 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Shaoyong Yu | + 2906 word(s) | 2906 | 2021-07-06 05:02:47 | | | |
| 2 | Peter Tang | Meta information modification | 2906 | 2021-07-16 10:39:19 | | |
Video Upload Options
Heartburn and non-cardiac chest pain are the predominant symptoms in many esophageal disorders, such as gastroesophageal reflux disease (GERD), non-erosive reflux disease (NERD), functional heartburn and chest pain, and eosinophilic esophagitis (EoE). At present, neuronal mechanisms underlying the process of interoceptive signals in the esophagus are still less clear. Noxious stimuli can activate a subpopulation of primary afferent neurons at their nerve terminals in the esophagus. The evoked action potentials are transmitted through both the spinal and vagal pathways to their central terminals, which synapse with the neurons in the central nervous system to induce esophageal nociception. Over the last few decades, progress has been made in our understanding on the peripheral and central neuronal mechanisms of esophageal nociception. In this review, we focus on the roles of capsaicin-sensitive vagal primary afferent nodose and jugular C-fiber neurons in processing nociceptive signals in the esophagus. We briefly compare their distinctive phenotypic features and functional responses to mechanical and chemical stimulations in the esophagus. Then, we summarize activation and/or sensitization effects of acid, inflammatory cells (eosinophils and mast cells), and mediators (ATP, 5-HT, bradykinin, adenosine, S1P) on these two nociceptive C-fiber subtypes. Lastly, we discuss the potential roles of capsaicin-sensitive esophageal afferent nerves in processing esophageal sensation and nociception. A better knowledge of the mechanism of nociceptive signal processes in primary afferent nerves in the esophagus will help to develop novel treatment approaches to relieve esophageal nociceptive symptoms, especially those that are refractory to proton pump inhibitors.
1. Esophageal Interoception and Nociception
2. Esophageal Primary Afferent Pathways
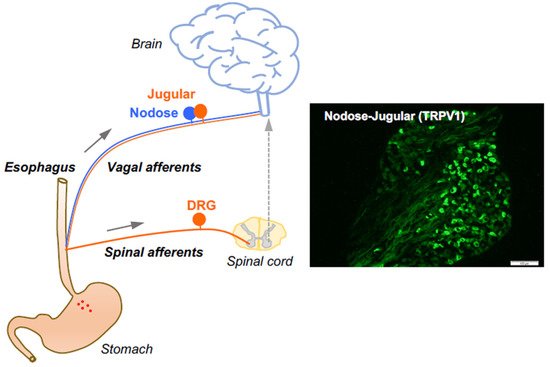
3. Capsaicin-Sensitive Vagal Afferent C-Fibers in the Esophagus
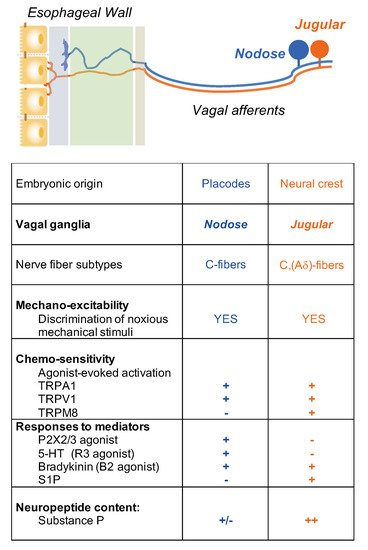
4. Activation and Sensitization of Esophageal Vagal Nodose and Jugular C-Fibers
5. Potential Roles of Capsaicin-Sensitive Afferents in Esophageal Disorders
5.1. Eosinophils
5.2. Mast Cells
5.3. TRPV1 and Acid
5.4. Serotonin (5-Hydroxytrytamine, 5-HT)
5.5. Bradykinin
5.6. Adenosine
5.7. S1P
6. Conclusions
References
- Chen, W.G.; Schloesser, D.; Arensdorf, A.M.; Simmons, J.M.; Cui, C.; Valentino, R.; Gnadt, J.W.; Nielsen, L.; Hillaire-Clarke, C.S.; Spruance, V.; et al. The emerging science of interoception: Sensing, integrating, interpreting, and regulating signals within the self. Trends Neurosci. 2021, 44, 3–16.
- Sherrington, S.C. The Integrative Action of the Nervous System; Yale University Press: New Haven, CT, USA, 1906.
- Brookes, S.J.; Spencer, N.; Costa, M.; Zagorodnyuk, V. Extrinsic primary afferent signalling in the gut. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 286–296.
- Page, A.J.; Blackshaw, L.A. Roles of gastro-oesophageal afferents in the mechanisms and symptoms of reflux disease. Organotypic Models Drug Dev. 2009, 194, 227–257.
- Yu, X.; Yu, M.; Liu, Y.; Yu, S. TRP channel functions in the gastrointestinal tract. Semin. Immunopathol. 2015, 38, 385–396.
- Undem, B.J.; Taylor-Clark, T. Mechanisms underlying the neuronal-based symptoms of allergy. J. Allergy Clin. Immunol. 2014, 133, 1521–1534.
- Yu, S.; Undem, B.J.; Kollarik, M. Vagal afferent nerves with nociceptive properties in guinea-pig oesophagus. J. Physiol. 2005, 563, 831–842.
- Ru, F.; Banovcin, P., Jr.; Kollarik, M. Acid sensitivity of the spinal dorsal root ganglia C-fiber nociceptors innervating the guinea pig esophagus. Neurogastroenterol. Motil. 2015, 27, 865–874.
- Driessen, A.K.; Farrell, M.J.; Mazzone, S.B.; McGovern, A. The Role of the Paratrigeminal Nucleus in Vagal Afferent Evoked Respiratory Reflexes: A Neuroanatomical and Functional Study in Guinea Pigs. Front. Physiol. 2015, 6, 378.
- Woolf, C.J.; Ma, Q. Nociceptors—Noxious stimulus detectors. Neuron 2007, 55, 353–364.
- Yu, S.; Ru, F.; Ouyang, A.; Kollarik, M. 5-Hydroxytryptamine selectively activates the vagal nodose C-fibre subtype in the guinea-pig oesophagus. Neurogastroenterol. Motil. 2008, 20, 1042–1050.
- Yu, S.; Kollarik, M.; Ouyang, A.; Myers, A.C.; Undem, B.J. Mast cell-mediated long-lasting increases in excitability of vagal C fibers in guinea pig esophagus. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G850–G856.
- Yu, S.; Ouyang, A. TRPA1 in bradykinin-induced mechanical hypersensitivity of vagal C fibers in guinea pig esophagus. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G255–G265.
- Yu, S.; Gao, G.; Peterson, B.Z.; Ouyang, A. TRPA1 in mast cell activation-induced long-lasting mechanical hypersensitivity of vagal afferent C-fibers in guinea pig esophagus. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G34–G42.
- Zhang, S.; Grabauskas, G.; Wu, X.; Joo, M.K.; Heldsinger, A.; Song, I.; Owyang, C.; Yu, S. Role of prostaglandin D2 in mast cell activation-induced sensitization of esophageal vagal afferents. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G908–G916.
- Liu, Z.; Hu, Y.; Yu, X.; Xi, J.; Fan, X.; Tse, C.M.; Myers, A.C.; Pasricha, P.J.; Li, X.; Yu, S. Allergen challenge sensitizes TRPA1 in vagal sensory neurons and afferent C-fiber subtypes in guinea pig esophagus. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G482–G488.
- Gu, Q.; Lim, M.E.; Gleich, G.J.; Lee, L.-Y. Mechanisms of eosinophil major basic protein-induced hyperexcitability of vagal pulmonary chemosensitive neurons. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L453–L461.
- Lee, L.-Y.; Gu, Q.; Gleich, G.J. Effects of human eosinophil granule-derived cationic proteins on C-fiber afferents in the rat lung. J. Appl. Physiol. 2001, 91, 1318–1326.
- Yu, S.; Ouyang, A. Effect of synthetic cationic protein on mechanoexcitability of vagal afferent nerve subtypes in guinea pig esophagus. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G1052–G1058.
- Mazzone, S.; Undem, B.J. Vagal afferent innervation of the airways in health and disease. Physiol. Rev. 2016, 96, 975–1024.
- Yu, X.; Hu, Y.; Yu, S. Effects of acid on vagal nociceptive afferent subtypes in guinea pig esophagus. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G471–G478.
- Kollarik, M.; Carr, M.J.; Ru, F.; Ring, C.J.A.; Hart, V.J.; Murdock, P.; Myers, A.C.; Muroi, Y.; Undem, B.J. Transgene expression and effective gene silencing in vagal afferent neuronsin vivousing recombinant adeno-associated virus vectors. J. Physiol. 2010, 588, 4303–4315.
- Mawe, G.M.; Hoffman, J.M. Serotonin signalling in the gut—functions, dysfunctions and therapeutic targets. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 473–486.
- Browning, K.N. Role of central vagal 5-HT3 receptors in gastrointestinal physiology and pathophysiology. Front. Neurosci. 2015, 9, 413.
- Nasr, I.; Attaluri, A.; Hashmi, S.; Gregersen, H.; Rao, S.S. Investigation of esophageal sensation and biomechanical properties in functional chest pain. Neurogastroenterol. Motil. 2010, 22, 520–526.
- Remes-Troche, J.M.; Chahal, P.; Mudipalli, R.; Rao, S.S.C. Adenosine modulates oesophageal sensorimotor function in humans. Gut 2008, 58, 1049–1055.
- Ru, F.; Surdenikova, L.; Brozmanova, M.; Kollarik, M. Adenosine-induced activation of esophageal nociceptors. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G485–G493.
- Brozmanova, M.; Mazurova, L.; Ru, F.; Tatar, M.; Hu, Y.; Yu, S.; Kollarik, M. Mechanisms of the adenosine A2Areceptor-induced sensitization of esophageal C fibers. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G215–G223.
- Trankner, D.; Hahne, N.; Sugino, K.; Hoon, M.A.; Zuker, C. Population of sensory neurons essential for asthmatic hyperreactivity of inflamed airways. Proc. Natl. Acad. Sci. USA 2014, 111, 11515–11520.
- Patil, M.J.; Meeker, S.; Bautista, D.; Dong, X.; Undem, B.J. Sphingosine-1-phosphate activates mouse vagal airway afferent C-fibres via S1PR3 receptors. J. Physiol. 2019, 597, 2007–2019.
- Yu, X.; Patil, M.J.; Yu, M.; Liu, Y.; Wang, J.; Undem, B.J.; Yu, S. Sphingosine-1-phosphate selectively activates vagal afferent C-fiber subtype in guinea pig esophagus. Neurogastroenterol. Motil. 2018, 30, e13359.
- Caterina, M.J.; Julius, D. The vanilloid receptor: A molecular gateway to the pain pathway. Annu. Rev. Neurosci. 2001, 24, 487–517.
- Spechler, S.J. Evaluation and treatment of patients with persistent reflux symptoms despite proton pump inhibitor treatment. Gastroenterol. Clin. N. Am. 2020, 49, 437–450.
- Katzka, D.A.; Pandolfino, J.E.; Kahrilas, P.J. Phenotypes of gastroesophageal reflux disease: Where rome, lyon, and montreal meet. Clin. Gastroenterol. Hepatol. 2020, 18, 767–776.

