 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Flávia Rezende | + 2375 word(s) | 2375 | 2021-08-11 08:59:08 | | | |
| 2 | Vivi Li | Meta information modification | 2375 | 2021-08-24 04:54:40 | | |
Video Upload Options
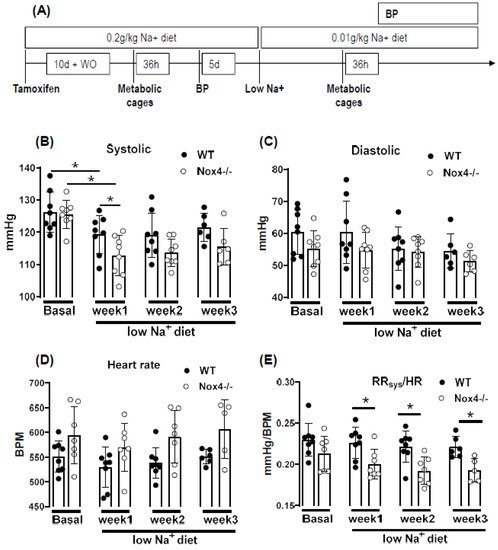
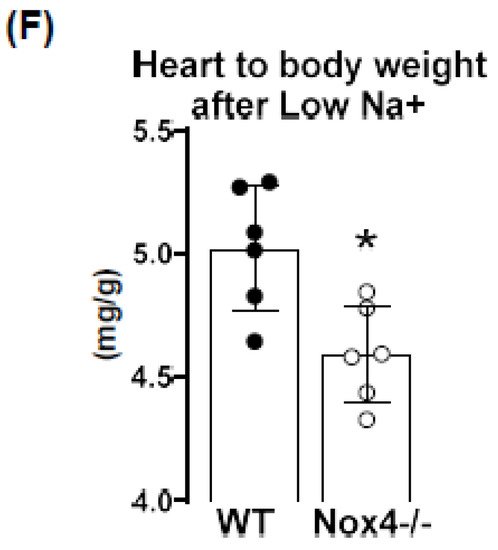
The NADPH oxidase Nox4 is a hydrogen peroxide (H2O2)-producing enzyme, with the highest expression in the kidney. As the kidney is involved in volume and blood pressure control through sodium handling, we set out to determine the impact of a low sodium diet on these parameters in WT and Nox4-/- mice. Nox4 expression in the murine kidney was restricted to the proximal tubule. Nevertheless, low-sodium-induced weight loss and sodium sparing function was similar in WT and Nox4-/- mice, disputing an important function of renal Nox4 in sodium handling. In contrast, a low sodium diet resulted in a reduction in systolic blood pressure in Nox4-/- as compared to WT mice. This was associated with a selectively lower pressure to heart-rate ratio, as well as heart to body weight ratio. In general, a low sodium diet leads to activation of sympathetic tone and the renin angiotensin system, which subsequently increases peripheral resistance. Our observations suggest that the control by this system is attenuated in Nox4-/- mice, resulting in lower blood pressure in response to low sodium.
1. Introduction
2. Nox4 Expression Is Restricted to Proximal Tubule Cells
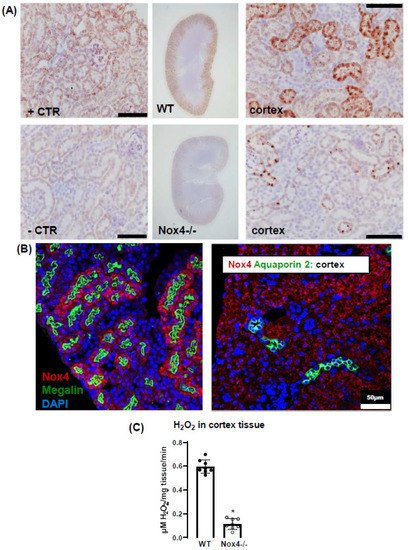
3. Nox4 Contributes to H2O2 Production of the Renal Cortex
4. Knockout of Nox4 Lowers Blood Pressure and Cardiac Mass in Response to Low Sodium Diet
5. Discussion
References
- Araujo, M.; Wilcox, C.S. Oxidative Stress in Hypertension: Role of the Kidney. Antioxidants Redox Signal. 2014, 20, 74–101.
- Gill, P.S.; Wilcox, C.S. NADPH Oxidases in the Kidney. Antioxidants Redox Signal. 2006, 8, 1597–1607.
- Chabrashvili, T.; Tojo, A.; Onozato, M.L.; Kitiyakara, C.; Quinn, M.; Fujita, T.; Welch, W.J.; Wilcox, C.S. Expression and Cellular Localization of Classic NADPH Oxidase Subunits in the Spontaneously Hypertensive Rat Kidney. Hypertension 2002, 39, 269–274.
- Sedeek, M.; Hébert, R.L.; Kennedy, C.R.; Burns, K.D.; Touyz, R.M. Molecular mechanisms of hypertension: Role of Nox family NADPH oxidases. Curr. Opin. Nephrol. Hypertens. 2009, 18, 122–127.
- Touyz, R.M.; Alves-Lopes, R.; Rios, F.; De Lucca Camargo, L.; Anagnostopoulou, A.; Arner, A.; Montezano, A.C. Vascular smooth muscle contraction in hypertension. Cardiovasc. Res. 2018, 114, 529–539.
- Makino, A.; Skelton, M.M.; Zou, A.-P.; Cowley, A.W. Increased Renal Medullary H2O2 Leads to Hypertension. Hypertension 2003, 42, 25–30.
- Makino, A.; Skelton, M.M.; Zou, A.-P.; Roman, R.J.; Cowley, A.W. Increased Renal Medullary Oxidative Stress Produces Hypertension. Hypertension 2002, 39, 667–672.
- Kopkan, L.; Castillo, A.; Navar, L.G.; Majid, D.S.A. Enhanced superoxide generation modulates renal function in ANG II-induced hypertensive rats. Am. J. Physiol. Physiol. 2006, 290, F80–F86.
- Babelova, A.; Avaniadi, D.; Jung, O.; Fork, C.; Beckmann, J.; Kosowski, J.; Weissmann, N.; Anilkumar, N.; Shah, A.; Schaefer, L.; et al. Role of Nox4 in murine models of kidney disease. Free. Radic. Biol. Med. 2012, 53, 842–853.
- Prior, K.-K.; Leisegang, M.S.; Josipovic, I.; Löwe, O.; Shah, A.M.; Weissmann, N.; Schröder, K.; Brandes, R.P. CRISPR/Cas9-mediated knockout of p22phox leads to loss of Nox1 and Nox4, but not Nox5 activity. Redox Biol. 2016, 9, 287–295.
- Takac, I.; Schröder, K.; Zhang, L.; Lardy, B.; Anilkumar, N.; Lambeth, J.D.; Shah, A.; Morel, F.; Brandes, R.P. The E-loop Is Involved in Hydrogen Peroxide Formation by the NADPH Oxidase Nox4. J. Biol. Chem. 2011, 286, 13304–13313.
- Löwe, O.; Rezende, F.; Heidler, J.; Wittig, I.; Helfinger, V.; Brandes, R.P.; Schröder, K. BIAM switch assay coupled to mass spectrometry identifies novel redox targets of NADPH oxidase 4. Redox Biol. 2019, 21, 101125.
- Santos, C.X.; Hafstad, A.D.; Beretta, M.; Zhang, M.; Molenaar, C.; Kopec, J.; Fotinou, D.; Murray, T.V.; Cobb, A.M.; Martin, D.; et al. Targeted redox inhibition of protein phosphatase 1 by Nox4 regulates eIF 2α-mediated stress signaling. EMBO J. 2016, 35, 319–334.
- Yang, Q.; Wu, F.-R.; Wang, J.-N.; Gao, L.; Jiang, L.; Li, H.-D.; Ma, Q.; Liu, X.-Q.; Wei, B.; Zhou, L.; et al. Nox4 in renal diseases: An update. Free Radic. Biol. Med. 2018, 124, 466–472.
- Holterman, C.; Read, N.C.; Kennedy, C.R.J. Nox and renal disease. Clin. Sci. 2014, 128, 465–481.
- Thallas-Bonke, V.; Jandeleit-Dahm, K.A.; Cooper, M.E. Nox-4 and progressive kidney disease. Curr. Opin. Nephrol. Hypertens. 2015, 24, 74–80.
- Liang, Y.; Liu, H.; Fang, Y.; Lin, P.; Lu, Z.; Zhang, P.; Jiao, X.; Teng, J.; Ding, X.; Dai, Y. Salvianolate ameliorates oxidative stress and podocyte injury through modulation of NOX4 activity in db/db mice. J. Cell. Mol. Med. 2021, 25, 1012–1023.
- Rajaram, R.D.; Dissard, R.; Faivre, A.; Ino, F.; Delitsikou, V.; Jaquet, V.; Cagarelli, T.; Lindenmeyer, M.; Jansen-Duerr, P.; Cohen, C.; et al. Tubular NOX4 expression decreases in chronic kidney disease but does not modify fibrosis evolution. Redox Biol. 2019, 26, 101234.
- Shi, Q.; Lee, D.-Y.; Féliers, D.; Abboud, H.E.; Bhat, M.A.; Gorin, Y. Interplay between RNA-binding protein HuR and Nox4 as a novel therapeutic target in diabetic kidney disease. Mol. Metab. 2020, 36, 100968.
- Jha, J.C.; Thallas-Bonke, V.; Banal, C.; Gray, S.P.; Chow, B.S.M.; Ramm, G.; Quaggin, S.E.; Cooper, M.E.; Schmidt, H.H.H.W.; Jandeleit-Dahm, K.A. Podocyte-specific Nox4 deletion affords renoprotection in a mouse model of diabetic nephropathy. Diabetologia 2016, 59, 379–389.
- You, Y.-H.; Quach, T.; Saito, R.; Pham, J.; Sharma, K. Metabolomics Reveals a Key Role for Fumarate in Mediating the Effects of NADPH Oxidase 4 in Diabetic Kidney Disease. J. Am. Soc. Nephrol. 2015, 27, 466–481.
- Thallas-Bonke, V.; Tan, S.M.; Lindblom, R.S.; Snelson, M.; Granata, C.; Jha, J.C.; Sourris, K.C.; Laskowski, A.; Watson, A.; Tauc, M.; et al. Targeted deletion of nicotinamide adenine dinucleotide phosphate oxidase 4 from proximal tubules is dispensable for diabetic kidney disease development. Nephrol. Dial. Transplant. 2021, 36, 988–997.
- Cowley, A.W.; Abe, M.; Mori, T.; O’Connor, P.M.; Ohsaki, Y.; Zheleznova, N.N. Reactive oxygen species as important determinants of medullary flow, sodium excretion, and hypertension. Am. J. Physiol. Physiol. 2015, 308, F179–F197.
- Serrander, L.; Cartier, L.; Bedard, K.; Banfi, B.; Lardy, B.; Plastre, O.; Sienkiewicz, A.; Fórró, L.; Schlegel, W.; Krause, K.-H. NOX4 activity is determined by mRNA levels and reveals a unique pattern of ROS generation. Biochem. J. 2007, 406, 105–114.
- Cowley, J.A.W.; Yang, C.; Zheleznova, N.N.; Staruschenko, A.; Kurth, T.; Rein, L.; Kumar, V.; Sadovnikov, K.; Dayton, A.; Hoffman, M.; et al. Evidence of the Importance of Nox4 in Production of Hypertension in Dahl Salt-Sensitive Rats. Hypertension 2016, 67, 440–450.
- Park, J.; Shrestha, R.; Qiu, C.; Kondo, A.; Huang, S.; Werth, M.; Li, M.; Barasch, J.; Suszták, K. Single-cell transcriptomics of the mouse kidney reveals potential cellular targets of kidney disease. Science 2018, 360, 758–763.
- Khodo, S.N.; Dizin, E.; Sossauer, G.; Szanto, I.; Martin, P.-Y.; Feraille, E.; Krause, K.H.; De Seigneux, S. NADPH-Oxidase 4 Protects against Kidney Fibrosis during Chronic Renal Injury. J. Am. Soc. Nephrol. 2012, 23, 1967–1976.
- Eshbach, M.L.; Sethi, R.; Avula, R.; Lamb, J.; Hollingshead, D.J.; Finegold, D.N.; Locker, J.D.; Chandran, U.R.; Weisz, O.A. The transcriptome of the Didelphis virginiana opossum kidney OK proximal tubule cell line. Am. J. Physiol. Physiol. 2017, 313, F585–F595.
- McDonough, A.A. Mechanisms of proximal tubule sodium transport regulation that link extracellular fluid volume and blood pressure. Am. J. Physiol. Integr. Comp. Physiol. 2010, 298, R851–R861.
- Satriano, J.; Wead, L.; Cardús, A.; Deng, A.; Boss, G.R.; Thomson, S.C.; Blantz, R.C. Regulation of ecto-5′-nucleotidase by NaCl and nitric oxide: Potential roles in tubuloglomerular feedback and adaptation. Am. J. Physiol. Physiol. 2006, 291, F1078–F1082.
- Infanger, D.W.; Cao, X.; Butler, S.D.; Burmeister, M.A.; Zhou, Y.; Stupinski, J.A.; Sharma, R.V.; Davisson, R.L. Silencing Nox4 in the Paraventricular Nucleus Improves Myocardial Infarction–Induced Cardiac Dysfunction by Attenuating Sympathoexcitation and Periinfarct Apoptosis. Circ. Res. 2010, 106, 1763–1774.
- Sun, Q.-A.; Hess, D.T.; Nogueira, L.; Yong, S.; Bowles, D.E.; Eu, J.; Laurita, K.R.; Meissner, G.; Stamler, J.S. Oxygen-coupled redox regulation of the skeletal muscle ryanodine receptor-Ca2+ release channel by NADPH oxidase 4. Proc. Natl. Acad. Sci. USA 2011, 108, 16098–16103.
- Evangelista, A.M.; Thompson, M.D.; Bolotina, V.M.; Tong, X.; Cohen, R.A. Nox4- and Nox2-dependent oxidant production is required for VEGF-induced SERCA cysteine-674 S-glutathiolation and endothelial cell migration. Free Radic. Biol. Med. 2012, 53, 2327–2334.
- Ding, R.; Jiang, H.; Sun, B.; Wu, X.; Li, W.; Zhu, S.; Liao, C.; Zhong, Z.; Chen, J. Advanced oxidation protein products sensitized the transient receptor potential vanilloid 1 via NADPH oxidase 1 and 4 to cause mechanical hyperalgesia. Redox Biol. 2016, 10, 1–11.
- Lin, C.-S.; Lee, S.-H.; Huang, H.-S.; Chen, Y.-S.; Ma, M.-C. H2O2 generated by NADPH oxidase 4 contributes to transient receptor potential vanilloid 1 channel-mediated mechanosensation in the rat kidney. Am. J. Physiol. Physiol. 2015, 309, F369–F376.
- Jiang, Q.; Fu, X.; Tian, L.; Chen, Y.; Yang, K.; Chen, X.; Zhang, J.; Lu, W.; Wang, J. NOX4 Mediates BMP4-Induced Upregulation of TRPC1 and 6 Protein Expressions in Distal Pulmonary Arterial Smooth Muscle Cells. PLoS ONE 2014, 9, e107135.
- Kim, E.Y.; Anderson, M.; Dryer, S.E. Insulin increases surface expression of TRPC6 channels in podocytes: Role of NADPH oxidases and reactive oxygen species. Am. J. Physiol. Physiol. 2012, 302, F298–F307.
- Clempus, R.E.; Sorescu, D.; Dikalova, A.E.; Pounkova, L.; Jo, P.; Sorescu, G.P.; Schmidt, H.H.H.; Lassègue, B.; Griendling, K.K. Nox4 Is Required for Maintenance of the Differentiated Vascular Smooth Muscle Cell Phenotype. Arter. Thromb. Vasc. Biol. 2007, 27, 42–48.
- Hilenski, L.L.; Clempus, R.E.; Quinn, M.; Lambeth, J.D.; Griendling, K.K. Distinct Subcellular Localizations of Nox1 and Nox4 in Vascular Smooth Muscle Cells. Arter. Thromb. Vasc. Biol. 2004, 24, 677–683.
- Lee, M.; Martín, A.S.; Valdivia, A.; Martin-Garrido, A.; Griendling, K.K. Redox-Sensitive Regulation of Myocardin-Related Transcription Factor (MRTF-A) Phosphorylation via Palladin in Vascular Smooth Muscle Cell Differentiation Marker Gene Expression. PLoS ONE 2016, 11, e0153199.
- Martin-Garrido, A.; Brown, D.I.; Lyle, A.N.; Dikalova, A.; Seidel-Rogol, B.; Lassègue, B.; Martín, A.S.; Griendling, K.K. NADPH oxidase 4 mediates TGF-β-induced smooth muscle α-actin via p38MAPK and serum response factor. Free Radic. Biol. Med. 2011, 50, 354–362.
- Panico, C.; Luo, Z.; Damiano, S.; Artigiano, F.; Gill, P.; Welch, W.J. Renal Proximal Tubular Reabsorption Is Reduced In Adult Spontaneously Hypertensive Rats. Hypertension 2009, 54, 1291–1297.
- Liu, J.; Yan, Y.; Liu, L.; Xie, Z.; Malhotra, D.; Joe, B.; Shapiro, J.I. Impairment of Na/K-ATPase Signaling in Renal Proximal Tubule Contributes to Dahl Salt-sensitive Hypertension. J. Biol. Chem. 2011, 286, 22806–22813.


