal, M.A.; Siddiqui, S.; Ur Rehman, A.; Siddiqui, F.A.; Singh, P.; Kumar, B.; Saluja, D. Multiomics integrative analysis reveals antagonistic roles of CBX2 and CBX7 in metabolic reprogramming of breast cancer. Mol. Oncol. 2021, 15, 1450–1465.
- Camarda, R.; Zhou, A.Y.; Kohnz, R.A.; Balakrishnan, S.; Mahieu, C.; Anderton, B.; Eyob, H.; Kajimura, S.; Tward, A.; Krings, G.; et al. Inhibition of fatty acid oxidation as a therapy for MYC-overexpressing triple-negative breast cancer. Nat. Med. 2016, 22, 427–432.
- Kang, Y.P.; Yoon, J.-H.; Long, N.P.; Koo, G.-B.; Noh, H.-J.; Oh, S.-J.; Lee, S.B.; Kim, H.M.; Hong, J.Y.; Lee, W.J.; et al. Spheroid-induced epithelial-mesenchymal transition provokes global alterations of breast cancer lipidome: A multi-layered omics analysis. Front. Oncol. 2019, 9, 145.
- Terunuma, A.; Putluri, N.; Mishra, P.; Mathé, E.A.; Dorsey, T.H.; Yi, M.; Wallace, T.A.; Issaq, H.J.; Zhou, M.; Killian, J.K.; et al. MYC-driven accumulation of 2-hydroxyglutarate is associated with breast cancer prognosis. J. Clin. Investig. 2014, 124, 398–412.
- Budczies, J.; Brockmöller, S.F.; Müller, B.M.; Barupal, D.K.; Richter-Ehrenstein, C.; Kleine-Tebbe, A.; Griffin, J.L.; Orešič, M.; Dietel, M.; Denkert, C.; et al. Comparative metabolomics of estrogen receptor positive and estrogen receptor negative breast cancer: Alterations in glutamine and beta-alanine metabolism. J. Proteom. 2013, 94, 279–288.
- Lv, Y.; Wang, X.; Li, X.; Xu, G.; Bai, Y.; Wu, J.; Piao, Y.; Shi, Y.; Xiang, R.; Wang, L. Nucleotide de novo synthesis increases breast cancer stemness and metastasis via CGMP-PKG-MAPK signaling pathway. PLoS Biol. 2020, 18, e3000872.
- Stagg, J.; Divisekera, U.; McLaughlin, N.; Sharkey, J.; Pommey, S.; Denoyer, D.; Dwyer, K.M.; Smyth, M.J. Anti-CD73 antibody therapy inhibits breast tumor growth and metastasis. Proc. Natl. Acad. Sci. USA 2010, 107, 1547–1552.
- Aghaei, M.; Karami-Tehrani, F.; Salami, S.; Atri, M. Adenosine deaminase activity in the serum and malignant tumors of breast cancer: The assessment of isoenzyme ADA1 and ADA2 activities. Clinical Biochemistry 2005, 38, 887–891.
- Mahajan, M.; Tiwari, N.; Sharma, R.; Kaur, S.; Singh, N. Oxidative stress and its relationship with adenosine deaminase activity in various stages of breast cancer. Ind. J. Clin. Biochem. 2013, 28, 51–54.
- Sigoillot, F.D.; Sigoillot, S.M.; Guy, H.I. Breakdown of the regulatory control of pyrimidine biosynthesis in human breast cancer cells. Int. J. Cancer 2004, 109, 491–498.
- Serrano-Carbajal, E.A.; Espinal-Enríquez, J.; Hernández-Lemus, E. Targeting metabolic deregulation landscapes in breast cancer subtypes. Front. Oncol. 2020, 10, 97.
- Mejía-Pedroza, R.A.; Espinal-Enríquez, J.; Hernández-Lemus, E. Pathway-based drug repositioning for breast cancer molecular subtypes. Front. Pharmacol. 2018, 9, 905.
- Davison, C.; Morelli, R.; Knowlson, C.; McKechnie, M.; Carson, R.; Stachtea, X.; McLaughlin, K.A.; Prise, V.E.; Savage, K.; Wilson, R.H.; et al. Targeting nucleotide metabolism enhances the efficacy of anthracyclines and anti-metabolites in triple-negative breast cancer. NPJ Breast Cancer 2021, 7, 38.
- Furuta, E.; Okuda, H.; Kobayashi, A.; Watabe, K. Metabolic genes in cancer: Their roles in tumor progression and clinical implications. Biochim. Biophys. Acta 2010, 1805, 141–152.
- Zhang, H.; Liu, X.; Warden, C.D.; Huang, Y.; Loera, S.; Xue, L.; Zhang, S.; Chu, P.; Zheng, S.; Yen, Y. Prognostic and therapeutic significance of ribonucleotide reductase small subunit M2 in estrogen-negative breast cancers. BMC Cancer 2014, 14, 664.
- Chen, W.-X.; Yang, L.-G.; Xu, L.-Y.; Cheng, L.; Qian, Q.; Sun, L.; Zhu, Y.-L. bioinformatics analysis revealing prognostic significance of RRM2 gene in breast cancer. Biosci. Rep. 2019, 39.
- Mercier, I.; Casimiro, M.C.; Wang, C.; Rosenberg, A.L.; Quong, J.; Minkeu, A.; Allen, K.G.; Danilo, C.; Sotgia, F.; Bonuccelli, G.; et al. Human breast cancer-associated fibroblasts (CAFs) show caveolin-1 downregulation and RB tumor suppressor functional inactivation: Implications for the response to hormonal therapy. Cancer Biol. Ther. 2008, 7, 1212–1225.
- Kawano, Y.; Inada, Y.; Sasano, T.; Nishimura, N.; Hata, H.; Matsuoka, M. The purine metabolic enzyme AMPD1 is a novel therapeutic target for multiple myeloma. Blood 2018, 132, 5614.
- Wang, N.; Li, Y.; Zhou, J. Downregulation of ribonucleotide reductase subunits M2 induces apoptosis and G1 arrest of cervical cancer cells. Oncol. Lett. 2018, 15, 3719–3725.
- Mazzu, Y.Z.; Armenia, J.; Chakraborty, G.; Yoshikawa, Y.; Coggins, S.A.; Nandakumar, S.; Gerke, T.A.; Pomerantz, M.M.; Qiu, X.; Zhao, H.; et al. A novel mechanism driving poor-prognosis prostate cancer: Overexpression of the DNA repair gene, ribonucleotide reductase small subunit M2 (RRM2). Clin. Cancer Res. 2019, 25, 4480–4492.
- Zhuang, S.; Li, L.; Zang, Y.; Li, G.; Wang, F. RRM2 elicits the metastatic potential of breast cancer cells by regulating cell invasion, migration and VEGF expression via the PI3K/AKT signaling. Oncol. Lett. 2020, 19, 3349–3355.
- Pallante, P.; Terracciano, L.; Carafa, V.; Schneider, S.; Zlobec, I.; Lugli, A.; Bianco, M.; Ferraro, A.; Sacchetti, S.; Troncone, G.; et al. The loss of the CBX7 gene expression represents an adverse prognostic marker for survival of colon carcinoma patients. Eur. J. Cancer 2010, 46, 2304–2313.
- Karamitopoulou, E.; Pallante, P.; Zlobec, I.; Tornillo, L.; Carafa, V.; Schaffner, T.; Borner, M.; Diamantis, I.; Esposito, F.; Brunner, T.; et al. Loss of the CBX7 protein expression correlates with a more aggressive phenotype in pancreatic cancer. Eur. J. Cancer 2010, 46, 1438–1444.
- Clermont, P.-L.; Sun, L.; Crea, F.; Thu, K.L.; Zhang, A.; Parolia, A.; Lam, W.L.; Helgason, C.D. Genotranscriptomic meta-analysis of the polycomb gene CBX2 in human cancers: Initial evidence of an oncogenic role. Br. J. Cancer 2014, 111, 1663–1672.
- Zheng, S.; Lv, P.; Su, J.; Miao, K.; Xu, H.; Li, M. Overexpression of CBX2 in breast cancer promotes tumor progression through the PI3K/AKT signaling pathway. Am. J. Transl. Res. 2019, 11, 1668–1682.
- Klintman, M.; Buus, R.; Cheang, M.C.U.; Sheri, A.; Smith, I.E.; Dowsett, M. Changes in expression of genes representing key biologic processes after neoadjuvant chemotherapy in breast cancer, and prognostic implications in residual disease. Clin. Cancer Res. 2016, 22, 2405–2416.
- Svensson, R.U.; Parker, S.J.; Eichner, L.J.; Kolar, M.J.; Wallace, M.; Brun, S.N.; Lombardo, P.S.; Van Nostrand, J.L.; Hutchins, A.; Vera, L.; et al. Inhibition of acetyl-CoA carboxylase suppresses fatty acid synthesis and tumor growth of non-small-cell lung cancer in preclinical models. Nat. Med. 2016, 22, 1108–1119.
- Jones, J.E.C.; Esler, W.P.; Patel, R.; Lanba, A.; Vera, N.B.; Pfefferkorn, J.A.; Vernochet, C. Inhibition of acetyl-CoA carboxylase 1 (ACC1) and 2 (ACC2) reduces proliferation and de novo lipogenesis of EGFRvIII human glioblastoma cells. PLoS ONE 2017, 12, e0169566.
- Bai, J.; Zhang, X.; Kang, X.; Jin, L.; Wang, P.; Wang, Z. Screening of core genes and pathways in breast cancer development via comprehensive analysis of multi gene expression datasets. Oncol. Lett. 2019, 18, 5821–5830.
- Jeong, D.; Ham, J.; Kim, H.W.; Kim, H.; Ji, H.W.; Yun, S.H.; Park, J.E.; Lee, K.S.; Jo, H.; Han, J.H.; et al. ELOVL2: A novel tumor suppressor attenuating tamoxifen resistance in breast cancer. Am. J. Cancer Res. 2021, 11, 2568–2589.
- Tang, X.; Lin, C.-C.; Spasojevic, I.; Iversen, E.S.; Chi, J.-T.; Marks, J.R. A joint analysis of metabolomics and genetics of breast cancer. Breast Cancer Res. 2014, 16, 415.
- Dang, L.; White, D.W.; Gross, S.; Bennett, B.D.; Bittinger, M.A.; Driggers, E.M.; Fantin, V.R.; Jang, H.G.; Jin, S.; Keenan, M.C.; et al. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 2009, 462, 739–744.
- Figueroa, M.E.; Abdel-Wahab, O.; Lu, C.; Ward, P.S.; Patel, J.; Shih, A.; Li, Y.; Bhagwat, N.; Vasanthakumar, A.; Fernandez, H.F.; et al. Leukemic IDH1 and IDH2 mutations result in a hypermethylation phenotype, disrupt TET2 function, and impair hematopoietic differentiation. Cancer Cell 2010, 18, 553–567.
- Chen, X.; Cao, Q.; Liao, R.; Wu, X.; Xun, S.; Huang, J.; Dong, C. Loss of ABAT-mediated GABAergic system promotes basal-like breast cancer progression by activating Ca2+−NFAT1 axis. Theranostics 2019, 9, 34–47.
- Jansen, M.P.H.M.; Sas, L.; Sieuwerts, A.M.; Van Cauwenberghe, C.; Ramirez-Ardila, D.; Look, M.; Ruigrok-Ritstier, K.; Finetti, P.; Bertucci, F.; Timmermans, M.M.; et al. Decreased expression of ABAT and STC2 hallmarks ER-positive inflammatory breast cancer and endocrine therapy resistance in advanced disease. Mol. Oncol. 2015, 9, 1218–1233.
- Armiñán, A.; Palomino-Schätzlein, M.; Deladriere, C.; Arroyo-Crespo, J.J.; Vicente-Ruiz, S.; Vicent, M.J.; Pineda-Lucena, A. Metabolomics facilitates the discrimination of the specific anti-cancer effects of free- and polymer-conjugated doxorubicin in breast cancer models. Biomaterials 2018, 162, 144–153.
- Lord, S.R.; Collins, J.M.; Cheng, W.-C.; Haider, S.; Wigfield, S.; Gaude, E.; Fielding, B.A.; Pinnick, K.E.; Harjes, U.; Segaran, A.; et al. Transcriptomic analysis of human primary breast cancer identifies fatty acid oxidation as a target for metformin. Br. J. Cancer 2020, 122, 258–265.
- Tenori, L.; Oakman, C.; Claudino, W.M.; Bernini, P.; Cappadona, S.; Nepi, S.; Biganzoli, L.; Arbushites, M.C.; Luchinat, C.; Bertini, I.; et al. Exploration of serum metabolomic profiles and outcomes in women with metastatic breast cancer: A pilot study. Mol. Oncol. 2012, 6, 437–444.
- Cao, M.D.; Giskeødegård, G.F.; Bathen, T.F.; Sitter, B.; Bofin, A.; Lønning, P.E.; Lundgren, S.; Gribbestad, I.S. Prognostic value of metabolic response in breast cancer patients receiving neoadjuvant chemotherapy. BMC Cancer 2012, 12, 39.
- Wei, S.; Liu, L.; Zhang, J.; Bowers, J.; Gowda, G.A.N.; Seeger, H.; Fehm, T.; Neubauer, H.J.; Vogel, U.; Clare, S.E.; et al. Metabolomics approach for predicting response to neoadjuvant chemotherapy for breast cancer. Mol. Oncol. 2013, 7, 297–307.
- Choi, J.S.; Baek, H.-M.; Kim, S.; Kim, M.J.; Youk, J.H.; Moon, H.J.; Kim, E.-K.; Nam, Y.K. Magnetic resonance metabolic profiling of breast cancer tissue obtained with core needle biopsy for predicting pathologic response to neoadjuvant chemotherapy. PLoS ONE 2013, 8, e83866.
- Miolo, G.; Muraro, E.; Caruso, D.; Crivellari, D.; Ash, A.; Scalone, S.; Lombardi, D.; Rizzolio, F.; Giordano, A.; Corona, G. Pharmacometabolomics study identifies circulating spermidine and tryptophan as potential biomarkers associated with the complete pathological response to trastuzumab-paclitaxel neoadjuvant therapy in HER-2 positive breast cancer. Oncotarget 2016, 7, 39809–39822.
- Yoneten, K.K.; Kasap, M.; Akpinar, G.; Gunes, A.; Gurel, B.; Utkan, N.Z. Comparative proteome analysis of breast cancer tissues highlights the importance of glycerol-3-phosphate dehydrogenase 1 and monoacylglycerol lipase in breast cancer metabolism. Cancer Genom. Proteom. 2019, 16, 377–397.
- Shenoy, A.; Belugali Nataraj, N.; Perry, G.; Loayza Puch, F.; Nagel, R.; Marin, I.; Balint, N.; Bossel, N.; Pavlovsky, A.; Barshack, I.; et al. Proteomic patterns associated with response to breast cancer neoadjuvant treatment. Mol. Syst. Biol. 2020, 16.
- Arroyo-Crespo, J.J.; Armiñán, A.; Charbonnier, D.; Deladriere, C.; Palomino-Schätzlein, M.; Lamas-Domingo, R.; Forteza, J.; Pineda-Lucena, A.; Vicent, M.J. Characterization of triple-negative breast cancer preclinical models provides functional evidence of metastatic progression. Int. J. Cancer 2019, 145, 2267–2281.
- Yamashita, S.; Hattori, N.; Fujii, S.; Yamaguchi, T.; Takahashi, M.; Hozumi, Y.; Kogawa, T.; El-Omar, O.; Liu, Y.-Y.; Arai, N.; et al. Multi-omics analyses identify HSD17B4 methylation-silencing as a predictive and response marker of HER2-positive breast cancer to HER2-directed therapy. Sci. Rep. 2020, 10, 15530.
- Colombié, M.; Jézéquel, P.; Rubeaux, M.; Frenel, J.-S.; Bigot, F.; Seegers, V.; Campone, M. The EPICURE study: A pilot prospective cohort study of heterogeneous and massive data integration in metastatic breast cancer patients. BMC Cancer 2021, 21, 333.
- Cava, C.; Sabetian, S.; Castiglioni, I. Patient-specific network for personalized breast cancer therapy with multi-omics data. Entropy 2021, 23, 225.
- Gong, Y.; Ji, P.; Yang, Y.-S.; Xie, S.; Yu, T.-J.; Xiao, Y.; Jin, M.-L.; Ma, D.; Guo, L.-W.; Pei, Y.-C.; et al. Metabolic-pathway-based subtyping of triple-negative breast cancer reveals potential therapeutic targets. Cell Metab. 2021, 33, 51–64.e9.
- Mahendralingam, M.J.; Kim, H.; McCloskey, C.W.; Aliar, K.; Casey, A.E.; Tharmapalan, P.; Pellacani, D.; Ignatchenko, V.; Garcia-Valero, M.; Palomero, L.; et al. Mammary epithelial cells have lineage-rooted metabolic identities. Nat. Metab. 2021, 3, 665–681.
- Hilvo, M.; Denkert, C.; Lehtinen, L.; Müller, B.; Brockmöller, S.; Seppänen-Laakso, T.; Budczies, J.; Bucher, E.; Yetukuri, L.; Castillo, S.; et al. Novel theranostic opportunities offered by characterization of altered membrane lipid metabolism in breast cancer progression. Cancer Res. 2011, 71, 3236–3245.
- Kim, H.; Park, J.; Won, H.; Lee, J.; Kong, G. CBX7 Inhibits breast tumorigenicity through DKK-1-mediated suppression of the Wnt/β-catenin pathway. FASEB J. 2015, 29, 300–313.
- Piqué, D.G.; Montagna, C.; Greally, J.M.; Mar, J.C. A novel approach to modelling transcriptional heterogeneity identifies the oncogene candidate CBX2 in invasive breast carcinoma. Br. J. Cancer 2019, 120, 746–753.
- Clermont, P.-L.; Crea, F.; Chiang, Y.T.; Lin, D.; Zhang, A.; Wang, J.Z.L.; Parolia, A.; Wu, R.; Xue, H.; Wang, Y.; et al. Identification of the epigenetic reader CBX2 as a potential drug target in advanced prostate cancer. Clin. Epigenetics 2016, 8, 16.
- Xiao, X.; Huang, X.; Ye, F.; Chen, B.; Song, C.; Wen, J.; Zhang, Z.; Zheng, G.; Tang, H.; Xie, X. The miR-34a-LDHA axis regulates glucose metabolism and tumor growth in breast cancer. Sci. Rep. 2016, 6, 21735.
- Li, L.; Kang, L.; Zhao, W.; Feng, Y.; Liu, W.; Wang, T.; Mai, H.; Huang, J.; Chen, S.; Liang, Y.; et al. MiR-30a-5p suppresses breast tumor growth and metastasis through inhibition of LDHA-mediated warburg effect. Cancer Lett. 2017, 400, 89–98.
- Wang, Z.-Y.; Loo, T.Y.; Shen, J.-G.; Wang, N.; Wang, D.-M.; Yang, D.-P.; Mo, S.-L.; Guan, X.-Y.; Chen, J.-P. LDH-A silencing suppresses breast cancer tumorigenicity through induction of oxidative stress mediated mitochondrial pathway apoptosis. Breast Cancer Res. Treat. 2012, 131, 791–800.
- Marini, C.; Salani, B.; Massollo, M.; Amaro, A.; Esposito, A.I.; Orengo, A.M.; Capitanio, S.; Emionite, L.; Riondato, M.; Bottoni, G.; et al. Direct inhibition of hexokinase activity by metformin at least partially impairs glucose metabolism and tumor growth in experimental breast cancer. Cell Cycle 2013, 12, 3490–3499.
- Al-Ziaydi, A.G.; Al-Shammari, A.M.; Hamzah, M.I.; Kadhim, H.S.; Jabir, M.S. Hexokinase inhibition using D-mannoheptulose enhances oncolytic newcastle disease virus-mediated killing of breast cancer cells. Cancer Cell Int. 2020, 20, 420.
- Shah, K.N.; Mehta, K.R.; Peterson, D.; Evangelista, M.; Livesey, J.C.; Faridi, J.S. AKT-induced tamoxifen resistance is overturned by RRM2 inhibition. Mol. Cancer Res. 2014, 12, 394–407.
- Wong, C.C.; Qian, Y.; Yu, J. Interplay between epigenetics and metabolism in oncogenesis: Mechanisms and therapeutic approaches. Oncogene 2017, 36, 3359–3374.
- Mazor, T.; Chesnelong, C.; Pankov, A.; Jalbert, L.E.; Hong, C.; Hayes, J.; Smirnov, I.V.; Marshall, R.; Souza, C.F.; Shen, Y.; et al. Clonal expansion and epigenetic reprogramming following deletion or amplification of mutant IDH1. Proc. Natl. Acad. Sci. USA 2017, 114, 10743–10748.
- Park, J.-W.; Turcan, Ş. Epigenetic reprogramming for targeting IDH-mutant malignant gliomas. Cancers 2019, 11, 1616.
- Struys, E.A.; Verhoeven, N.M.; Ten Brink, H.J.; Wickenhagen, W.V.; Gibson, K.M.; Jakobs, C. Kinetic characterization of human hydroxyacid-oxoacid transhydrogenase: Relevance to D-2-hydroxyglutaric and γ-hydroxybutyric acidurias. J. Inherit. Metab. Dis. 2005, 28, 921–930.
- Mishra, P.; Tang, W.; Putluri, V.; Dorsey, T.H.; Jin, F.; Wang, F.; Zhu, D.; Amable, L.; Deng, T.; Zhang, S.; et al. ADHFE1 is a breast cancer oncogene and induces metabolic reprogramming. J. Clin. Investig. 2018, 128, 323–340.
- Mishra, P.; Tang, W.; Ambs, S. ADHFE1 is a MYC-linked oncogene that induces metabolic reprogramming and cellular de-differentiation in breast cancer. Mol. Cell. Oncol. 2018, 5, e1432260.
- Chajès, V.; Cambot, M.; Moreau, K.; Lenoir, G.M.; Joulin, V. Acetyl-CoA carboxylase alpha is essential to breast cancer cell survival. Cancer Res. 2006, 66, 5287–5294.
- Mauvoisin, D.; Charfi, C.; Lounis, A.M.; Rassart, E.; Mounier, C. Decreasing stearoyl-CoA desaturase-1 expression inhibits β-catenin signaling in breast cancer cells. Cancer Sci. 2013, 104, 36–42.
- Bao, J.; Zhu, L.; Zhu, Q.; Su, J.; Liu, M.; Huang, W. SREBP-1 is an independent prognostic marker and promotes invasion and migration in breast cancer. Oncol. Lett. 2016, 12, 2409–2416.
- Nie, L.-Y.; Lu, Q.-T.; Li, W.-H.; Yang, N.; Dongol, S.; Zhang, X.; Jiang, J. Sterol regulatory element-binding protein 1 is required for ovarian tumor growth. Oncol. Rep. 2013, 30, 1346–1354.
- Wen, Y.-A.; Xiong, X.; Zaytseva, Y.Y.; Napier, D.L.; Vallee, E.; Li, A.T.; Wang, C.; Weiss, H.L.; Evers, B.M.; Gao, T. Downregulation of SREBP inhibits tumor growth and initiation by altering cellular metabolism in colon cancer. Cell Death Dis. 2018, 9, 265.
- Zhang, H.; Liu, S.; Cai, Z.; Dong, W.; Ye, J.; Cai, Z.; Han, Z.; Liang, Y.; Zhuo, Y.; Luo, Y.; et al. Down-regulation of ACACA suppresses the malignant progression of prostate cancer through inhibiting mitochondrial potential. J. Cancer 2021, 12, 232–243.
- Menendez, J.A.; Lupu, R. Fatty acid synthase (FASN) as a therapeutic target in breast cancer. Expert Opin. Ther. Targets 2017, 21, 1001–1016.
- Yokoyama, C.; Wang, X.; Briggs, M.R.; Admon, A.; Wu, J.; Hua, X.; Goldstein, J.L.; Brown, M.S. SREBP-1, a basic-helix-loop-helix-leucine zipper protein that controls transcription of the low density lipoprotein receptor gene. Cell 1993, 75, 187–197.
- Jeon, T.-I.; Osborne, T.F. SREBPs: Metabolic integrators in physiology and metabolism. Trends Endocrinol. Metab. 2012, 23, 65–72.
- Huang, W.-C.; Li, X.; Liu, J.; Lin, J.; Chung, L.W.K. Activation of androgen receptor, lipogenesis, and oxidative stress converged by SREBP-1 is responsible for regulating growth and progression of prostate cancer cells. Mol. Cancer Res. 2012, 10, 133–142.
- Miyachi, K.; Sawada, Y.; Shida, Y.; Sugawara, A.; Hisatomi, H. Lipogenic gene expression profile in patients with gastric cancer. Mol. Clin. Oncol. 2013, 1, 825–827.
- Tan, Z.; Xiao, L.; Tang, M.; Bai, F.; Li, J.; Li, L.; Shi, F.; Li, N.; Li, Y.; Du, Q.; et al. Targeting CPT1A-mediated fatty acid oxidation sensitizes nasopharyngeal carcinoma to radiation therapy. Theranostics 2018, 8, 2329–2347.
- Han, S.; Wei, R.; Zhang, X.; Jiang, N.; Fan, M.; Huang, J.H.; Xie, B.; Zhang, L.; Miao, W.; Butler, A.C.-P.; et al. CPT1A/2-mediated FAO enhancement—A metabolic target in radioresistant breast cancer. Front. Oncol. 2019, 9, 1201.
- Xiong, X.; Wen, Y.-A.; Fairchild, R.; Zaytseva, Y.Y.; Weiss, H.L.; Evers, B.M.; Gao, T. Upregulation of CPT1A is essential for the tumor-promoting effect of adipocytes in colon cancer. Cell Death Dis. 2020, 11, 736.
- Pucci, S.; Zonetti, M.J.; Fisco, T.; Polidoro, C.; Bocchinfuso, G.; Palleschi, A.; Novelli, G.; Spagnoli, L.G.; Mazzarelli, P. Carnitine palmitoyl transferase-1A (CPT1A): A new tumor specific target in human breast cancer. Oncotarget 2016, 7, 19982–19996.
- Schlaepfer, I.R.; Joshi, M. CPT1A-mediated fat oxidation, mechanisms, and therapeutic potential. Endocrinology 2020, 161.
More
Information
Subjects:
Pharmacology & Pharmacy
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
899
Revisions:
2 times
(View History)
Update Date:
24 Sep 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No