Membrane proteins (MPs) are essential for cellular functions. Understanding the functions of MPs is crucial as they constitute an important class of drug targets. However, MPs are a challenging class of biomolecules to analyze because they cannot be studied outside their native environment. Their structure, function and activity are highly dependent on the local lipid environment, and these properties are compromised when the protein does not reside in the cell membrane. Mammalian cell membranes are complex and composed of different lipid species. Model membranes have been developed to provide an adequate environment to envisage MP reconstitution. Among them, tethered-Bilayer Lipid Membranes (tBLMs) appear as the best model because they allow the lipid bilayer to be decoupled from the support. Thus, they provide a sufficient aqueous space to envisage the proper accommodation of large extra-membranous domains of MPs, extending outside. Additionally, as the bilayer remains attached to tethers covalently fixed to the solid support, they can be investigated by a wide variety of surface-sensitive analytical techniques.
1. Introduction
Cellular membranes, and more particularly the plasma membrane, are of upmost importance in the living cells. Hosting a vast plethora of proteins, plasma membrane not only serves as a physical boundary, but also mediates exchange processes between the cell and the extracellular matrix. Cellular membranes are also essential inside the cell. They aid the different organelles to carry out their cellular functions. Furthermore, many vital biochemical processes essential for cell life are managed by the biological membranes.
Only a few nanometers thick, biological membranes are very complex in terms of composition but exhibit a perfect organization at the molecular level
[1]. Lipids, held together by hydrophobic interactions, play a structural role by forming a continuous self-assembled bilayer acting as a passive diffusion barrier. Proteins associated with the membrane, either transmembrane proteins or peripheral membrane proteins, respectively embedded within the lipid bilayer or transiently associated with it, represent ~30% of the open reading frames in complex organisms
[2]. Due to their abundance, they are the key factors of the cell metabolism, involving cell–cell communication, cell adhesion, nutrient import, signal transduction, biocatalysis processes, energy production and others
[3]. As a result, membrane receptors are currently the target of over 60% of medicinal drugs
[4][5].
Nowadays, cell membranes are no longer considered as a simple double lipid layer but as a set of complex and dynamic protein–lipid structures and segregated microdomains, that serve as functional spatiotemporal platforms for the interaction of lipids and proteins involved in cellular signaling pathways
[6][7][8][9][10]. The membrane composition, and therefore the overall function of the cell membranes, is altered in a wide range of human diseases, including cancer, neurodegenerative disorders, cardiovascular pathologies, obesity, etc. A lipid alteration can affect the localization and activity of transmembrane proteins and thus impact on the intracellular cell signaling. From this belief, a new concept of membrane lipid therapy (MLT) has emerged
[11][12] with the idea that lipid components of biological membranes can also be selectively targeted to induce membrane disorder and reverse the malfunction
[13]. This approach now represents a target of choice for pharmaceutical companies
[14]. Hence, investigating membranes and membrane proteins (MPs), including lipid–lipid, protein–lipid or ligand–protein receptor interactions, is of critical importance. However, due to their complexity, in situ investigations to unlock the secret of the biological membranes remains a great challenge. In this respect, the development of artificial models that mimic the cell membrane by constitution and composition, is an asset to study biological membrane properties.
In the plasma membrane, hundreds of different lipid species can be found. For instance, some of them have a negative charge, which can promote interactions with positively charged amino acids in proteins
[11]. Depending on the size of their polar head group, certain lipids allow docking of bulky protein lipid anchors or form tightly packed areas to help some membrane proteins to bind to regions where these lipids are abundant. It is now well-accepted that the membrane lipid composition may have a profound role in membrane functioning and cell signaling
[13]. In this respect, the crucial role of non-bilayer lipids present in large amounts in biological membranes on the MP activities must be underlined
[15][16]. Conversely, reconstitution of functional membrane proteins after in vitro production or purification is challenging. Due to their amphiphilic nature, they are prone to early denaturation during in vitro handling. To properly evaluate their functionality, they require a native lipid environment. Ideally, MPs should be reconstituted in natural lipid extracts as it is now well-known that lipids in the immediate vicinity of membrane proteins influence their activity
[13][15][17]. As a result, there is a great need to develop biomimetic membrane platforms, in which, not only one but several purified membrane lipid components can be used for in vitro reconstitution, and in which reincorporated membrane proteins can retain their structural integrity and functional activity.
Different types of models have been developed through the years to mimic cell membranes as well as possible and reproduce the basic functions of cell membranes. These models are solid-supported lipid membranes
[18][19], polymer-cushioned membranes
[20][21][22], hybrid lipid bilayers
[23][24][25], free-standing lipid layers or suspended-lipid bilayers
[26][27][28] and tethered-bilayer lipid membranes or tBLMs
[29][30][31][32][33][34][35][36][37][38][39]. All these models are suitable for systematic studies of different types of membrane-related processes and provide the lipid environment required for the study of membrane-associated proteins. They correspond to models of planar membranes confined to a solid support and localized at the bulk interface, allowing the application of a manifold of surface-sensitive techniques for their own characterization
[40] or biosensing applications
[41][42][43].
Besides all these advantages, tBLMs appear as very attractive platforms for the reinsertion of transmembrane proteins. Because they are lifted from the support, they best mimic the cellular environment, and transmembrane proteins with protrudant domains extending outside the membrane can “comfortably” reinsert into the bilayer without steric hindrance or loss of mobility due to a close contact of the membrane with the support
[44].
2. Design of Tethered-Bilayer Lipid Membranes
tBLMs are a natural progression from the planar supported lipid bilayers (SLBs), first reported by McConnell et al.
[19]. SLBs, classically obtained by the spreading of small unilamellar vesicles on hydrophilic solid supports
[45][46][47], including glass, mica, titanium and silicon oxides, or gold (for recent reviews see articles by Lind & Cardenas 2016
[48] and Clifton et al. 2020
[40]), consist in a lipid bilayer deposited and separated from the solid substrate by an ultrathin film of water (0.5–2 nm)
[45][49][50][51]. This aqueous layer acts as a lubricant and confers to SLBs the fluidity required for lateral diffusion in 2D space
[45]. In this model, lateral and rotational mobility of individual lipids are preserved and anything linked to the phospholipids or glycolipids in the upper leaflet retains this mobility
[52]. Given this key feature, SLBs have been used extensively over the past decades to study the spatially and temporally regulated lipid–lipid or lipid–protein lateral interactions
[53][54][55][56], lipid segregation
[57][58][59], protein clustering and cell adhesion
[60][61][62][63] and membrane dynamics
[64][65][66].
However, the close proximity of SLBs to the substrate affects the diffusion of lipids and proteins, which is more than twice slower than in free-standing bilayers under the same conditions
[67]. This limitation is due to the fact that the substrate exerts a greater influence on the behavior of the proximal leaflet than the distal leaflet of the SLBs, due to its closer proximity to the surface
[40][68][69]. The roughness of the substrate and the complementarity between the surface and lipid charge will determine the magnitude of this surface influence
[50]. Furthermore, SLBs suffer from the crucial drawback of not possessing a large hydration reservoir on both sides of the membranes, which limit examination of transmembrane proteins. The fundamental requirement for a membrane protein to function properly is to be surrounded by buffered-saline solution on both sides of membranes
[70]. In SLBs, hydration layer is often not thick enough for proper folding of large extra-membranous domains of transmembrane proteins, which can extend to several tens of nanometers far out from the bilayer
[18]. The limited membrane-substrate distance, which can lead to strong non-physiological interactions with the solid support
[18][20], can cause both a loss of protein dynamics and a partial loss of its functionality, or even complete denaturation of the protein
[71][72][73]. In addition, anionic substrates (such as quartz, mica, silica, silicon oxide) may hinder (in the absence of divalent cations) the formation of SLBs enriched in negatively charged lipids in the proximal leaflet, due to electrostatic repulsions
[46][66][74]. However, negatively charged lipids, like phosphatidylglycerol (PG), phosphatidylinositol (PI), cardiolipin (CL) or even lipopolysaccharide (LPS) in the Gram-negative bacteria membrane, are important signaling lipids which can trigger membrane protein association and affect membrane-regulated pathways. They are essential in membrane function, and studies of the membrane phenomenon regulated by these lipids are becoming crucial for a realistic understanding of membrane-related events. The use of mimetic sample systems with ever greater biological precision in lipid composition is now required.
For all these reasons, more advanced planar model membranes are currently in development with the aim to create more accurate biomimetic systems adapted for integral (trans-)membrane protein characterization, where the substrate interactions are minimized and large solution reservoirs on both sides of the bilayer are provided. In tBLMs, the lipid bilayer is separated from the surface of the substrate by insertion of a soft and flexible hydrophilic layer of “tethering” molecules that anchor the proximal leaflet
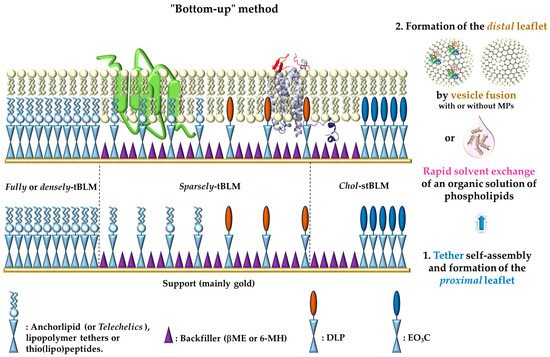
[31][34][71][72][75][76]. This layer solves the substrate proximity by lifting the membrane off the surface and provides a reservoir underneath the bilayer in which the membrane proteins can fold into a native-like conformation, while keeping the membrane anchored to the support ().
Figure 1. Different types of
tethered-Bilayer Lipid Membranes (or tBLMs) obtained by a “bottom-up” approach.

: represents either anchorlipid (i.e., Telechelics) mainly 2,3-di-
O-phytanyl-
sn-glycerol-1-tetraethylene glycol-
d,
l-α-lipoic acid ester lipid (DPhyTL), lipopolymer tethers or thio(lipo)peptides;
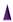
: Backfillers, mainly β-mercaptoethaonl (βME) or 6-mercaptohexanol (6-MH);
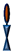
: benzyl-disulfide (tetra-ethyleneglycol)
n=2 C20-phytanyl tether (or DLP);
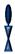
: ethyleneoxy-linked cholesterol (or EO
3C).
The large variety of assembling molecules capable of forming a tethering layer offers multiple possibilities for fine-tuning the properties of tBLMs
[37]. Whatever its proper nature, the role of the anchor (spacer group) is multiple. It should at the same time (i) maintain bilayer fluidity and provide a sufficient well-hydrated sub-membrane space to accommodate incorporated proteins, (ii) cover small surface roughness features in order to reduce the hydrophobic influence of the metal surface and the unfavorable frictions to the support, (iii) provide a hydrated reservoir between the substrate and the membrane, and (iv) supply ample space to harbor membrane protein ectodomains. The different types of tBLMs vary mainly in the chemical structure of the tethers and in their density, two factors significantly influencing the structural characteristics of the bilayer as well as the functional reincorporation of membrane components
[51]. Ideally, and in order to mimic a natural membrane, a tBLM should have a high electrical impedance and a low capacitance—to be sure that transport across the membrane is mainly due to the function of the embedded protein, as well as high fluidity and high sub-membrane hydration—to ensure protein function. However, increases in membrane hydration and fluidity are generally accompanied by a reduction of the electrical sealing properties, resulting from a higher defect density
[77][78]. Subsequently, applying ultrasensitive surface imaging techniques allowing the direct characterization of all the steps of the tBLM formation with a high lateral resolution could lead to an optimization, step-by-step during the tBLM building, in order to reduce the number of defects.
3. Characterization of Tethered-Bilayer Lipid Membranes
Because they are firmly held in place, tBLMs are considerably more robust than supported lipid bilayers such as black or bilayer lipid membranes (BLMs)
[79][80], also renamed free-standing lipid layer or suspended-lipid bilayers, which are originally formed across on a microsized-aperture, and more recently, on nanopores
[26][27][28]. Generally speaking, tBLMs typically show a high robustness and long-term stability and hence, they are accessible to a portfolio of different analytical tools operating at a bulk interface
[42]. They include imaging techniques, like
atomic force microscopy (AFM)
[30][81][82][83][84][85][86][87] and
fluorescence microscopy (FM)
[34][87][88][89],
fluorescence recovering after photobleaching (FRAP)
[20][35][81][87][90][91][92][93],
neutron reflectometry (NR)
[77][94][95][96][97][98][99][100][101] and
X-ray photoelectron spectroscopy (XPS)
[20][82][86][87][102][103], spectroscopic techniques such as
ellipsometry [94][104],
infrared reflection absorption spectroscopy (IRRAS)
[96][105][106] or
surface-enhanced infrared absorption spectroscopy (SEIRAS)
[107][108][109][110],
surface plasmon resonance (SPR)
[30][34][35][37][39][111][112][113] or
quartz crystal micro-balance with dissipation monitoring (QCM-D)
[30][39][85][112], as well as electrochemical methods such as
electrochemical impedance spectroscopy (EIS)
[24][32][41][88][94][108][109][114][115][116][117] and current-voltage (CV) analysis
[118][119][120][121][122][123]. These techniques, sensitive to net changes in packing or interfacial mass (QCM-D or ellipsometry), bilayer morphology (AFM, FM), the presence of chemical groups (IR, XPS), the structure and composition (NR), have been used so far to evaluate the full picture of the lipid membranes (i.e., structure, composition and functional properties) and represent a very powerful combination to unravel the mechanism of biomolecular interactions.
While SPR and QCM-D allow real-time monitoring of the tBLM formation in a label-free format, fluorescence microscopy (FM) and FRAP investigate domain morphology and membrane dynamics with the measurement of the lateral diffusion of lipids, respectively. AFM has been used to gather surface details in terms of occurrence of peculiar structure and defects. One of the unique features of this latter technique is that it can measure surface forces with a nanometer lateral resolution. Recently, single-molecule force spectroscopy (FS) measurements have provided in-depth insight to assess the orientation of reconstituted transmembrane proteins in tBLMs
[86]. NR also provides high resolution structural information on lipid bilayer stacking and internal distribution of components after interaction between intrinsic proteins and disordered membrane
[124]. EIS is an excellent tool to characterize the electric properties of membrane including resistance and capacitance.
This large panel allows a fine characterization of tBLMs during and after their formation, in terms of structure, (optical or acoustic) thickness, fluidity and sealing
[125]. It shows that the chemical nature of the sub-membrane space has a significant impact on both the structure of the lipid bilayer and the functional incorporation of membrane components
[51]. The possible combination of multiple complementary measurements with biologically accurate samples is key for a realistic understanding of membrane related phenomena. Only through the use of complementary techniques, such as the ones hereby mentioned, does it become a realistic aim to resolve the relative position, orientation and distribution of the membrane components to obtain detailed information on molecular mechanisms by which peptides, proteins or other chemical compounds (e.g., drugs) interact with biomembranes. , adapted from Rossi and Chopineau
[126], Sondhi et al.
[70], Clifton et al.
[40], presents a synoptic of the characteristics of all the techniques useful for the study of supported planar membrane models, including tBLMs. For more details, see the recent review by Clifton et al.
[40], which presents the main information that can be deduced from model membranes due to the different surface-sensitive techniques listed above.
Table 1. Characteristics of main surface-sensitive analytical techniques useful for investigation of supported planar model membranes included tBLMs (adapted from
[40][70][126]).
| Techniques |
Bilayer Characterization |
Surfaces |
| Surface plasmon resonance (SPR) imaging |
Optical thickness of the bilayer, highly sensitive real-time monitoring of interactions without labeling of the analytes or the ligand, real-time monitoring of bilayer formation |
Gold, silver, aluminum |
| Quartz crystal microbalance with dissipation (QCM-D) |
Interfacial wet mass determination and viscoelasticity (dissipation sensitive to viscoelastic properties of the adsorbed material), (acoustic) film thickness, real-time monitoring of bilayer formation |
Gold, SiO2, mica, metal oxides |
| Imaging ellipsometry (IE) |
Indirect quantitative characterization of structural and functional properties of bilayers such as thickness and dry adsorbed mass (i.e., lipids in the adsorbed layer), anisotropy (lateral uniformity and phase separation), molecular area, and receptor-protein interaction affinities. Real-time large area imaging with high sensitivity |
Oxide (silicon) substrates |
| Fluorescence recovery after photobleaching (FRAP) |
Dynamics, fluidity, and mobility characterisation of lipids and proteins (peripheral or integral), intergrity of artificial membranes |
Optically transparent substrates: glass, silica, silcon, gold |
| Electrochemical impedance spectroscopy (EIS) |
Electrical properties (resistance and capacitance) of lipid bilayer membranes, formation process in real-time, stability of the membrane, characterization of incorporated ion channels |
Gold, silicon |
| Atomic force microscopy (AFM) |
In-plane structure and morphology: surface roughness determination, investigation of bilayer surface at the nanoscale range in real-time and in aqueous environment, direct measure of physical properties at high spatial resolution, phase separation (domain formation) and quantification of bilayer thickness |
Atomically flat surfaces: mica, silicon, quartz, flat gold |
| (AFM) single-molecule Force Spectroscopy (FS) |
Membrane stiffness and mechanical stability on the nanometer length scale, in-depth insight of the orientation of reconstituted transmembrane proteins |
Mica, silicon, quartz, flat gold |
| Neutron Reflectometry (NR) |
Non-damaging technique giving high structural information on lipid bilayer and internal distribution of components (lipid or protein) within the bilayer (thickness of stratified layers normal to the interface), roughness and interaction with inserted proteins (easy differentiation of lipid and polypeptide components across the membrane structure after interaction) |
Gold, silicon |
| X-ray photoelectron spectroscopy (XPS) |
Provides quantitaive analysis of elemental composition of a surface and its chemical state |
Quartz |
| Grazing incidence small angle neutron or X-ray scattering (GISANS and GISAXS) |
Non-destructive method for the structural investigation of biomembranes and mixed lipids systems with different topologies |
Performed in quartz glass |
 +1 credit
+1 credit