Heartburn and non-cardiac chest pain are the predominant symptoms in many esophageal disorders, such as gastroesophageal reflux disease (GERD), non-erosive reflux disease (NERD), functional heartburn and chest pain, and eosinophilic esophagitis (EoE). At present, neuronal mechanisms underlying the process of interoceptive signals in the esophagus are still less clear. Noxious stimuli can activate a subpopulation of primary afferent neurons at their nerve terminals in the esophagus. The evoked action potentials are transmitted through both the spinal and vagal pathways to their central terminals, which synapse with the neurons in the central nervous system to induce esophageal nociception. Over the last few decades, progress has been made in our understanding on the peripheral and central neuronal mechanisms of esophageal nociception. In this review, we focus on the roles of capsaicin-sensitive vagal primary afferent nodose and jugular C-fiber neurons in processing nociceptive signals in the esophagus. We briefly compare their distinctive phenotypic features and functional responses to mechanical and chemical stimulations in the esophagus. Then, we summarize activation and/or sensitization effects of acid, inflammatory cells (eosinophils and mast cells), and mediators (ATP, 5-HT, bradykinin, adenosine, S1P) on these two nociceptive C-fiber subtypes. Lastly, we discuss the potential roles of capsaicin-sensitive esophageal afferent nerves in processing esophageal sensation and nociception. A better knowledge of the mechanism of nociceptive signal processes in primary afferent nerves in the esophagus will help to develop novel treatment approaches to relieve esophageal nociceptive symptoms, especially those that are refractory to proton pump inhibitors.
- capsaicin
- esophagus
- vagal afferent
- nodose
- jugular
- nociception
- interoception
- C-fiber
1. Esophageal Interoception and Nociception
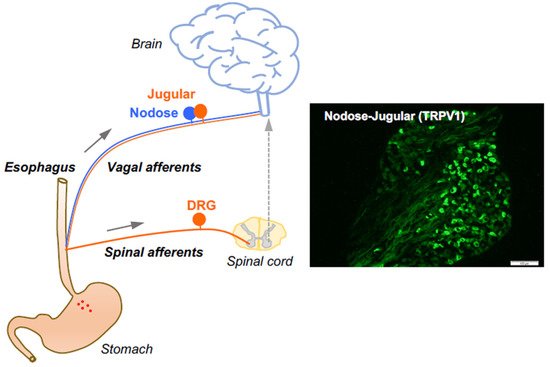
2. Esophageal Primary Afferent Pathways
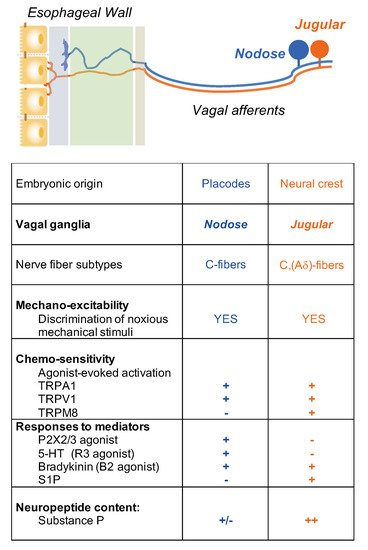
3. Capsaicin-Sensitive Vagal Afferent C-Fibers in the Esophagus
4. Activation and Sensitization of Esophageal Vagal Nodose and Jugular C-Fibers