 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Raghad AL-Ishaq | + 4801 word(s) | 4801 | 2021-08-09 07:59:16 | | | |
| 2 | Vivi Li | + 237 word(s) | 5038 | 2021-08-09 11:28:08 | | |
Video Upload Options
Gastrointestinal (GI) cancer is a prevalent global health disease with a massive burden on health care providers. Internal and external factors such as obesity, smoking, diet (red meat), low socioeconomic status and infection with Helicobacter pylori are the critical risk factors of GI cancers. Flavonoids are natural phenolic compounds found abundantly in fruits and vegetables. Upon ingestion, 90% of flavonoids consumed require further enzymatic metabolism by the gut microbiome to enhance their bioavailability and absorption. Several epidemiological studies reported that consumption of flavonoids and their enzymatic conversion by gut microbes is strongly associated with the reduced risk of GI cancer development. This review summarizes the current knowledge on the enzymatic conversion of flavonoids by the human gut microbiome. It also addresses the underlying anti-GI cancer effects on metabolic pathways such as apoptosis and cellular proliferation. Overall, metabolites produced from flavonoid’s enzymatic conversion illustrate anti-GI cancer effects, but the mechanisms of action need further clarification.
1. Introduction
1.1. Gastrointestinal Cancer
1.2. Flavonoids and Cancer
1.3. Flavonoids Metabolism in the Gut
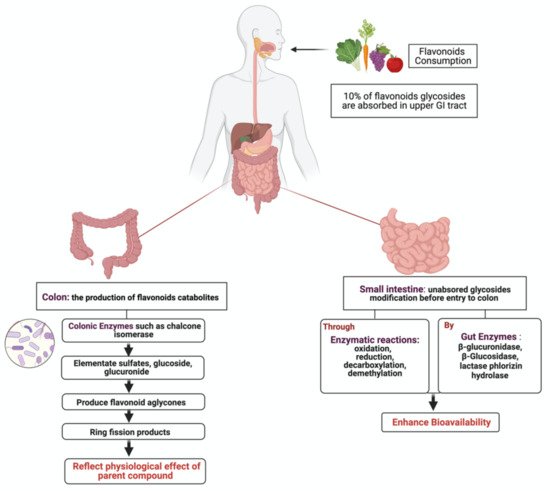
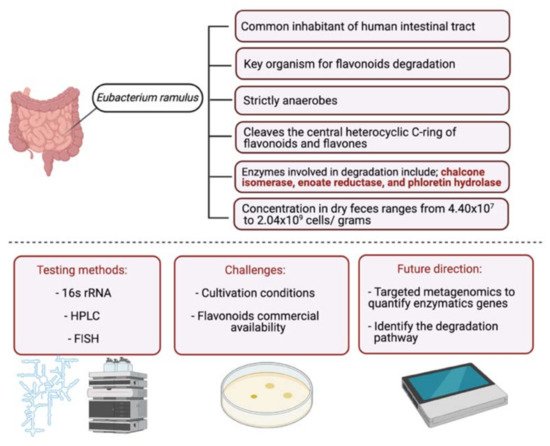
2. Metabolism of Flavonoids by Gut Microbiota
| Flavonoid Subclass | Name of Flavonoid | Structure of Flavonoid | Dietary Source | Metabolites Produced by Gut Microbiota | Bacteria Involved in Metabolism | Enzymes Involved in Metabolism | Site of Metabolism | Conversion Mechanism | Effect of Microbiota on Flavonoids | Model Used | References | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| In Vivo | In Vitro | ||||||||||||
| Flavonol | 1. Rutin | 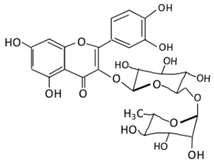 |
Lemons, berried, limes and oranges |
Quercetin -3- O- glucoside Quercetin |
Lachnoclostridium Eisenbergiella Escherichia Parabacteroides Erysipelatoclostridium |
α-rhamnosidases β-glucosidases |
Colon | Hydrolysis of rutin to remove sugar moiety | Permit the absorption of the aglycone Enhance bioavailability |
10 fecal samples from healthy individual following omnivore diet. |
[27][28][29][30][31] | ||
| 2. Fisetin | 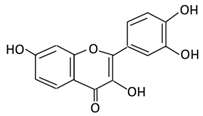 |
Persimmon and onions | No available data | [32][33] | |||||||||
| 3.Kaempferol | 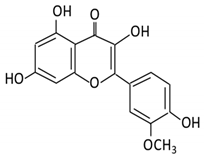 |
Tea, berries and cruciferous vegetables |
Kaempferol -3-O- glucoside p-coumaric acid kaempferol 3-(4 hydroxyphenyl) propionic acid 3-phenylpropionic acid |
Lactobacillus paracasei A221 |
β-glucosidases | Intestinal tract | (A)Degradation through multiple chemical reactions: - Deglycosylation - Reduction - Dehydroxylation (B) Hydrolysis |
Permit the absorption of the aglycone Enhance bioavailability |
Mice | 3 fecal samples from healthy individual Caco-2 intestinal barrier model |
[34][35][36][37] | ||
| 4. Quercetin | 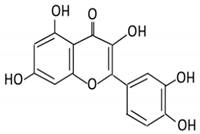 |
Black currants, cherries, apples and chokeberries |
4-hydroxybenzoic acid 3,4-dihydroxyphenylacetic acid 3,4-dihydroxybenzoic 3-(3-hydroxyphenyl) propionic acid |
Bacteroides fragilis Clostridium perfringens Eubacterium ramulus Streptococcus S-2 Lactobacillus L-2 Bifidobacterium B-9 Bacteroides JY-6 |
Lactate phlorizin hydrolase | Small intestine |
Deglycosylation | Enhance bioavailability | Mice | 87 fecal samples from healthy elderly individual |
[38][39][40][41][42][43] | ||
| 5. Isorhamnetin | 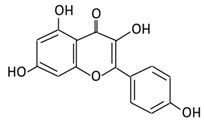 |
Ginkgo biloba and Hippophae rhamnoides |
3-O-neohesperidoside Isorhamnetin -3- glucoside Aglycone isorhamnetin |
Escherichia Enterococcus Bacillus. |
Not identified | Small intestine |
Deglycosylation | Permit the absorption of the aglycone | Rats | 1 fecal sample from healthy female |
[44][45] | ||
| 6. Morin | 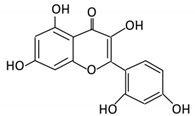 |
Psidium guajava, Prunus dulcis (Almond), chlorophora tinctoria and fruits |
Morin glucuronides Morin sulfates |
No available data | [12] | ||||||||
| Flavanones | 7. Hesperidin | 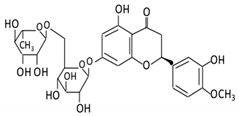 |
Orange citrus aurantium | Hesperetin | Not identified | Rutinose | Large intestine |
Cleaves the attached rutinose moiety |
Permit the absorption of the aglycone Enhance bioavailability |
fecal/ blood samples from 18 Lewis male rats |
[46][47][48][49] | ||
| 8. Naringenin | 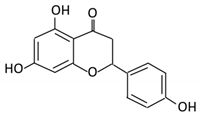 |
C.aurantium (chinese herbs) and grapefruit | Aglycone naringenin | Ruminococcus gauvreauii Bifidobacterium catenulatum Enterococcus caccae Eubacterium ramulus |
Chalcone isomerase | Large intestine |
Remove sugar group | Permit the absorption of the aglycone Enhance bioavailability |
fecal samples from healthy individuals |
[50][51][52] | |||
| 9. Eriodictyol | 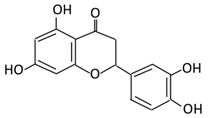 |
Lemon (Eriocitrin), Torr, Eridictyon californicum, | Eriodictyol 3,4-dihydroxyhydrocinnamic acid Phloroglucinol |
Parabacteroides distasonis Bacteroides uniformis JCM 5828 |
Chalcone isomerase | Colon | O-Deglycosylation | Permit the absorption of the aglycone Enhance bioavailability |
fecal samples from healthy individuals | [53][54][55] | |||
| Isoflavones | 10. Genistein | 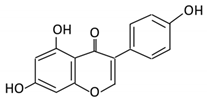 |
Fava and soy beans | Dihydrogenistein 6-hydroxy-O-desmethylangolensin 2-(4-hydroxyphenyl) propionic acid |
Lactobacillus Eubacterium ramulus |
Lactate phlorizin hydrolase | Small intestine |
Fermentation by anerobic bacteria | Permit the absorption of the aglycone Enhance bioavailability |
Samples from C57BL/6 female mouse |
[56][57][58][59] | ||
| 11. Daidzein | 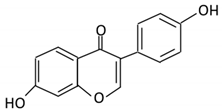 |
Soybeans, nuts and soymilk | Dihydrodaidzein O-desmethylangolensin S- equol |
Clostridium-like strain | β- glucosidase Lactate phlorizin hydrolase |
Colon | Reduction | Permit the absorption of the aglycone Enhance bioavailability |
Mice | [60][61][62] | |||
| Anthocyanins | 12. Cyanidin | 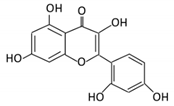 |
Bilberry, blueberry, grapes, blackberries, hawthorn | Cyanidin-3- glucoside | Clostridium saccharogumia Eubacterium ramulus |
Combined activity of bacteria and host enzymes | Small intestine |
Degrade polyphenolic glycosides |
Permit the absorption of the aglycone Enhance bioavailability |
Dawley rats | Fecal samples from healthy individuals | [63][64][65] | |
| 13. Delphinidin | 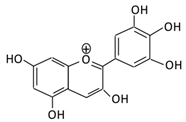 |
Dark grapes, eggplant, berries, red cabbage, | Gallic acids | Lactobacillus | β-d-glucuronidase β-d-glucosidase α-rhamnosidase α-galactosidase |
Colon | Cleavage of glycosidic bonds | Permit the absorption of the aglycone Enhance bioavailability |
Mice | [66][67][68][69][70] | |||
| 14. Pelargonidin | 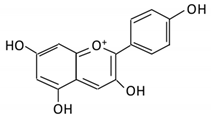 |
Bilberry and ficus bengalensis Linn | 4-hydroxybenzoic | Lactobacillus | β-d-glucosidase β-d-glucuronidase α-galactosidase α-rhamnosidase |
Colon | Cleavage of glycosidic bonds | Permit the absorption of the aglycone Enhance bioavailability |
Mice | [71][72] | |||
| Flavones | 15. Baicalein | 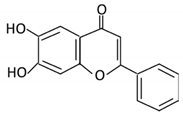 |
Scutellaria lateriflora L | Baicalein | E. coli | β-glucuronidase | Intestine | Hydrolysis to remove moiety |
Permit the absorption of the aglycone Enhance bioavailability |
Mice | HCT-166 cells SW-480 cells |
[73][74][75][76] | |
| 16. Luteolin | 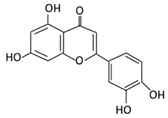 |
broccoli, celery and parsley, | No available data | [12] | |||||||||
| 17. Diosmin | 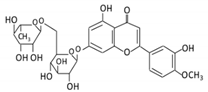 |
Citrus fruits | Diosmetin | Not identified | α- glucosidase β- glucosidase |
Small intestine |
Hydrolysis | Enhance bioavailability | Blood samples from healthy participants |
[77][78][79][80] | |||
| 18. Apigenin | 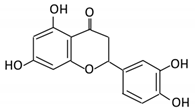 |
Tea, chamomile, parsley and oranges |
3-(4-hydroxyphenyl) propionic acid Apigenin |
Bacteroides distasonis Eubacterium ramulus Clostridium orbiscindens |
β- glucosidase lactase-phlorizin hydrolase |
Small intestine |
Glucuronidation Hydrolysis |
Enhance bioavailability |
Rats | Fecal and urine samples | [81][82][83][84] | ||
| 19. Tangeretin | 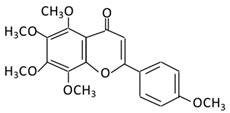 |
Poncirus trifoliate L, citrus fruit | Tangeretin-O-glucuronides | Lactobacillus Bifidobacterium |
Possibly by: A) rhamno glucosides B) C- glycosyl |
Small intestine |
Demethylation | Permit the absorption of the aglycone |
Rats | [85][86] | |||
| 20. Wogonin | 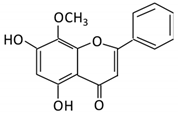 |
Scutellaria baicalensis Georgi | Wogonin | Not identified | β-glucuronidase | Intestine | Hydrolysis | Enhance absorption and bioavailability | Sprague-Dawley rats | [87][88][89][90] | |||
| 21. Chrysin | 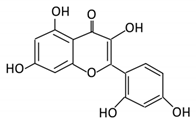 |
Honey | Chrysin glucuronides | Flavonifractor plautii ATCC 49531 | Flavone reductase |
Intestine | Reduction | catalyzes the hydrogenation of the C2–C3 double bond | ATCC 49,531 strain | [91][92][93] | |||
2.1. Flavonol
2.1.1. Rutin
2.1.2. Fisetin
2.1.3. Kaempferol
2.1.4. Quercetin
2.1.5. Isorhamnetin
2.2. Flavanones
2.2.1. Hesperidin
2.2.2. Naringenin
2.2.3. Eriodictyol
2.3. Isoflavones
2.3.1. Genistein
2.3.2. Daidzein
2.4. Anthocyanins
2.4.1. Cyanidin
2.4.2. Delphinidin
2.4.3. Pelargonidin
2.5. Flavones
2.5.1. Baicalein
2.5.2. Diosmin
2.5.3. Apigenin
2.5.4. Tangeretin
2.5.5. Wogonin
2.5.6. Chrysin
3. Effects of Flavonoids on Gastrointestinal Cancer
3.1. Flavonoids Effects on Impaired General Pathways
3.1.1. Apoptosis
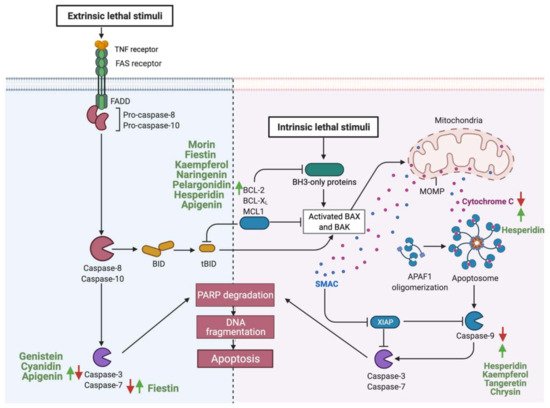
3.1.2. Cellular Proliferation
4. Discussion
4.1. Clinical Implementation of Flavonoids
4.2. Impact of Current Cancer Treatment on Gut Enzymes
4.3. Possible Synergistic Effects of Flavonoids?
References
- Hassanzade, J.; Molavi, E.V.H.; Farahmand, M.; Rajaiifard, A.R. Incidence and Mortality Rate of Common Gastrointestinal Cancers in South of Iran, a Population Based Study. Iran. J. Cancer Prev. 2011, 4, 163–169.
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Busselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2018, 11, 28.
- Rozen, P. Cancer of the gastrointestinal tract: Early detection or early prevention? Eur. J. Cancer Prev. 2004, 13, 71–75.
- Pourhoseingholi, M.A.; Vahedi, M.; Baghestani, A.R. Burden of gastrointestinal cancer in Asia; an overview. Gastroenterol. Hepatol. Bed Bench. 2015, 8, 19–27.
- Zali, H.; Rezaei-Tavirani, M.; Azodi, M. Gastric cancer: Prevention, risk factors and treatment. Gastroenterol. Hepatol. Bed Bench. 2011, 4, 175–185.
- Han, C.J.; Reding, K.; Cooper, B.A.; Paul, S.M.; Conley, Y.P.; Hammer, M.; Miaskowski, C. Symptom Clusters in Patients With Gastrointestinal Cancers Using Different Dimensions of the Symptom Experience. J. Pain Symptom Manag. 2019, 58, 224–234.
- Correa, P. Gastric cancer: Overview. Gastroenterol. Clin N. Am. 2020, 42, 211–217.
- Igney, F.H.; Krammer, P.H. Death and anti-death: Tumour resistance to apoptosis. Nat. Rev. Cancer 2002, 2, 277–288.
- Qiao, L.; Wong, B.C. Targeting apoptosis as an approach for gastrointestinal cancer therapy. Drug Resist. Updat. 2009, 12, 55–64.
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–15.
- Kozlowska, A.; Szostak-Wegierek, D. Flavonoids—food sources and health benefits. Rocz. Panstw. Zakl. Hig. 2014, 65, 79–85.
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Busselberg, D. Flavonoids and Their Anti-Diabetic Effects: Cellular Mechanisms and Effects to Improve Blood Sugar Levels. Biomolecules 2019, 9, 430.
- Romagnolo, D.F.; Selmin, O.I. Flavonoids and cancer prevention: A review of the evidence. J. Nutr. Gerontol. Geriatr. 2012, 31, 206–238.
- Jun, S.; Shin, S.; Joung, H. Estimation of dietary flavonoid intake and major food sources of Korean adults. Br. J. Nutr. 2016, 115, 480–489.
- Yu, Y.; Jing, X.; Li, H.; Zhao, X.; Wang, D. Soy isoflavone consumption and colorectal cancer risk: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 25939.
- Molina-Montes, E.; Sanchez, M.J.; Zamora-Ros, R.; Bueno-de-Mesquita, H.B.; Wark, P.A.; Obon-Santacana, M.; Duell, E.J. Flavonoid and lignan intake and pancreatic cancer risk in the European prospective investigation into cancer and nutrition cohort. Int. J. Cancer 2016, 139, 1480–1492.
- Xu, M.; Chen, Y.M.; Huang, J.; Fang, Y.J.; Huang, W.Q.; Yan, B.; Zhang, C.X. Flavonoid intake from vegetables and fruits is inversely associated with colorectal cancer risk: A case-control study in China. Br. J. Nutr. 2016, 116, 1275–1287.
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672.
- Chen, Z.; Zheng, S.; Li, L.; Jiang, H. Metabolism of flavonoids in human: A comprehensive review. Curr. Drug Metab. 2014, 15, 48–61.
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341.
- Pei, R.; Liu, X.; Bolling, B. Flavonoids and gut health. Curr. Opin. Biotechnol. 2020, 61, 153–159.
- Murota, K.; Nakamura, Y.; Uehara, M. Flavonoid metabolism: The interaction of metabolites and gut microbiota. Biosci. Biotechnol. Biochem. 2018, 82, 600–610.
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370.
- Del Rio, D.; Calani, L.; Scazzina, F.; Jechiu, L.; Cordero, C.; Brighenti, F. Bioavailability of catechins from ready-to-drink tea. Nutrition 2010, 26, 528–533.
- Braune, A.; Engst, W.; Elsinghorst, P.W.; Furtmann, N.; Bajorath, J.; Gutschow, M.; Blaut, M. Chalcone Isomerase from Eubacterium ramulus Catalyzes the Ring Contraction of Flavanonols. J. Bacteriol. 2016, 198, 2965–2974.
- Simmering, R.; Pforte, H.; Jacobasch, G.; Blaut, M. The growth of the flavonoid-degrading intestinal bacterium, Eubacterium ramulus, is stimulated by dietary flavonoids in vivo. FEMS Microbiol. Ecol. 2002, 40, 243–248.
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hemon, B.; Moskal, A.; Overvad, K.; Scalbert, A. Dietary polyphenol intake in Europe: The European Prospective Investigation into Cancer and Nutrition (EPIC) study. Eur. J. Nutr. 2016, 55, 1359–1375.
- Ghorbani, A. Mechanisms of antidiabetic effects of flavonoid rutin. Biomed Pharm. 2017, 96, 305–312.
- Shin, N.R.; Moon, J.S.; Shin, S.Y.; Li, L.; Lee, Y.B.; Kim, T.J.; Han, N.S. Isolation and characterization of human intestinal Enterococcus avium EFEL009 converting rutin to quercetin. Lett. Appl. Microbiol. 2016, 62, 68–74.
- Riva, A.; Kolimar, D.; Spittler, A.; Wisgrill, L.; Herbold, C.W.; Abranko, L.; Berry, D. Conversion of Rutin, a Prevalent Dietary Flavonol, by the Human Gut Microbiota. Front. Microbiol. 2020, 11, 585428.
- Mazzeo, M.F.; Lippolis, R.; Sorrentino, A.; Liberti, S.; Fragnito, F.; Siciliano, R.A. Lactobacillus acidophilus-Rutin Interplay Investigated by Proteomics. PLoS ONE 2015, 10, e0142376.
- Khan, N.; Syed, D.N.; Ahmad, N.; Mukhtar, H. Fisetin: A dietary antioxidant for health promotion. Antioxid. Redox Signal. 2013, 19, 151–162.
- Kawabata, K.; Sugiyama, Y.; Sakano, T.; Ohigashi, H. Flavonols enhanced production of anti-inflammatory substance(s) by Bifidobacterium adolescentis: Prebiotic actions of galangin, quercetin, and fisetin. Biofactors 2013, 39, 422–429.
- Imran, M.; Salehi, B.; Sharifi-Rad, J.; Aslam Gondal, T.; Saeed, F.; Imran, A.; Estevinho, L.M. Kaempferol: A Key Emphasis to Its Anticancer Potential. Molecules 2019, 24, 2277.
- Crespy, V.; Morand, C.; Besson, C.; Cotelle, N.; Vezin, H.; Demigne, C.; Remesy, C. The splanchnic metabolism of flavonoids highly differed according to the nature of the compound. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G980–G988.
- Vollmer, M.; Esders, S.; Farquharson, F.M.; Neugart, S.; Duncan, S.H.; Schreiner, M.; Rohn, S. Mutual Interaction of Phenolic Compounds and Microbiota: Metabolism of Complex Phenolic Apigenin-C- and Kaempferol-O-Derivatives by Human Fecal Samples. J. Agric. Food Chem. 2018, 66, 485–497.
- Shimojo, Y.; Ozawa, Y.; Toda, T.; Toda, T.; Igami, K.; Shimizu, T. Probiotic Lactobacillus paracasei A221 improves the functionality and bioavailability of kaempferol-glucoside in kale by its glucosidase activity. Sci. Rep. 2018, 8, 9239.
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Cho, W.C. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega. 2020, 5, 11849–11872.
- Guo, Y.; Bruno, R.S. Endogenous and exogenous mediators of quercetin bioavailability. J. Nutr. Biochem. 2015, 26, 201–210.
- Bischoff, S.C. Quercetin: Potentials in the prevention and therapy of disease. Curr. Opin. Clin Nutr. Metab. Care 2008, 11, 733–740.
- Najmanova, I.; Pourova, J.; Voprsalova, M.; Pilarova, V.; Semecky, V.; Novakova, L.; Mladenka, P. Flavonoid metabolite 3-(3-hydroxyphenyl)propionic acid formed by human microflora decreases arterial blood pressure in rats. Mol. Nutr. Food Res. 2016, 60, 981–991.
- Santangelo, R.; Silvestrini, A.; Mancuso, C. Ginsenosides, catechins, quercetin and gut microbiota: Current evidence of challenging interactions. Food Chem. Toxicol. 2019, 123, 42–49.
- Tamura, M.; Hoshi, C.; Kobori, M.; Takahashi, S.; Tomita, J.; Nishimura, M.; Nishihira, J. Quercetin metabolism by fecal microbiota from healthy elderly human subjects. PLoS ONE 2017, 12, e0188271.
- Vinayagam, R.; Xu, B. Antidiabetic properties of dietary flavonoids: Acellular mechanism review. Nutr. Metab. Lond. 2015, 12, 60.
- Du, L.Y.; Zhao, M.; Tao, J.H.; Qian, D.W.; Jiang, S.; Shang, E.X.; Duan, J.A. The Metabolic Profiling of Isorhamnetin-3-O-Neohesperidoside Produced by Human Intestinal Flora Employing UPLC-Q-TOF/MS. J. Chromatogr. Sci. 2017, 55, 243–250.
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and anti-inflammatory properties of the citrus flavonoids hesperidin and hesperetin: An updated review of their molecular mechanisms and experimental models. Phyther. Res. 2015, 29, 323–331.
- Williamson, G. The role of polyphenols in modern nutrition. Nutr. Bull. 2017, 42, 226–235.
- Garg, A.; Garg, S.; Zaneveld, L.J.; Singla, A.K. Chemistry and pharmacology of the citrus bioflavonoid hesperidin. Phyther. Res. 2001, 15, 655–669.
- Estruel-Amades, S.; Massot-Cladera, M.; Perez-Cano, F.J.; Franch, A.; Castell, M.; Camps-Bossacoma, M. Hesperidin Effects on Gut Microbiota and Gut-Associated Lymphoid Tissue in Healthy Rats. Nutrients 2019, 11, 324.
- Chen, T.; Wu, H.; He, Y.; Pan, W.; Yan, Z.; Liao, Y.; Yao, H. Simultaneously Quantitative Analysis of Naringin and Its Major Human Gut Microbial Metabolites Naringenin and 3-(4′-Hydroxyphenyl) Propanoic Acid via Stable Isotope Deuterium-Labeling Coupled with RRLC-MS/MS Method. Molecules 2019, 24, 4287.
- Feng, T.; Wang, K.; Liu, F.; Ye, R.; Zhu, X.; Zhuang, H.; Xu, Z. Structural characterization and bioavailability of ternary nanoparticles consisting of amylose, α β -linoleic acid and _-lactoglobulin complexed with naringin. Int. J. Biol. Macromol. 2017, 99, 365–374.
- Firrman, J.; Liu, L.; Argoty, G.A.; Zhang, L.; Tomasula, P.; Wang, M.; Xiao, W. Analysis of Temporal Changes in Growth and Gene Expression for Commensal Gut Microbes in Response to the Polyphenol Naringenin. Microbiol. Insights. 2018, 11.
- Islam, A.; Islam, M.S.; Rahman, M.K.; Uddin, M.N.; Akanda, M.R. The pharmacological and biological roles of eriodictyol. Arch. Pharm. Res. 2020, 43, 582–592.
- Braune, A.; Blaut, M. Bacterial species involved in the conversion of dietary flavonoids in the human gut. Gut. Microbes. 2016, 7, 216–234.
- Miyake, Y.; Yamamoto, K.; Osawa, T. Metabolism of Antioxidant in Lemon Fruit (Citrus limon BURM. f.) by Human Intestinal Bacteria. J. Agric. Food Chem. 1997, 45, 3738–3742.
- Li, H.Q.; Luo, Y.; Qiao, C.H. The mechanisms of anticancer agents by genistein and synthetic derivatives of isoflavone. Mini. Rev. Med. Chem. 2012, 12, 350–362.
- Matthies, A.; Clavel, T.; Gutschow, M.; Engst, W.; Haller, D.; Blaut, M.; Braune, A. Conversion of daidzein and genistein by an anaerobic bacterium newly isolated from the mouse intestine. Appl. Environ. Microbiol. 2008, 74, 4847–4852.
- Wang, X.L.; Shin, K.H.; Hur, H.G.; Kim, S.I. Enhanced biosynthesis of dihydrodaidzein and dihydrogenistein by a newly isolated bovine rumen anaerobic bacterium. J. Biotechnol. 2005, 115, 261–269.
- Heinonen, S.; Wahala, K.; Adlercreutz, H. Identification of isoflavone metabolites dihydrodaidzein, dihydrogenistein, 6′-OH-O-dma, and cis-4-OH-equol in human urine by gas chromatography-mass spectroscopy using authentic reference compounds. Anal. Biochem. 1999, 274, 211–219.
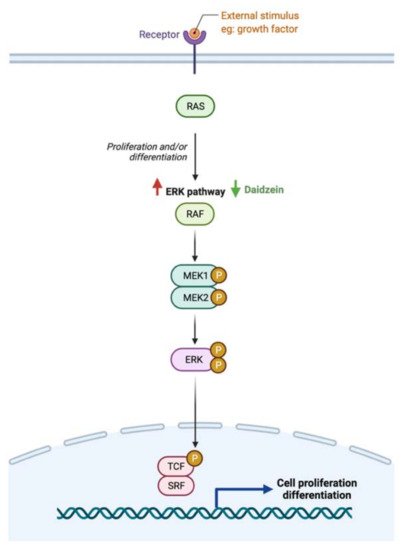
- Das, D.; Sarkar, S.; Bordoloi, J.; Wann, S.B.; Kalita, J.; Manna, P. Daidzein, its effects on impaired glucose and lipid metabolism and vascular inflammation associated with type 2 diabetes. Biofactors 2018, 44, 407–417.
- Maddalena, R.; Alberto, A.; Roncaglia, L.; Alan, L.; Stefano, R. Dietary isoflavones and intestinal microbiota. In Isoflavones Biosynthesis, Occurence and Health Effects; Thompson, J., Ed.; Nova Science Publishers: New York, NY, USA, 2010; pp. 137–161.
- Wang, X.L.; Kim, H.J.; Kang, S.I.; Kim, S.I.; Hur, H.G. Production of phytoestrogen S-equol from daidzein in mixed culture of two anaerobic bacteria. Arch. Microbiol. 2007, 187, 155–160.
- Akkarachiyasit, S.; Charoenlertkul, P.; Yibchok-Anun, S.; Adisakwattana, S. Inhibitory activities of cyanidin and its glycosides and synergistic effect with acarbose against intestinal alpha-glucosidase and pancreatic alpha-amylase. Int. J. Mol. Sci. 2010, 11, 3387–3396.
- McGhie, T.K.; Walton, M.C. The bioavailability and absorption of anthocyanins: Towards a better understanding. Mol. Nutr. Food Res. 2007, 51, 702–713.
- Hanske, L.; Engst, W.; Loh, G.; Sczesny, S.; Blaut, M.; Braune, A. Contribution of gut bacteria to the metabolism of cyanidin 3-glucoside in human microbiota-associated rats. Br. J. Nutr. 2013, 109, 1433–1441.
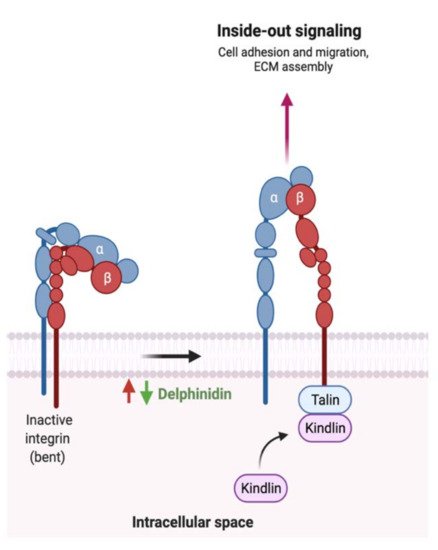
- Ko, H.; Jeong, M.H.; Jeon, H.; Sung, G.J.; So, Y.; Kim, I.; Choi, K.C. Delphinidin sensitizes prostate cancer cells to TRAIL-induced apoptosis, by inducing DR5 and causing caspase-mediated HDAC3 cleavage. Oncotarget 2015, 6, 9970–9984.
- Lim, W.; Song, G. Inhibitory effects of delphinidin on the proliferation of ovarian cancer cells via PI3K/AKT and ERK 1/2 MAPK signal transduction. Oncol. Lett. 2017, 14, 810–818.
- Eker, M.E.; Aaby, K.; Budic-Leto, I.; Brncic, S.R.; El, S.N.; Karakaya, S.; Pascual-Teresa, S. A Review of Factors Affecting Anthocyanin Bioavailability: Possible Implications for the Inter-Individual Variability. Foods 2019, 9, 2.
- Aura, A.M.; Martin-Lopez, P.; O’Leary, K.A.; Williamson, G.; Oksman-Caldentey, K.M.; Poutanen, K.; Santos-Buelga, C. In vitro metabolism of anthocyanins by human gut microflora. Eur. J. Nutr. 2005, 44, 133–142.
- Flores, G.; Costabile, A.; Klee, A.; Guergoletto, K.; Gibson, G. In vitro fermentation of anthocyanins encapsulated with cyclodextrins: Release, metabolism and influence on gut microbiota growth. J. Funct. Foods 2015, 16, 10.
- Mazza, G. Compositional and Functional Properties of Saskatoon Berry and Blueberry. Int. J. Fruit Sci. 2006, 5, 101–120.
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A Comprehensive Review of Their Chemical Properties and Health Effects on Cardiovascular and Neurodegenerative Diseases. Molecules 2020, 25, 3809.
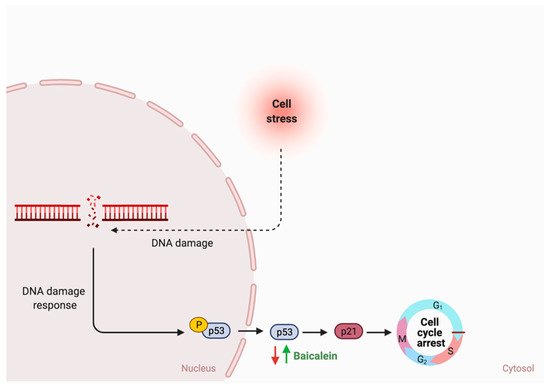
- Sowndhararajan, K.; Deepa, P.; Kim, M.; Park, S.J.; Kim, S. Baicalein as a potent neuroprotective agent: A review. Biomed. Pharm. 2017, 95, 1021–1032.
- Li-Weber, M. New therapeutic aspects of flavones: The anticancer properties of Scutellaria and its main active constituents wogonin, baicalein and baicalin. Cancer Treat. Rev. 2009, 35, 57–68.
- Noh, K.; Kang, Y.; Nepal, M.R.; Jeong, K.S.; Oh, D.G.; Kang, M.J.; Jeong, T.C. Role of Intestinal Microbiota in Baicalin-Induced Drug Interaction and Its Pharmacokinetics. Molecules 2016, 21, 337.
- Wang, C.Z.; Zhang, C.F.; Chen, L.; Anderson, S.; Lu, F.; Yuan, C.S. Colon cancer chemopreventive effects of baicalein, an active enteric microbiome metabolite from baicalin. Int. J. Oncol. 2015, 47, 1749–1758.
- Feldo, M.; Wozniak, M.; Wojciak-Kosior, M.; Sowa, I.; Kot-Wasik, A.; Aszyk, J.; Bogucka-Kocka, A. Influence of Diosmin Treatment on the Level of Oxidative Stress Markers in Patients with Chronic Venous Insufficiency. Oxid. Med. Cell Longev. 2018, 2018, 2561705.
- Zhao, Y.; Zhang, J.; Liu, W. Diosmin Regulates Oxidative Stress and Inflammatory Marker Levels in N-Methyl-N-Nitrosourea-Induced Gastric Carcinogenesis in Rats. J. Env. Pathol. Toxicol. Oncol. 2020, 39, 375–384.
- Patel, K.; Gadewar, M.; Tahilyani, V.; Patel, D.K. A review on pharmacological and analytical aspects of diosmetin: A concise report. Chin. J. Integr. Med. 2013, 19, 792–800.
- Russo, R.; Chandradhara, D.; De Tommasi, N. Comparative Bioavailability of Two Diosmin Formulations after Oral Administration to Healthy Volunteers. Molecules 2018, 23, 2174.
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kregiel, D.; Sharifi-Rad, J.; Durazzo, A.; Martins, N. The Therapeutic Potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305.
- Sharma, A.; Kaur, M.; Katnoria, J.K.; Nagpal, A.K. Polyphenols in Food: Cancer Prevention and Apoptosis Induction. Curr. Med. Chem. 2018, 25, 4740–4757.
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A Review on Flavonoid Apigenin: Dietary Intake, ADME, Antimicrobial Effects, and Interactions with Human Gut Microbiota. Biomed. Res. Int. 2019, 2019, 7010467.
- Steed, L.; Christophi, G.P.; Kaiko, G.E. The microbial metabolite desaminotyrosine protects from influenza through type I interferon. Science 2017, 357, 498–502.
- Ashrafizadeh, M.; Ahmadi, Z.; Mohammadinejad, R.; Ghasemipour Afshar, E. Tangeretin: A mechanistic review of its pharmacological and therapeutic effects. J. Basic Clin Physiol. Pharm. 2020, 31, 4.
- Chen, J.; Wang, Y.; Zhu, T.; Yang, S.; Cao, J.; Li, X.; Sun, C. Beneficial Regulatory Effects of Polymethoxyflavone-Rich Fraction from Ougan (Citrus reticulata cv. Suavissima) Fruit on Gut Microbiota and Identification of Its Intestinal Metabolites in Mice. Antioxidants 2020, 9, 831.
- Enomoto, R.; Suzuki, C.; Koshiba, C.; Nishino, T.; Nakayama, M.; Hirano, H.; Lee, E. Wogonin prevents immunosuppressive action but not anti-inflammatory effect induced by glucocorticoid. Ann. N. Y. Acad. Sci. 2007, 1095, 412–417.
- Seong, R.K.; Kim, J.A.; Shin, O.S. Wogonin, a flavonoid isolated from Scutellaria baicalensis, has anti-viral activities against influenza infection via modulation of AMPK pathways. Acta. Virol. 2018, 62, 78–85.
- Xing, S.H.; Wang, M.Y.; Peng, Y.; Chen, D.F.; Li, X.B. Simulated gastrointestinal tract metabolism and pharmacological activities of water extract of Scutellaria baicalensis roots. J. Ethnopharmacol. 2014, 152, 183–189.
- Xing, S.; Wang, M.; Peng, Y.; Li, X. Effects of Intestinal Microecology on Metabolism and Pharmacokinetics of Oral Wogonoside and Baicalin. Nat. Prod. Commun. 2017, 12, 509–514.
- Farkhondeh, T.; Abedi, F.; Samarghandian, S. Chrysin attenuates inflammatory and metabolic disorder indices in aged male rat. Biomed Pharm. 2019, 109, 1120–1125.
- Bolca, S.; Van de Wiele, T.; Possemiers, S. Gut metabotypes govern health effects of dietary polyphenols. Curr. Opin. Biotechnol. 2013, 24, 220–225.
- Yang, G.; Hong, S.; Yang, P.; Sun, Y.; Wang, Y.; Zhang, P.; Gu, Y. Discovery of an ene-reductase for initiating flavone and flavonol catabolism in gut bacteria. Nat. Commun. 2021, 12, 790.
- Pollastri, S.; Tattini, M. Flavonols: Old compounds for old roles. Ann. Bot. 2011, 108, 1225–1233.
- Najmanova, I.; Voprsalova, M.; Saso, L.; Mladenka, P. The pharmacokinetics of flavanones. Crit. Rev. Food Sci. Nutr. 2020, 60, 3155–3171.
- Pabich, M.; Materska, M. Biological Effect of Soy Isoflavones in the Prevention of Civilization Diseases. Nutrients 2019, 11, 1660.
- Guimarães, R.M.; Silva, T.E.; Lemes, A.C.; Boldrin, M.C.F.; Silva Pereira, M.A.; Silva, F.G.; Egea, M.B. Okara: A soybean by-product as an alternative to enrich vegetable paste. LWT Food Sci. Technol. 2018, 92, 593–599.
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779.
- Lin, B.W.; Gong, C.C.; Song, H.F.; Cui, Y.Y. Effects of anthocyanins on the prevention and treatment of cancer. Br. J. Pharmacol. 2017, 174, 1226–1243.
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435.
- Martens, S.; Mithofer, A. Flavones and flavone synthases. Phytochemistry 2005, 66, 2399–2407.
- Debnath, J.; Baehrecke, E.H.; Kroemer, G. Does autophagy contribute to cell death? Autophagy 2005, 1, 66–74.
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516.
- Danial, N.N.; Korsmeyer, S.J. Cell death: Critical control points. Cell 2004, 116, 205–219.
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448.
- Lopez, J.; Tait, S.W.G. Mitochondrial apoptosis: Killing cancer using the enemy within. Br. J. Cancer 2015, 112, 957–962.
- Ponte, L.G.S.; Pavan, I.C.B.; Mancini, M.C.S.; da Silva, L.G.S.; Morelli, A.P.; Severino, M.B.; Simabuco, F.M. The Hallmarks of Flavonoids in Cancer. Molecules 2021, 26, 2029.
- Sabarwal, A.; Agarwal, R.; Singh, R.P. Fisetin inhibits cellular proliferation and induces mitochondria-dependent apoptosis in human gastric cancer cells. Mol. Carcinog. 2017, 56, 499–514.
- Shih, P.H.; Yeh, C.T.; Yen, G.C. Effects of anthocyanidin on the inhibition of proliferation and induction of apoptosis in human gastric adenocarcinoma cells. Food Chem. Toxicol. 2005, 43, 1557–1566.
- Zhang, J.; Wu, D.; Vikash, S.J.; Wang, J.; Yi, J.; Dong, W. Hesperetin Induces the Apoptosis of Gastric Cancer Cells via Activating Mitochondrial Pathway by Increasing Reactive Oxygen Species. Dig. Dis. Sci. 2015, 60, 2985–2995.
- Wang, B.; Zhao, X.H. Apigenin induces both intrinsic and extrinsic pathways of apoptosis in human colon carcinoma HCT-116 cells. Oncol. Rep. 2017, 37, 1132–1140.
- Matson, J.P.; Cook, J.G. Cell cycle proliferation decisions: The impact of single cell analyses. FEBS J. 2017, 284, 362–375.
- Davis, C.D.; Emenaker, N.J.; Milner, J.A. Cellular proliferation, apoptosis and angiogenesis: Molecular targets for nutritional preemption of cancer. Semin. Oncol. 2010, 37, 243–257.
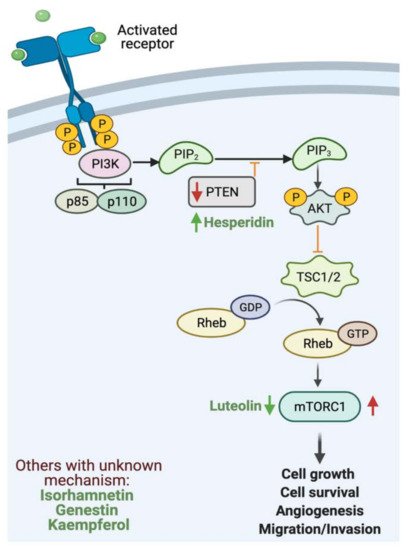
- Engelman, J.A.; Luo, J.; Cantley, L.C. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat. Rev. Genet. 2006, 7, 606–619.
- Guo, H.; German, P.; Bai, S.; Barnes, S.; Guo, W.; Qi, X.; Ding, Z. The PI3K/AKT Pathway and Renal Cell Carcinoma. J. Genet. Genom. 2015, 42, 343–353.
- Hennessy, B.T.; Smith, D.L.; Ram, P.T.; Lu, Y.; Mills, G.B. Exploiting the PI3K/AKT pathway for cancer drug discovery. Nat. Rev. Drug Discov. 2005, 4, 988–1004.
- Yang, J.; Nie, J.; Ma, X.; Wei, Y.; Peng, Y.; Wei, X. Targeting PI3K in cancer: Mechanisms and advances in clinical trials. Mol. Cancer 2019, 18, 26.
- Alshatwi, A.A.; Ramesh, E.; Periasamy, V.S.; Subash-Babu, P. The apoptotic effect of hesperetin on human cervical cancer cells is mediated through cell cycle arrest, death receptor, and mitochondrial pathways. Fundam. Clin Pharm. 2013, 27, 581–592.
- He, P.; Ma, J.; Liu, Y.; Deng, H.; Dong, W. Hesperetin Promotes Cisplatin-Induced Apoptosis of Gastric Cancer In Vitro and In Vivo by Upregulating PTEN Expression. Front. Pharm. 2020, 11, 1326.
- Pu, Y.; Zhang, T.; Wang, J.; Mao, Z.; Duan, B.; Long, Y.; Gao, Z. Luteolin exerts an anticancer effect on gastric cancer cells through multiple signaling pathways and regulating miRNAs. J. Cancer 2018, 9, 3669–3675.
- Aklilu, M.; Kindler, H.L.; Donehower, R.C.; Mani, S.; Vokes, E.E. Phase II study of flavopiridol in patients with advanced colorectal cancer. Ann. Oncol. 2003, 14, 1270–1273.
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA Cancer, J. Clin. 2016, 66, 271–289.
- Ahmad, S.S.; Reinius, M.A.; Hatcher, H.M.; Ajithkumar, T.V. Anticancer chemotherapy in teenagers and young adults: Managing long term side effects. BMJ 2016, 354, i4567.
- Okubo, R.; Kinoshita, T.; Katsumata, N.; Uezono, Y.; Xiao, J.; Matsuoka, Y.J. Impact of chemotherapy on the association between fear of cancer recurrence and the gut microbiota in breast cancer survivors. Brain Behav. Immun. 2020, 85, 186–191.
- Ichim, T.E.; Kesari, S.; Shafer, K. Protection from chemotherapy-and antibiotic-mediated dysbiosis of the gut microbiota by a probiotic with digestive enzymes supplement. Oncotarget 2018, 9, 30919–30935.
- Li, Q.; Ren, L.; Zhang, Y.; Gu, Z.; Tan, Q.; Zhang, T.; Chen, S. P38 Signal Transduction Pathway Has More Cofactors on Apoptosis of SGC-7901 Gastric Cancer Cells Induced by Combination of Rutin and Oxaliplatin. Biomed Res. Int. 2019, 2019, 6407210.
- Ren, L.Q.; Li, Q.; Zhang, Y. Luteolin Suppresses the Proliferation of Gastric Cancer Cells and Acts in Synergy with Oxaliplatin. Biomed Res. Int. 2020, 2020, 9396512.

