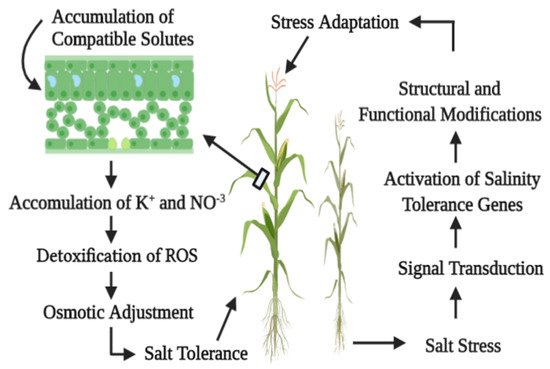
1.1. Overview of Salinity
Abiotic stresses like salinity, drought, and high temperature have undesirable effects on crop productivity and quality, and negative trends in sustainable agriculture
[1]. Salinity in particular is an important limiting factor, causing low yield with inferior quality. Climate change is considered one of the major contributing factors to soil salinization, leading to land degradation and desertification
[2]. According to Flowers et al.
[3], high salt concentration is responsible for negative impacts on 7% of total land surface, and 5% of cultivated land. Poor irrigation water quality is another important factor contributing to soil salinization
[4]. For these reasons, soil salinization is a reported major cause of reductions in the productivity of irrigated and rainfed lands of the world
[5][6].
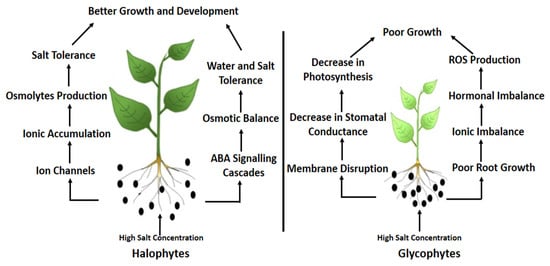
The adverse effects of salinization on plants are evident from negative growth trends from alteration or inhibition of biochemical and physiological processes. Plants can be classified as glycophytes or halophytes by their ability to survive under high salt concentrations
[7]. Glycophytes are plants that are severely affected by saline conditions both at the cellular and whole-plant level. Under saline conditions, these plants exhibit greater accumulation of solutes, and ionic and osmotic stresses confer nutritional imbalances, which limit the productivity of these plants. The majority of terrestrial plants are glycophytes, including crop plants
[1][8].
Conversely, halophytes regulate their biochemical and physiological processes through ionic compartmentalization, production of osmolytes and compatible solutes, enzymatic changes, and absorption of selective ions. These adaptations promote seed germination, succulence, and salt exclusion for these plants in a saline environment
[9][10]. Halophyte succulence keeps the proportion of ions to water in balance under high salt conditions by maintaining high water contents. Succulence is expressed as large cell size, reduced growth and surface area per tissue volume, and increased water constituents. Interestingly, halophytes also have a greater number of mitochondria, indicating that more energy is required to survive under saline conditions
[11][12]. Halophytes also have less sodium and chloride ion accumulation in their cytoplasms, allowing their chloroplasts to survive even while the plant experiences salinity shocks
[13][14]. Halophytes also have a specialized system for salt excretion from the plant tissues via specific glands. These glands are characteristic of halophytic leaves. The leaves will remove the salts onto the leaf surface before the salts can reach the shoots of the plant. The presence of halophytes is limited to habitats with plentiful water (e.g., salt marshes, etc.). “Salt hairs,” which regulate water loss, replace “secretary glands”, if a plant is adapted to a relatively drier climate as compared to marshes
[15][16]. Hydathodes are another adaptation by plants to remove excessive salts, with less stomatal conductance and transpiration water loss
[17][18].
Various undesirable effects appear because of high salt concentration. Ion imbalance is one of the major consequences. A high concentration of Na and Cl ions, as an example, can lead to biochemical processes which can prove to be fatal for the plants
[19][20][21]. Sodium and chloride toxicity not only induce nutritional disorders but also cause physiological drought by lowering the osmotic potential of the soil solutions
[22]. Soil salinity prevents the plant from taking up water from the soil, resulting in a decline in cellular water, thus affecting cell turgor. Soil salinity also adversely affects photosynthetic activity in the plant and encourages the production of reactive oxygen species (ROS), thus reducing plant growth
[23][24].
The identification of salt stress by plant species and their subsequent response is controlled by signals—signals which are generated by ions, osmotic differential, hormones, or ROS
[25]. These signals bind to their respective receptors and initiate the physiological mechanisms which enable a plant to adapt to stress conditions (). Under abiotic stress conditions, three types of signal transduction have been categorized, i.e., the ionic signaling pathway, the osmolyte regulation pathway, and the gene regulation pathway
[26]. For signal transduction under salinity stress, the ionic stress signaling pathway has been elucidated. Calcium (Ca) occupies a central position in this regard. It induces signal transduction in plants to adapt to stress conditions
[27]. High cytosolic Ca concentration initiates many processes involving enzymatic activity regulation, ion channel performance, and gene expression
[28]. Exogenously applied calcium regulates K
+/Na
+ selectivity, and thus confers salt adaptation by improving signal transduction. Glycinebetaine is reported to maintain signal transduction and ion homeostasis under salt-stressed conditions
[29][30].
Figure 1. Salt stress signals that bind to their respective receptors and initiate the physiological and molecular mechanisms to enable a plant to survive under stressed conditions.
Glycophytes and halophytes mediate serious effects of salinity at the cellular level by inducing changes in the plasma membrane and cytoplasm. As a tolerance mechanism, the plant alters the structure and composition of their plasma membrane, especially lipid and protein contents. The cell membrane is usually the foremost target of any stress
[31]. Salt stress also alters the cell cytoplasmic viscosity and composition
[32][33].
It is essential to understand each tolerance mechanism at the cellular level in order to understand each tolerance mechanism at the plant level. The protoplasmic features studied at the cellular level include plasma membrane permeability, cytoplasmic viscosity, cytoplasmic streaming, and cell solute potential. Cytoplasmic viscosity describes the water contents of the cytoplasm in conjunction with its inter-macromolecular interactions. Cell solute potential represents the solute contents of the cell. Cell membrane permeability significantly increases with the increase in salinity
[34][35]. The ability of the plasma membrane to repair, regenerate, and maintain its integrity stabilizes the cell structure and function under stress conditions. It is mainly dependent on the composition of the plasma membrane (i.e., mainly lipid contents). Saline conditions result in enhanced lipid peroxidation
[36].
The cell membrane stability technique is widely utilized to judge the behavior of various plant genotypes in response to salt stress
[37]. Thus, it can contribute to assessing the salt tolerance of plant genotypes. Cell membrane stability is also reported to correlate with potassium (K) ions, osmotic potential, osmotic adjustment, and relative water contents
[38][39]. The differing patterns of cell membrane permeability help in characterizing genotypes as tolerant or sensitive. In a saline environment, salt-sensitive genotypes show marked alterations, whereas salt-tolerant genotypes show minor changes. Salinity also alters the degree of saturation of membrane fatty acids and membrane fluidity
[40].
The salt-tolerant genotypes have high cytoplasmic viscosity due to augmentation in hydrophilic cytoplasmic proteins and other macromolecules
[41]. In more sensitive genotypes, a saline environment results in a high concentration of salts in plant cells, lowering the solute potential
[42]. Salinity inflicts serious irregularities during cell division, one of the various metabolic processes which face severe alterations in a saline environment. A saline environment especially alters the leaf anatomy by affecting mitochondria and vacuoles
[43][44], plant leaf area, and stomatal thickness
[45]. One way plants exhibit tolerance to saline environments is portioning or compartmentalization of toxic ions. This mechanism enables salt-tolerant plant species to retain toxic levels of harmful ions in vacuoles and inhibit their interference with cytoplasmic metabolic activities
[46]. The Na
+ and Cl
− partitioning in the vacuole stimulates higher concentrations of K
+ and organic osmolytes in the cytoplasm in order to adjust osmotic pressure of the ions in the vacuole
[47].
1.2. Salinity and Morphological Attributes
Halophytes have the unique feature of succulence, a feature which keeps the ionic uptake in proper proportion with water, by maintaining high water contents (). Succulence results in large cell size, reduced growth
[48][49], reduced surface area per tissue volume, and increased water constituents. The salt tolerance is evident as maintenance of vegetative growth and yield and lower necrotic percentage
[50][51]. Moreover, halophytes also have a greater number of mitochondria, indicating that more energy is required to survive under saline conditions
[11][21]. Sodium (Na) accumulation causes necrosis in old leaves, initiating from tips and then extending towards the leaf base. It also decreases leaf life span, net productivity, and crop yield
[52]. Biomass reduction and foliar damage become more prominent with time and at higher salinity levels. High salt concentration caused a reduction in fresh fruit yield in various vegetables
[31][53][54][55]. However, salinity treatments did not prove harmful for vegetative growth and the number of flowers
[53][55]. In contrast to the above report, Chartzoulakis and Klapaki,
[56], Kaya et al.
[57], and Giuffrida et al.
[58], noted a reduction in fruit numbers and fruit weight under salt stress. Saline conditions were reported as producing non-significant results on some growth attributes, water status, and tissue concentration of major nutrients
[59]. Salt treatments caused a significant reduction in plant height, root length, and dry weight
[60][61][62]. Salt stress caused reduction in fresh and dry weight of cotton seedlings
[63][64] and seed germination percentage in wheat varieties
[35][65]. Tomato (
Solanum lycopersicum) and pepper plants grown under salt stress experienced a reduction in dry weight, plant height
[57][66], fruit weight, and relative water contents
[67]. Broad bean, which is a green vegetable, experienced a significant reduction in plant height, leaf area, pod weight, number of pods per plant, seed yield, number of seeds, and product quality due to salt stress. However, a significant positive trend was observed between dry leaf matter, specific leaf weight, and salinity
[68]. Suppression in seedling growth and dry matter accumulation was observed in Indian mustard (
Brassica juncea L.) because of salt stress, which was ameliorated by putrescine application
[69]. Exogenously applied sugar beet extract and shikimic acid on salt stressed eggplant and tomato crops respectively displayed a marked influence on fresh fruit weight, number of fruits, and shoot and root fresh and dry weight
[54][70].
Figure 2. Regulation of physiological and biochemical process in halophytes through ionic compartmentalization, osmotic adjustment, enzymatic activities, polyamines, and stress signaling regulation.
According to Caines and Shennan
[71], root growth is more susceptible to saline conditions than shoot growth, but both are affected, making them reasonable indicators of salinity damage. A similar suppression in the shoot and root growth was also observed by Evers et al.
[72] and Gao et al.
[73] in the case of
Solanum tuberosum L. Under high salt concentrations, potato root and shoot development was hindered
[74][75]. High salt concentrations reduced the leaf area and increased the root:shoot ratio in wild type (Ailsa Craig) and ABA-deficit mutant (
notabilis) tomato genotypes
[76]. Suppression of fresh and dry weight of tomato plants because of salt stress can be alleviated by promoting the growth of
Achromobacter piechaudii in the tomato growth media.
A. piechaudii also caused a reduction in ethylene production by tomato seedlings, the opposite effect of salt stress on tomato seedlings
[77]. High ethylene levels proved to be harmful to the growth of the plant
[78]. Interestingly, Hu et al.
[79] reported that potato root growth could be improved through brassinosteroid application. Salt stress caused a reduction in marketable yield of pepper plants grown hydroponically. Saline conditions also imparted negative features to the fruit quality in terms of fruit pulp thickness and firmness. It also resulted in increased fructose, glucose, and
myo-inositol fruit concentrations
[4][42]. The salt-sensitive pepper genotypes showed maximum damage and experienced severe chlorosis and necrosis, whereas tolerant genotypes were slightly less affected. Sodium exclusion can be regarded as a criterion to allocate salt stress tolerance status to pepper genotypes
[80]. Korkmaz et al.
[81] have reported that the exogenous application of glycine betaine can reduce the effect of salinity in pepper plants.
High salt concentration has been observed to cause detrimental effects on leguminous crops. In fact, saline conditions induced smaller sized nodules, reduced nodule volume per plant, less nodulation, and inferior plant growth. At the cellular level, saline conditions caused drastic alteration in the mechanism of nodule formation. It reduced the turgor of the peripheral cells of the nodule, altered its zonation, enhanced the infection thread enlargement, reduced the release of bacteria from infection threads, and stimulated the electron-dense material (phenolics) and its accumulation in vacuoles. Salinity caused a reduction in nitrogen-fixing ability of the nodules, which has the outcome of reduced respiration rate and protein synthesis
[82][83][84]. Salinity tolerance patterns vary considerably among leguminous crops. It is distinctly apparent in
Vicia faba,
Glycine max,
Pisum sativum, and
Casuarina glauca, which can be categorized as salt-sensitive
[85][86]. In addition, germination of seeds faces serious limitations upon exposure to salt stress
[87][88]. Halophytes and glycophytes differ significantly in their germination behavior. Halophytes have the ability to maintain their germination mechanism to an extent with the advent of salinity. However, a sharp decline in germination occurs for glycophytes under salt stress. Imbibition is affected by the lower solute potential of the soil solution. It results in enzymatic deregulations and imbalances in source-sink relationships and ratios of different plant growth regulators present in the seed and required for efficient seed germination
[89][90].
1.3. Salinity and Physiological Attributes
Photosynthesis is of prime importance in the production of adenosine triphosphate (ATP), which provides the energy required for CO
2 fixation to sugars. Various abiotic stresses alter photosynthetic mechanisms
[91] by disrupting thylakoid membranes, modifying the electron transport chain, altering enzymatic activity and protein synthesis, and changing Calvin cycle patterns. All these abnormalities cause deregulation in ATP synthesis
[92], which can lead to the deficiency of certain ions due to ion degradation and synthesis inhibition
[93][94][95][96].
Stepien and Klobus
[97] reported a decline in the relative water content of cucumber (
Cucumis sativus) leaves after exposure to saline solutions. They attributed their results to higher Na and reduced K content, a situation which reduces photosynthesis because of the antagonistic competition of Na for ion uptake. Gas exchange is inversely related to the concentration of Na and chloride ions
[98][99], and parameters like photosynthesis, stomatal conductance, and transpiration tend to be negatively influenced by saline treatments
[100][101]. Photosynthesis of pepper plants has been reported to be lower when the entire root system is affected by salt stress compared to partial root exposure, and factors like stomatal conductance and transpiration are similarly affected by complete or partial salt stress
[99][102]. A low stomatal conductance was reported in wild type (Ailsa Craig) and ABA-deficit mutant (
notabilis) tomato genotypes, which was negatively correlated with increasing xylem ABA for both genotypes
[76]. Stomatal conductance is often correlated with photosynthetic efficiency, which is a prerequisite for higher biomass production and yield
[10]. In addition, salinity is reported to reduce the maximum quantum efficiency of photosystem II (PSII)
[48][94][103][104][105][106]. A distinct correlation has been found between Na ion contents and chlorophyll fluorescence, which is often used to estimate salt tolerance of plants
[48]. Higher salt levels are also known to alter photosynthesis via non-stomatal limitations, including variations in photosynthetic enzyme activity and changes in the concentration of chlorophyll and carotenoids
[107][108]. Pepper leaves have exhibited a significant reduction in chlorophyll pigment under saline conditions
[109]. A similar reduction in chlorophyll
a and
b contents has been reported in melon by Kaya et al.
[57]. This chlorophyll degradation under salt stress can be attributed to an enzyme called chlorophyllase
[110][111][112][113] and to the absolute concentration of chloride and Na in the leaves
[98][114][115][116][117][118]. Although carotenoid content has been reported to decline in response to salinity, anthocyanin pigments typically increase as a result of salinity.
Plant antioxidants include secondary metabolites (phenolic compounds), which are generated in response to stress conditions. These secondary metabolites may include tocopherol, which serves to stabilize membrane integrity
[119][120], ascorbic acid, carotenoids, flavonoids, and glutathione
[90][121]. Tocopherol plays a key role as a signaling molecule between cells
[93]. Ascorbic acid is a significant antioxidant involved in plant adaptation
[122] and occurs abundantly in cell organelles and apoplasts
[99]. It has the ability to scavenge superoxide, hydroxyl, and singlet oxygen. Carotenoids are found in chloroplasts and reported to aid in light reception for photosynthesis. Moreover, they are also protective compounds which scavenge ROS
[95][123]. Putrescine is reported to increase the level of carotenoids and glutathione in Indian mustard (
B. juncea L.) against salt stress. Putrescine supplementation inhibits ROS generation by accelerating antioxygenic enzymes and therefore aiding in the maintenance of chloroplastic membranes and the NADP
+/NADPH ratio
[124][125][126].
C
4 plants are reported to be more resistant against salinity stress than C
3 plants by having a better capacity to preserve the photosynthetic apparatus against oxidative stress
[127][128]. Other physiological responses that are indirectly related to salinity stress include changes in water use efficiency and evapotranspiration, which can benefit from the use of beneficial bacteria
[77][129][130][131]. In addition, higher salt concentration can stimulate the accumulation of spermine and spermidine, which contribute to the induction of salt tolerance in plants and lead to maintenance of fruit quality ()
[132][133][134][135].
Figure 3. Bunches, grapevine cultivar Shiraz. Control bunch (
A); Salt-stressed bunch (
B); showing symptoms of coulure and millerandage. Salt treatment was applied from budburst until veraison via fertigation with 35 mM NaCl added to control nutrient solution
[136].
2. Salinity and Biochemical Attributes
Plants have the ability to sustain their life under a saline environment through synthesis and accumulation of compatible solutes in the cytosol. These are soluble compounds with low molecular mass. These chemical compounds can maintain physiological and biochemical processes, without having interference in these processes. The chief components of compatible solutes are sugar alcohols (mannitol, sorbitol, ononitol), quaternary ammonia compounds (glycine betaine, proline betaine), proline, and tertiary sulfonium compounds. They act to scavenge the reactive oxygen species (ROS) and inhibit lipid peroxidation, hence preventing damage at the cellular level. These compatible solutes act in favor of osmotic adjustment and prevent ROS damage at the cellular level
[137][138]. These maintain macromolecular conformation in the cytosol, which may be changed due to the accumulation of charged ions, under saline conditions
[8][97]. These organic compounds are termed as compatible due to their consistency with the cell’s metabolism
[139], and their lowering of the water potential without altering cell water contents. These organic compounds are hydrophilic in nature, having the ability to replace water present on protein surfaces
[140][141], without interfering with their structure and function. These solutes play a key role in preventing the drastic effects of high ion concentration on enzymatic activities
[24][142][143][144]. Such important roles of compatible solutes lead to osmoregulation of plant cells under osmotic stress. In addition to osmoregulation, these organic compounds have a distinct role in protein stabilization, maintenance of membrane integrity, protection of OEC of PSII from dissociation
[145], and scavenging of reactive oxygen species (ROS). Mannitol, sorbitol, glycerol, proline, ononitol, and pinitol have been reported to scavenge ROS species
[146].
Mannitol (sugar alcohol) metabolism in higher plants is a superior attribute, contributing to salt and osmotic stress tolerance while playing a significant role as a compatible solute. It also improves plant responses under biotic stress as well, like under pathogen infestation
[147]. Mannitol is reported to be synthesized at the same time with either sucrose or raffinose saccharide. In salt-tolerant species, mannitol accumulation increases, indicating that high mannitol levels contribute to salt tolerance. Mannitol acts to scavenge the reactive oxygen species, thus protecting protein molecules
[148][149]. Pinitol and ononitol have been reported to accumulate under various stresses, predominantly drought and salt stress
[150][151]. Interestingly, polyols can be used as a potential biochemical marker for genetically engineered stress resistance plant genotypes
[152][153].
2.1. Salinity and Proline
Proline is an osmolyte, an amino acid, which is thought to play a significant role in inducing tolerance in plants against stressed conditions
[154]. Salt stress can result in elevation in proline levels
[74]. Ethephon, when used with sodium chloride in spinach, also increased proline levels
[155]. The importance of proline is highlighted by its existence in bacteria with a relationship to plants experiencing water or salinity stress. High proline levels can serve as a nitrogen source for plants during recovery
[156]. The precursor of proline synthesis is glutamate, involving pyrroline carboxylic acid synthetase and pyrroline carboxylic reductase
[157]. An increase was noted in the activity of pyrroline-5-carboxylate synthetase (P5CS) and a decline was recorded in proline dehydrogenase activity in potato seedlings under salt stress. These changes of enzymatic activity were more pronounced in salt-sensitive cultivars
[74][158]. However, an increase in proline contents of potato clones was recorded upon salt exposure
[50][159]. It serves to stabilize ultra-structural changes in cells, scavenge ROS (reactive oxygen species), and maintain cellular redox potential. Under stress conditions, a higher accumulation of proline is reported in cell cytosol, strengthening the ability of the cell to make ionic adjustments. Its accumulation is linearly related to stress tolerance in plants
[160]. Proline biosynthesis is reported to be mediated by Ca
[161][162][163] and abscisic acid
[8]. Previously, contrasting views about proline accumulation were also reported in plants under stress
[19][149], where it appeared as a salt stress injury symptom, e.g., rice
[164] and sorghum
[165].
Some plant genotypes do not respond to proline accumulation, but their salt tolerance potential can be enhanced through the exogenous application of proline
[31][166]. It may be helpful in counteracting the harmful effects through osmo-protection, resulting in a higher growth rate. Proline also increases the activities of antioxidant enzymes like SOD (superoxide dismutase) and POD (peroxidase)
[167]. Proline is not reported to scavenge ROS directly, but through enhanced antioxidant enzyme activity. It is reported to be more effective in mitigating the drastic effects of salinity than glycine betaine
[168]. Proline used at higher concentrations may prove to be lethal for the plant, causing ultra-structural damages leading to ROS generation
[169]. The effective dose of proline varies with genotype and plant developmental stage
[170][171][172][173]. Proline accumulation has been reported for drought sensitive and tolerant barley genotypes grown under saline conditions. Under salt stress, a considerable amount of proline was present, with relatively lower quantities in root tissues. Proline accumulation is reported to be more prominent in tolerant genotypes
[174].
2.2. Salinity and Polyamines
Polyamines are multivalent compounds consisting of two or more amino groups. In higher plants, these are identified as putrescine, spermidine, and spermine
[138][175]. These are involved in various physiological mechanisms including rhizogenesis, somatic embryogenesis, maintenance of cell pH and ionic balance
[29], pollen and flower formation, abscission, senescence, and dormancy. Endogenous polyamine synthesis can be stimulated by cytokinin
[176][177]. These compounds act to stabilize macromolecules like DNA and RNA. Moreover, polyamines have a significant role in numerous abiotic and biotic stresses
[142][178]. At the cellular level, polyamines contribute to regulating the plasma membrane potential, ionic homeostasis, and tolerance against salinity
[179][180]. Exogenously applied polyamine or ornithine caused a reduction in proline accumulation in plant tissues under salt stress. However, an alternate trend was observed in the case of non-stressed beans
[99][181]. Putrescine is characterized as a de-stressor agent and a nitrogen source under stressed conditions
[182]. Putrescine has been reported to reverse the biomass reduction in Indian mustard
[68][183]. Its production in plant cells follows two alternative pathways: conversion from ornithine or arginine. Putrescine is then converted to spermidine and subsequently to spermine by addition of an aminopropyl group. Spermine deficiency caused Ca ion imbalance in
Arabidopsis thaliana, thus indicating spermine as a maintainer of plant cell ionic homeostasis under salt stress
[184].
2.3. Salinity and Glycine-Betaine
Glycine-betaine (GB) is present in a wide range of organisms, from bacteria to higher plants and animals. In addition to being involved in osmoregulation, it maintains and regulates the performance of PSII protein complexes by protecting extrinsic regulatory protein against denaturation. It also stabilizes macromolecules, due to its ability to form strong bonds with water
[136]. It protects these macromolecules during drought and thermal stress, which is why it is sometimes called an “osmoprotectant”
[129][185]. Glycine-betaine accumulates in some crops under stress, like members of family
Poaceae and
Chenopodiaceae [186], and is absent entirely from other plants, like rice and tobacco. This directed the scientists to develop transgenic plants that have the ability to produce GB. In transgenic plants, the reproductive organs are capable of tolerating abiotic stresses if they can accumulate GB
[186]. The precursor for GB is choline, and the conversion is managed by enzymes like choline monooxygenase and betaine-aldehyde dehydrogenase
[142][187]. Choline supplementation to the growth media of the salt-stressed plant can act to restore the suppressed growth
[188]. GB is water soluble, is not harmful at higher concentrations, and accumulates mainly in plastids and chloroplasts. Exogenous application of GB promotes salinity tolerance in plant species which do not naturally produce GB. A plant can utilize exogenously applied GB via leaves
[189], as well as roots
[190]. After absorption, GB is translocated in phloem
[99][191]. GB is not directly involved in scavenging ROS species, but it alleviates the damaging effects of ROS by promoting enzymes responsible for the destruction or production suppression of ROS
[192].
The reproductive stage of any plant during GB application is considered critical to ensure maximum yield. In various studies, it was reported that the plant reproductive organs acquire higher levels of GB than the vegetative parts under stressed conditions. This indicates that high GB accumulation is more necessary for protecting the reproductive organs than it is for protecting the vegetative tissues from abiotic stresses, indicating that application timing is key
[172][193]. The natural GB accumulating species include sugar beets, spinach, wheat, barley, and sorghum. High GB concentration is linearly linked with increased tolerance. Osmotic adjustment is the major mechanism involved in increased tolerance to abiotic stresses, especially salt stress. GB is responsible for turgor maintenance through osmotic adjustment
[54][173][194]. However, this relationship is not satisfactory in some cases like
Triticum spp. and
Agropyron spp.
[195]. Thus, this relationship varies with genotype
[149]. The plant species which do not produce GB naturally can give a satisfactory yield and survival rate under salt stress conditions through the exogenous application of GB
[42][196]. Exogenous GB, once applied, is transported rapidly throughout the plant. Exogenous application of GB has been reported in many plant species, including tobacco, rice, soybean, barley, and wheat. In barley, GB application improved stress tolerance by lowering water potential, which improved survivability. GB plays a role in osmotic adjustment and ionic homeostasis by maintaining high K
+ concentration compared to Na
+ ions. Exogenous application of GB also increased the K
+/Na
+ ratio
[197][198][199]. GB also protects the photosynthetic apparatus. It enhances photosynthetic activity through increased stomatal conductance and reduced photorespiration
[42][200][201].
In contrast to its positive influence, some researchers have also suggested neutral or somewhat negative responses to exogenously applied GB in some plant genotypes. For example, it appeared to have a neutral influence on growth in cotton
[202], turnip, rapeseed, and tomato
[203]. For the commercial application of GB, the rate, duration, timing, and frequency should be considered
[149][204]. It can be used for seed treatment as well as foliar application. The application method is dependent on the plant material on which it will be applied, the timing of the application relative to plant developmental stage, and environmental conditions during the time of application
[205].
Exogenously applied GB improved salt tolerance in rice by improving relative water contents in the leaves and increasing antioxidant levels, including superoxide dismutase, ascorbate peroxidase, catalase, and glutathione reductase (GR)
[204]. Reduction in peroxidase activity was reported in a salt-tolerant rice genotype under salt stress. GB is also reported to reduce lipid peroxidation
[206][207][208]. GB can prevent membrane adulterations due to osmotic stress more efficiently than proline
[209]. Proline accumulation in leaves of salt-stressed plants is not reported to be correlated with exogenously applied glycine betaine
[54][171]. Sugar beet is identified as the foremost source of GB
[149][210]. It is appreciated as a valuable source of GB along with other beneficial compounds and is useful in inducing tolerance against salt stress in eggplant (
Solanum melongena L.) as compared to pure GB. It has a marked influence on the morphological (growth and yield) as well as physiological and biochemical (gas exchange, photosynthetic rate, transpiration, GB accumulation) attributes
[177][211].
3. Salinity and Phytohormones
Plant hormones are signaling molecules with the ability to alter the physiological mechanisms of the plant, even if present in very minute quantities
[212]. Common plant hormones are auxins, gibberellins, cytokinins, abscisic acid, and ethylene
[213]. Plants growing under salt stress experience imbalances in hormonal homeostasis. Stressed conditions drastically alter physiological mechanisms of the plant, creating massive changes in endogenous hormonal contents. Higher concentrations of toxic ions are negatively correlated with the levels of plant hormones like gibberellins, auxins, and cytokinin and are positively associated with the abscisic acid level
[214][215]. Exogenous plant hormone application on salt stressed plants was found to alleviate the negative effects of salinity on the morphological (leaf area, dry mass), physiological (chlorophyll content, stomatal conductance, photosynthetic rate), and yield characteristics of crops
[216][217].
Abscisic acid (ABA) and ethylene are involved in signaling under stress conditions. An increase in ABA concentration in plant cells has been reported under saline conditions
[218]. Carotenoids are the precursor for ABA synthesis, with roots and leaves the sites of synthesis
[219]. Water deficit in the root zone causes ABA generation in roots, and ABA transport to shoots is xylem mediated. The increase in pH of xylem sap increases the transport of ABA to the guard cells, where it regulates the stomatal opening and closing through the involvement of Ca ions
[220]. Alterations in ABA levels indirectly affect photosynthesis through disruption in stomatal opening and closing. The photosynthetic efficiency of the plant cell declines along with deregulation of translocation and assimilate partitioning of photosynthates
[8][221].
Exogenously applied plant growth regulators have been widely reported to enhance stress tolerance in numerous plant species
[222]. ABA is reported to be useful in alleviating plant salt stress under low water potential. ABA production in the plant cell is also related to ethylene synthesis under salt stress. The interaction between these two stress hormones is apparent from vegetative growth and seed germination under salt stress. During root inhibition by salt stress, ethylene regulates the ABA concentration. However, the reverse has been reported in the case of seed germination
[223]. Salicylic acid has displayed a defensive role in plants experiencing stress, signaling the plant to adapt to the stressful environment
[224].
Salinity and Growth Regulation
Brassinosteroids (BRs) are growth regulators which mitigate adverse growth patterns caused by salinity. It improves the germination of seeds experiencing salt stress. The improved germination rate has been reported for rice
[225] and tobacco. Application of BRs as a seed treatment enhanced the growth of rice seedlings under salt stress
[226]. It helps the plant to retain its green pigments and enhances nitrate reductase activity
[227][228][229] and nitrogen-fixing capability. Brassinosteroids (28-homoBL) increased dry matter accumulation and seed yield
[230]. Foliar application of brassinosteroids (24-epibrassinolide) on pepper plants grown with saline water greatly affected shoot growth parameters and leaf water contents as compared to roots. However, its effect on chlorophyll fluorescence was non-significant
[231][232]. Similar patterns of brassinosteroidal effects were observed in wheat grown under salt stress. 24-epibrassinolide application on salt-stressed wheat seedlings exhibited non-significant results in terms of plant biomass, chlorophyll content, photosynthesis rate, substomatal CO
2 concentration, and water use efficiency. The incremented water use efficiency can be related to higher transpiration rate shown by salt-stressed wheat seedlings, as a result of 24-epibrassinolide application
[233][234]. The efficiency of exogenously applied brassinosteroids to mitigate salinity effects varies with plant species, appropriate growth stage, dose, frequency, and method of brassinosteroidal application
[235][236][237]. The results of BR application also vary with climatic conditions—mainly temperature, light duration, and applied fertilizers
[238]. Brassinolide application to salt stressed
Vigna radiata caused enhancement in growth, photosynthetic rate, and maximum quantum yield of PSII. Generally, brassinolide has the potential to protect the photosynthetic apparatus under salt stress. It also contributed to improving the membrane stability index and leaf water potential. However, no significant results were recorded in the case of lipid peroxidation and electrolyte leakage. Brassinolide increases antioxidant enzyme and proline contents
[239][240][241][242]. Increases in proline level create a protective shield when the plant is under stress by acting as a source of carbon and nitrogen, a stabilizer of the plasma membrane, and an oxygen radical scavenger
[243]. Brassinosteroids also increase pigment levels in the plant
[244]. BRs also improve the nitrate and nitrite reductase activity in
Vigna radiata under salt stress. This effect can be attributed to the ability of BRs to modulate transcription and translation at the gene level, and to increase cell nitrate uptake. Increased stress tolerance caused by brassinolide application is observable as improved growth parameters such as shoot length, root length, and plant biomass
[106][245].
 +1 point
+1 point