 +1 credit
+1 credit
Video Upload Options
Muscarinic acetylcholine receptors (mAChRs) belong to the superfamily of G-protein-coupled receptors (GPCRs). The family of mAChRs is composed of five subtypes, M1, M2, M3, M4 and M5, which have distinct expression patterns and functions. In the eye and its adnexa, mAChRs are widely expressed and exert multiple functions, such as modulation of tear secretion, regulation of pupil size, modulation of intraocular pressure, participation in cell-to-cell signaling and modula-tion of vascular diameter in the retina. Due to this variety of functions, it is reasonable to assume that abnormalities in mAChR signaling may contribute to the development of various ocular diseases. On the other hand, mAChRs may offer an attractive therapeutic target to treat ocular diseases. Thus far, non-subtype-selective mAChR ligands have been used in ophthalmology to treat dry eye disease, myopia and glaucoma. However, these drugs were shown to cause various side-effects. Thus, the use of subtype-selective ligands would be useful to circumvent this problem.
1. Introduction
2. Expression and Distribution of mAChRs in the Retina
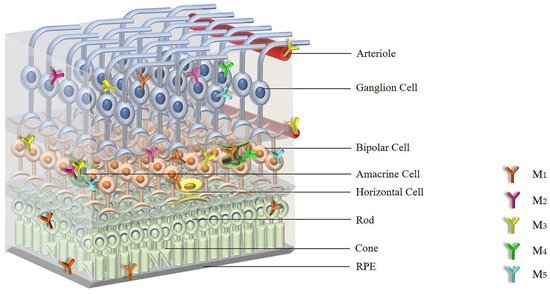
3. Cellular Signaling of mAChRs
4. Functional Roles of Individual mAChR Subtypes in the Retina
References
- Dale, H.H. The Action of Certain Esters and Ethers of Choline, and Their Relation to Muscarine. J. Pharmacol. Exp. Ther. 1914, 6, 147.
- McCorry, L.K. Physiology of the Autonomic Nervous System. Am. J. Pharm Educ. 2007, 71, 78.
- Caulfield, M.P.; Birdsall, N.J.M. International Union of Pharmacology. XVII. Classification of Muscarinic Acetylcholine Receptors. Pharmacol. Rev. 1998, 50, 279.
- Wessler, I.; Kirkpatrick, C.J. Acetylcholine Beyond Neurons: The Non-neuronal Cholinergic System in Humans. Br. J. Pharmacol. 2008, 154, 1558–1571.
- Liu, S.; Li, J. Expression and Function of Muscarinic Receptor Subtypes on Human Cornea and Conjunctiva. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2987–2996.
- Ríos, J.D.; Forde, K. Development of Conjunctival Goblet Cells and Their Neuroreceptor Subtype Expression. Investig. Ophthalmol. Vis. Sci. 2000, 41, 2127–2137.
- Liu, Q.; Wu, J. Changes in Muscarinic Acetylcholine Receptor Expression in Form Deprivation Myopia in Guinea Pigs. Mol. Vis. 2007, 13, 1234–1244.
- Mitchelson, F. Muscarinic Receptor Agonists and Antagonists: Effects on Ocular Function. In Muscarinic Receptors; Fryer, A.D., Christopoulos, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 263–298.
- Sloniecka, M. Neuropeptides and Neurotransmitters in Keratocytes: Importance in Corneal Wound Healing Processes. Ph.D. Thesis, Comprehensive Summary, Umeå University, Umeå, Sweden, 2015.
- Smith, E.L., III; Redburn, D.A. Permanent Alterations in Muscarinic Receptors and Pupil Size Produced by Chronic Atropinization in Kittens. Investig. Ophthalmol. Vis. Sci. 1984, 25, 239–243.
- McBrien, N.A.; Jobling, A.I. Expression of Muscarinic Receptor Subtypes in Tree Shrew Ocular Tissues and Their Regulation during the Development of Myopia. Mol. Vis. 2009, 15, 464–475.
- Ríos, J.D.; Zoukhri, D. Immunolocalization of Muscarinic and Vip Receptor Subtypes and Their Role in Stimulating Goblet Cell Secretion. Investig. Ophthalmol. Vis. Sci 1999, 40, 1102–1111.
- Duncan, G.; Collison, D.J. Role of the Non-neuronal Cholinergic System in the Eye: A Review. Life Sci. 2003, 72, 2013–2019.
- Lin, H.-J.; Wan, L. Muscarinic Acetylcholine Receptor 3 is Dominant in Myopia Progression. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6519–6525.
- Almasieh, M.; Zhou, Y. Structural and Functional Neuroprotection in Glaucoma: Role of Galantamine-mediated Activation of Muscarinic Acetylcholine Receptors. Cell Death Dis. 2010, 1, e27.
- Weinreb, R.N.; Aung, T. The Pathophysiology and Treatment of Glaucoma: A Review. JAMA 2014, 311, 1901–1911.
- Quigley, H.A.; Broman, A.T. The Number of People with Glaucoma Worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267.
- Tham, Y.-C.; Li, X. Global Prevalence of Glaucoma and Projections of Glaucoma Burden Through 2040: A Systematic Review and Meta-analysis. Ophthalmology 2014, 121, 2081–2090.
- Quigley, H.A.; Addicks, E.M. Optic Nerve Damage in Human Glaucoma. II. The Site of Injury and Susceptibility to Damage. Arch. Ophthalmol. 1981, 99, 635–649.
- Weinreb, R.N.; Khaw, P.T. Primary Open-angle Glaucoma. Lancet 2004, 363, 1711–1720.
- Heijl, A.; Leske, M.C. Reduction of Intraocular Pressure and Glaucoma Progression: Results from the Early Manifest Glaucoma trial. Arch. Ophthalmol. 2002, 120, 1268–1279.
- Nucci, C.; Martucci, A. Neuroprotective Agents in the Management of Glaucoma. Eye 2018, 32, 938–945.
- Grossniklaus, H.E.; Green, W.R. Pathologic Findings in Pathologic Myopia. Retina 1992, 12, 127–133.
- Morgan, I.; Rose, K. How Genetic is School Myopia? Prog. Retin. Eye Res. 2005, 24, 1–38.
- Holden, B.A.; Fricke, T.R. Global Prevalence of Myopia and High Myopia and Temporal Trends from 2000 Through 2050. Ophthalmology 2016, 123, 1036–1042.
- Schultz, G.; Rosenthal, W. Role of G Proteins in Calcium Channel Modulation. Annu. Rev. Physiol. 1990, 52, 275–292.
- Simon, M.I.; Strathmann, M.P. Diversity of G Proteins in Signal Transduction. Science 1991, 252, 802–808.
- Dreyer, F. Acetylcholine Receptor. Br. J. Anaesth. 1982, 54, 115–130.
- Kostenis, E.; Zeng, F.-Y. Structure-function Analysis of Muscarinic Receptors and Their Associated G Proteins. Life Sci. 1999, 64, 355–362.
- Laspas, P.; Zhutdieva, M.B. The M(1) Muscarinic Acetylcholine Receptor Subtype is Important for Retinal Neuron Survival in Aging Mice. Sci. Rep. 2019, 9, 5222.
- Yu, P.; Dong, W.-P. Huperzine a Lowers Intraocular Pressure via the M3 mAChR and Provides Retinal Neuroprotection via the M1 mAChR: A Promising Agent for the Treatment of Glaucoma. Ann. Transl. Med. 2021, 9, 332.
- Broadley, K.J.; Kelly, D.R. Muscarinic Receptor Agonists and Antagonists. Molecules 2001, 6, 142–193.
- Barathi, V.A.; Beuerman, R.W. Molecular Mechanisms of Muscarinic Receptors in Mouse Scleral Fibroblasts: Prior to and after Induction of Experimental Myopia with Atropine Treatment. Mol. Vis. 2011, 17, 680–692.
- Marquis, R.E.; Whitson, J.T. Management of Glaucoma: Focus on Pharmacological Therapy. Drugs Aging 2005, 22, 1–21.
- Lusthaus, J.; Goldberg, I. Current Management of Glaucoma. Med. J. Aust. 2019, 210, 180–187.
- McBrien, N.A.; Stell, W.K. How does Atropine Exert Its Anti-myopia Effects? Ophthalmic Physiol. Opt. 2013, 33, 373–378.
- Hutchins, J.B. Review: Acetylcholine as a Neurotransmitter in the Vertebrate Retina. Exp. Eye Res. 1987, 45, 1–38.
- Hruska, R.E.; White, R. Muscarinic Cholinergic Receptors in Mammalian Retina. Brain Res. 1978, 148, 493–498.
- Atterwill, C.K.; Mahoney, A. The Uptake and Subcellular Distribution of 3-h-choline by the Retina. Br. J. Pharmacol. 1975, 53, 447P.
- Sugiyama, H.; Daniels, M.P. Muscarinic Acetylcholine Receptors of the Developing Retina. Proc. Natl. Acad. Sci. USA 1977, 74, 5524–5528.
- Polans, A.S.; Hutchins, J.B. Muscarinic Cholinergic Receptors in the Retina of the Larval Tiger Salamander. Brain Res. 1985, 340, 355–362.
- Hutchins, J.B.; Hollyfield, J.G. Acetylcholine Receptors in the Human Retina. Investig. Ophthalmol. Vis. Sci. 1985, 26, 1550–1557.
- Cho, N.J.; Klein, W.L. Muscarinic Acetylcholine Receptors from Avian Retina and Heart Undergo Different Patterns of Molecular Maturation. J. Neurochem. 1988, 50, 1403–1411.
- Hutchins, J.B. Development of Muscarinic Acetylcholine Receptors in the Ferret Retina. Dev. Brain Res. 1994, 82, 45–61.
- Townes-Anderson, E.; Vogt, B.A. Distribution of Muscarinic Acetylcholine Receptors on Processes of Isolated Retinal Cells. J. Comp. Neurol. 1989, 290, 369–383.
- Moroi-Fetters, S.E.; Neff, N.H. Muscarinic Receptor-mediated Phosphoinositide Hydrolysis in the Rat Retina. J. Pharmacol. Exp. Ther. 1988, 246, 553.
- Bonner, T.I. New Subtypes of Muscarinic Acetylcholine Receptors. Trends Pharmacol. Sci. 1989, (Suppl.), 11–15.
- Wall, S.J.; Yasuda, R.P. The Ontogeny of M1–M5 Muscarinic Receptor Subtypes in Rat Forebrain. Dev. Brain Res. 1992, 66, 181–185.
- Bonner, T.I. The Molecular Basis of Muscarinic Receptor Diversity. Trends Neurosci. 1989, 12, 148–151.
- McKinnon, L.A.; Rosoff, M. Regulation of Muscarinic Receptor Expression and Function in Cultured Cells and in Knock-out Mice. Life Sci. 1997, 60, 1101–1104.
- Fischer, A.J.; McKinnon, L.A. Identification and Localization of Muscarinic Acetylcholine Receptors in the Ocular Tissues of the Chick. J. Comp. Neurol. 1998, 392, 273–284.
- Belmonte, K.E.; McKinnon, L.A. Developmental Expression of Muscarinic Acetylcholine Receptors in Chick Retina: Selective Induction of M2 Muscarinic Receptor Expression in ovo by a Factor Secreted by Muller Glial Cells. J. Neurosci. 2000, 20, 8417–8425.
- Zhang, L.H.; Yan, D.S. Expression of Muscarinic Acetylcholine Receptor-1 in Human Retinal Pigment Epithelium. Zhonghua Yan Ke Za Zhi Chin. J. Ophthalmol. 2006, 42, 1109–1112.
- Tao, Y.; Li, X.-L. Effect of Green Flickering Light on Myopia Development and Expression of M1 Muscarinic Acetylcholine Receptor in Guinea Pigs. Int. J. Ophthalmol. 2018, 11, 1755–1760.
- Strang, C.E.; Renna, J.M. Muscarinic Acetylcholine Receptor Localization and Activation Effects on Ganglion Response Properties. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2778–2789.
- Gericke, A.; Sniatecki, J.J. Identification of the Muscarinic Acetylcholine Receptor Subtype Mediating Cholinergic Vasodilation in Murine Retinal Arterioles. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7479–7484.
- Gericke, A.; Mayer, V.G.A. Cholinergic Responses of Ophthalmic Arteries in M3 and M5 Muscarinic Acetylcholine Receptor Knockout Mice. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4822–4827.
- Peralta, E.G.; Ashkenazi, A. Distinct Primary Structures, Ligand-binding Properties and Tissue-specific Expression of Four Human Muscarinic Acetylcholine Receptors. EMBO J. 1987, 6, 3923–3929.
- Bonner, T.I.; Young, A.C. Cloning and Expression of the Human and Rat M5 Muscarinic Acetylcholine Receptor Genes. Neuron 1988, 1, 403–410.
- van Koppen, C.J.; Kaiser, B. Regulation of Muscarinic Acetylcholine Receptor Signaling. Pharmacol. Ther. 2003, 98, 197–220.
- Rümenapp, U.; Asmus, M. The M3 Muscarinic Acetylcholine Receptor Expressed in HEK-293 Cells Signals to Phospholipase D via G12 but not Gq-type G Proteins: Regulators of G Proteins as Tools to Dissect Pertussis Toxin-resistant G Proteins in Receptor-Effector Coupling. J. Biol. Chem. 2001, 276, 2474–2479.
- Mathes, C.; Thompson, S.H. The Nitric Oxide/Cgmp Pathway Couples Muscarinic Receptors to the Activation of Ca2+ Influx. J. Neurosci. 1996, 16, 1702–1709.
- Haga, T. Molecular Properties of Muscarinic Acetylcholine Receptors. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2013, 89, 226–256.
- Goin, J.C.; Nathanson, N.M. Subtype-specific Regulation of the Expression and Function of Muscarinic Acetylcholine Receptors in Embryonic Chicken Retinal Cells. J. Neurochem. 2002, 83, 964–972.
- Jardon, B.; Bonaventure, N. Possible Involvement of Cholinergic and Glycinergic Amacrine Cells in the Inhibition Exerted by the on Retinal Channel on the Off Retinal Channel. Eur. J. Pharmacol. 1992, 210, 201–207.
- Dowling, J.E. The Retina: An Approachable Part of the Brain; Harvard University Press: London, UK, 1987.
- Niemeyer, G.; Jurklies, B. Binding and Electrophysiology of the Muscarinic Antagonist QNB in the Mammalian Retina. Klin. Mon. Augenheilkd. 1995, 206, 380–383.
- Jositsch, G.; Papadakis, T. Suitability of Muscarinic Acetylcholine Receptor Antibodies for Immunohistochemistry Evaluated on Tissue Sections of Receptor Gene-deficient Mice. Naunyn Schmiedebergs Arch. Pharmacol. 2009, 379, 389–395.
- Hamilton, S.E.; Loose, M.D. Disruption of the M1 Receptor Gene Ablates Muscarinic Receptor-dependent M Current Regulation and Seizure Activity in Mice. Proc. Natl. Acad. Sci. USA 1997, 94, 13311.
- Gomeza, J.; Shannon, H. Pronounced Pharmacologic Deficits in M2 Muscarinic Acetylcholine Receptor Knockout Mice. Proc. Natl. Acad. Sci. USA 1999, 96, 1692–1697.
- Matsui, M.; Motomura, D. Multiple Functional Defects in Peripheral Autonomic Organs in Mice Lacking Muscarinic Acetylcholine Receptor Gene for the M3 Subtype. Proc. Natl. Acad. Sci. USA 2000, 97, 9579.
- Gomeza, J.; Zhang, L. Enhancement of d1 Dopamine Receptor-mediated Locomotor Stimulation in M4 Muscarinic Acetylcholine Receptor Knockout Mice. Proc. Natl. Acad. Sci. USA 1999, 96, 10483.
- Woolley, M.L.; Carter, H.J. Attenuation of Amphetamine-induced Activity by the Non-selective Muscarinic Receptor Agonist, Xanomeline, is Absent in Muscarinic M4 Receptor Knockout Mice and Attenuated in Muscarinic M1 Receptor Knockout Mice. Eur. J. Pharmacol. 2009, 603, 147–149.
- Yamada, M.; Lamping, K.G. Cholinergic Dilation of Cerebral Blood Vessels is Abolished in M(5) Muscarinic Acetylcholine Receptor Knockout Mice. Proc. Natl. Acad. Sci. USA 2001, 98, 14096.
- Steidl, S.; Yeomans, J.S. M5 Muscarinic Receptor Knockout Mice Show Reduced Morphine-induced Locomotion but Increased Locomotion after Cholinergic Antagonism in the Ventral Tegmental Area. J. Pharmacol. Exp. Ther. 2009, 328, 263.
- Barathi, V.A.; Kwan, J.L. Muscarinic Cholinergic Receptor (M2) Plays a Crucial Role in the Development of Myopia in Mice. Dis. Model. Mech. 2013, 6, 1146–1158.
- Gericke, A.; Steege, A. Role of the M3 Muscarinic Acetylcholine Receptor Subtype in Murine Ophthalmic Arteries after Endothelial Removal. Investig. Ophthalmol. Vis. Sci. 2014, 55, 625–631.
- Laspas, P.; Sniatecki, J.J. Effect of the M1 Muscarinic Acetylcholine Receptor on Retinal Neuron Number Studied with Gene-Targeted Mice. J. Mol. Neurosci. 2015, 56, 472–479.
- Lanzafame, A.A.; Christopoulos, A. Cellular Signaling Mechanisms for Muscarinic Acetylcholine Receptors. Recept. Channels 2003, 9, 241–260.
- Brown, D.A.; Abogadie, F.C. Muscarinic Mechanisms in Nerve Cells. Life Sci. 1997, 60, 1137–1144.
- Brown, D.A.; Selyanko, A.A. Membrane Currents Underlying the Cholinergic Slow Excitatory Post-synaptic Potential in the Rat Sympathetic Ganglion. J. Physiol. 1985, 365, 365–387.
- Wess, J.; Liu, J. Structural Basis of Receptor/G Protein Coupling Selectivity Studied with Muscarinic Receptors as Model Systems. Life Sci. 1997, 60, 1007–1014.
- Tan, P.-P.; Yuan, H.-H. Activation of Muscarinic Receptors Protects Against Retinal Neurons Damage and Optic Nerve Degeneration in Vitro and in Vivo Models. CNS Neurosci. Ther. 2014, 20, 227–236.
- Zhou, W.; Zhu, X. Neuroprotection of Muscarinic Receptor Agonist Pilocarpine Against Glutamate-induced Apoptosis in Retinal Neurons. Cell. Mol. Neurobiol. 2008, 28, 263–275.
- Pereira, S.P.F.; Medina, S.V. Cholinergic Activity Modulates the Survival of Retinal Ganglion Cells in Culture: The Role of M1 Muscarinic Receptors. Int. J. Dev. Neurosci. 2001, 19, 559–567.
- Zhu, X.; Zhou, W. Pilocarpine Protects Cobalt Chloride-induced Apoptosis of rgc-5 Cells: Involvement of Muscarinic Receptors and hif-1 Alpha Pathway. Cell. Mol. Neurobiol. 2010, 30, 427–435.
- Yu, P.; Zhou, W. L-satropane Prevents Retinal Neuron Damage by Attenuating Cell Apoptosis and aβ Production via Activation of M1 Muscarinic Acetylcholine Receptor. Curr. Eye Res. 2017, 42, 1319–1326.
- Granja, M.G.; Gomes Braga, L.E. Igf-1 and igf-1r Modulate the Effects of il-4 on Retinal Ganglion Cells Survival: The Involvement of M1 Muscarinic Receptor. Biochem. Biophys. Res. Commun. 2019, 519, 53–60.
- Braga, L.E.G.; Miranda, R.L. Pkc Delta Activation Increases Neonatal Rat Retinal Cells Survival in Vitro: Involvement of Neurotrophins and M1 Muscarinic Receptors. Biochem. Biophys. Res. Commun. 2018, 500, 917–923.
- dos Santos, A.A.; Medina, S.V. Protein Kinase C Regulates the Expression of M1 Receptors and BDNF in Rat Retinal Cells. Neurochem. Res. 2009, 34, 884–890.
- Braga, L.E.G.; Granja, M.G. Pma Increases M3 Muscarinic Receptor Levels and Decreases Retinal Cells Proliferation Through a Change in the Levels of Cell-cycle Regulatory Proteins. Neurosci. Lett. 2013, 550, 29–34.
- Borda, E.; Berra, A. Correlations between Neuronal Nitric Oxide Synthase and Muscarinic M3/M1 Receptors in the Rat Retina. Exp. Eye Res. 2005, 80, 391–399.
- Berra, A.; Ganzinelli, S. Inducible Nitric Oxide Synthase Subserves Cholinergic Vasodilation in Retina. Vis. Neurosci. 2005, 22, 371–377.
- Liu, J.; McGlinn, A.M. Nicotinic Acetylcholine Receptor Subunits in Rhesus Monkey Retina. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1408–1415.
- Strang, C.E.; Amthor, F.R. Rabbit Retinal Ganglion Cell Responses to Nicotine can be Mediated by Beta2-containing Nicotinic Acetylcholine Receptors. Vis. Neurosci. 2003, 20, 651–662.
- Strang, C.E.; Andison, M.E. Rabbit Retinal Ganglion Cells Express Functional Aalpha7 Nicotinic Acetylcholine Receptors. Am. J. Physiol. Cell Physiol. 2005, 289, C644–C655.
- Strang, C.E.; Long, Y. Nicotinic and Muscarinic Acetylcholine Receptors Shape Ganglion Cell Response Properties. J. Neurophysiol. 2015, 113, 203–217.
- Smith, M.L.; Souza, F.G.O. Acetylcholine Receptors in the Retinas of the α7 Nicotinic Acetylcholine Receptor Knockout Mouse. Mol. Vis. 2014, 20, 1328–1356.
- Schmidt, M.; Bienek, C. Differential Calcium Signalling by M2 and M3 Muscarinic Acetylcholine Receptors in a Single Cell Type. Naunyn Schmiedebergs Arch. Pharmacol. 1995, 352, 469–476.
- Pals-Rylaarsdam, R.; Hosey, M.M. Two Homologous Phosphorylation Domains Differentially Contribute to Desensitization and Internalization of the M2 Muscarinic Acetylcholine Receptor. J. Biol. Chem. 1997, 272, 14152–14158.
- Zang, W.J.; Yu, X.J. On the Role of G Protein Activation and Phosphorylation in Desensitization to Acetylcholine in Guinea-Pig Atrial Cells. J. Physiol. 1993, 464, 649–679.
- Antal, M.; Acuna-Goycolea, C. Cholinergic Activation of M2 Receptors Leads to Context-dependent Modulation of Feedforward Inhibition in the Visual Thalamus. PLoS Biol. 2010, 8, e1000348.
- Cimini, B.A.; Strang, C.E. Role of Acetylcholine in Nitric Oxide Production in the Salamander Retina. J. Comp. Neurol. 2008, 507, 1952–1963.
- Onali, P.; Olianas, M. Muscarinic M4 Receptor Inhibition of Dopamine d1-like Receptor Signalling in Rat Nucleus Accumbens. Eur. J. Pharmacol. 2002, 448, 105–111.
- Arumugam, B.; McBrien, N.A. Muscarinic Antagonist Control of Myopia: Evidence for M4 and M1 Receptor-based Pathways in the Inhibition of Experimentally-induced Axial Myopia in the Tree Shrew. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5827–5837.
- Cottriall, C.L.; Truong, H.-T. Inhibition of Myopia Development in Chicks Using Himbacine: A Role for M4 Receptors? Neuroreport 2001, 12, 2453–2456.
- Inagaki, Y.; Mashima, Y. Polymorphism of Beta-adrenergic Receptors and Susceptibility to Open-angle Glaucoma. Mol. Vis. 2006, 12, 673–680.
- Liao, Q.; Wang, D.-H. Association of Genetic Polymorphisms of Enos with Glaucoma. Mol. Vis. 2011, 17, 153–158.
- Michel, M.C.; Teitsma, C.A. Polymorphisms in Human Muscarinic Receptor Subtype Genes. Handb. Exp. Pharmacol. 2012, 208, 49–59.

