 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Himani Punia | + 2270 word(s) | 2270 | 2021-01-07 03:41:34 | | | |
| 2 | Dean Liu | -673 word(s) | 1597 | 2021-05-07 05:20:53 | | |
Video Upload Options
The primary objectives of modern agriculture includes the environmental sustainability, low production costs, improved plants’ resilience to various biotic and abiotic stresses, and high sowing seed value. Delayed and inconsistent field emergence poses a significant threat in the production of agri-crop, especially during drought and adverse weather conditions. To open new routes of nutrients’ acquisition and revolutionizing the adapted solutions, stewardship planswill be needed to address these questions.One approach is the identification of plant based bioactive molecules capable of altering plant metabolism pathways which may enhance plant performance in a brief period of time and in a cost-effective manner.A biostimulant is a plant material, microorganism, or any other organic compound that not only improves the nutritional aspects, vitality, general health but also enhances the seed quality
performance.
1. Introduction
The primary objectives of modern agriculture includes the environmental sustainability, low production costs, improved plants’ resilience to various biotic and abiotic stresses, and high sowing seed value. Delayed and inconsistent field emergence poses a significant threat in the production of agri-crop, especially during drought and adverse weather conditions. To open new routes of nutrients’ acquisition and revolutionizing the adapted solutions, stewardship plans will be needed to address these questions. One approach is the identification of plant based bioactive molecules capable of altering plant metabolism pathways which may enhance plant performance in a brief period of time and in a cost-effective manner. A biostimulant is a plant material, microorganism, or any other organic compound that not only improves the nutritional aspects, vitality, general health but also enhances the seed quality performance. They may be effectively utilized in both horticultural and cereal crops. The biologically active substances in biostimulant biopreparations are protein hydrolysates (PHs), seaweed extracts, fulvic acids, humic acids, nitrogenous compounds, beneficial bacterial, and fungal agents. In this review, the state of the art and future prospects for biostimulant seedlings are reported and discussed. Biostimulants have been gaining interest as they stimulate crop physiology and biochemistry such as the ratio of leaf photosynthetic pigments (carotenoids and chlorophyll), enhanced antioxidant potential, tremendous root growth, improved nutrient use efficiency (NUE), and reduced fertilizers consumption. Thus, all these properties make the biostimulants fit for internal market operations. Furthermore, a special consideration has been given to the application of biostimulants in intensive agricultural systems that minimize the fertilizers’ usage without affecting quality and yield along with the limits imposed by European Union (EU) regulations.
2. Biostimulant Applications for Crop Agronomy
The role of bioactive compounds in signaling of primary and secondary metabolic pathways has been generally correlated with promoting germination, plant growth and crop productivity under the influence of PBs [1]. Hydrolyzed collagen of various forms, such as gelatin mixtures of hydrolysates and amino acids, and granulated gelatin inducing the formulation of gelatine was evaluated for cucumber growth [2]. Gelatin hydrolysates regulated the expression of permeases encoding genes (AAP3 and AAP6) and nitrogen and amino acids transporters. Hence, the authors inferred that gelatin hydrolysate may be used as a reliable nitrogen (N) source. Moreover, Luziatelli et al. [3] performed a greenhouse experiment on lettuce to assess the impact on three commercially available PBs: vegetal-derived protein hydrolysate (PH), vegetal-derived tropical plant extract, and Cu supplemented PH and epiphytic bacterial colony. Results showed that PBs enhanced the fresh weight of shoot with no significant variations among the organic PBs. They also showed that PBs may boost epiphytic bacterial growth (Acinetobacter, Bacillus, and Pseudomonas) with the aid of PGP and biological pathogens control activity, thus working in synergy with PB organic compounds to improve the fresh lettuce production for marketing. Moreover, Mahnert et al. [4] demonstrated the biostimulant ability comprising of stone dust, malt sprouts, and organic herbs to assess the beneficial effect on growth, development, and efficiency by the field application of microbiota and also in the surrounding. Furthermore, Lucini et al. [5] conducted study to evaluate the metabolomics and physiological responses in melon via the application of lateral root promoting peptides, lingo-sulphonates and micronutrients, a combination of biopolymer-based biostimulants. Different doses (0, 0.3, 0.6, 1.2, or 2.4 L ha−1) of vegetal-based biostimulants were applied around the collar tissue. The substrate drench elicits dose-dependent biomass accumulation in melon transplants. The root characteristics in biopolymer-treated plants were significantly higher at 0.24 mL as compared to 0.06 mL per plant. The enhanced accumulation of shoot and root biomass might be due to involvement of direct and indirect physiological processes in biopolymer-treated melon transplants. For example, signaling molecules specifically, the bioactive peptides and lignosulfonates may trigger signal transduction pathways by induction of target endogenous phytohormones [6]. Palumbo et al. [7] observed that humic acid extracted from municipal solid waste and olive mill water filters might be utilized as reliable biostimulants in dose-dependent manner in maize to enhance the plant growth, marker enzyme activities and nutrient mobilization. Ertani et al. [6] studied the effect of seaweed-based extracts, viz., from Laminaria and A. nodosum as potential biostimulants with a concentration of 0.5 mL L−1 in maize. By implication of different biochemical and morphological approaches, A. nodosum extract significantly promotes the characteristics of root morphological because of higher levels of indole-3-acetic acid (IAA). These results demonstrate the effectiveness of stability characteristics of commercially available algae extracts predicting the cellular targets prior to commercialization (Figure 1).
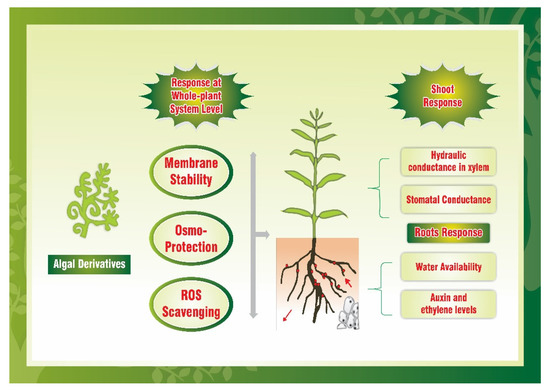
Figure 1. The key metabolic processes at the whole-plant level and physiological level are targeted by algal-based biostimulants [8].
In addition, the major plant growth traits, grain yield and its components were seen in two pepper varieties where Cladosporium sphaerospermum was applied to the seedlings [9]. Similar results were observed in tobacco exposed to C. sphaerospermum with significantly maximum plant growth. This may be due to induction of putative biochemical and molecular pathways like, photosynthesis, phytohormone homeostasis, cell expansion, and defense responses. In regard to ornamental crops, animal-based PH significantly affected the agronomical and morpho-physiological behavior of snapdragon hybrids as foliar pulp or substratum drench in three concentrations (0, 0.1 and 0.2 g L−1) [10]. At both PB levels, animal PH treated plants particularly at the substratum drench, boosted quality traits and the ornamental characteristics of plants in cultivar-dependent manner in comparison to the control.
In contrast to microbial and non-microbial stimulation activity, PBs may have dual effects involving resistance to diverse range of stresses by using certain natural substances or microorganisms. Sharma et al. [11] reported that exogenous use of jasmonic acid may enable the mustard (Brassica juncea) seedlings to be recovered from adverse effects of oxidative damage induced by pesticides across the enhanced expression of P450, RUBISCO, CXE, and NADH by inducing plant’s antioxidant defense mechanism. Likewise, after challenged environments of Fusarium oxysporum f. sp. lycopersici, bio-priming of Trichoderma erinaceum stimulated the defense transcriptome in tomato, where the following adaptations: (i) enhanced accumulation of defensive proteins, like WRKY (a category of proteins bound with DNA) [12], (ii) enhanced antioxidant defense mechanism, and (iii.) increased accumulation of lignification of the cells resulting in increased plant growth [13][14]. At last, Dal Cortivo et al. [15] demonstrated that the antagonist of fungicide action, i.e., sedaxane, a succinate dehydrogenase enzyme showed a significant hormonal activity in maize seeds which were related to auxin and gibberellin. The authors suggested that the application of sedaxane may promote root production and enhance the N accumulation and phenylpropanoid metabolism in young maize seedlings, thereby eliminating abiotic stress constraints at early development stages. Thus, PBs has tremendous potential to mitigate the toxic effects of synthetic pesticides via. Enhanced expression of genes governing tolerance to oxidative damage.
3. Role of Biostimulants on Nutrient Use Efficiency of Plants
A valuable tool in improving the availability of soil nutrients, incorporation, and assimilation can easily be achieved by the application of bioactive natural products and microbial inoculants [16]. Increased productivity of resource usage is very important for environmental and economic concerns [17]. The use of legume-based PH, particularly as substrate drench, strengthened the leaf number, SPAD index (Soil Plant Analyzing Development), and foliage production in greenhouse tomatoes under optimal and suboptimal N regions (110, and 6 mg/L, respectively) [18]. Improve agronomic responses of the tomato treated with PH was coupled with the root stimulation that facilitated N absorption and translocation. In addition, the PH application was improved under suboptimal N concentrations by up-regulation of expression of genes which are considered to be active in N assimilation for amino acids transporters and ferredoxin-glutamate (NADH/NADPH dependent) and glutamine synthesis in the roots. In contrast, Fiorentino et al. [19] investigated the biostimulant behavior of the two Trichoderma strains (T. virens GV41 or T. harzianum T22) under ideal growth conditions of N availability in two leafy vegetables, rocket, and lettuce. They observed that T. virens GV41 improved NUE in lettuce and promoted the native N uptake both in vegetables and soil. The effect was more pronounced in lettuce in a species-dependent manner. The results also revealed that inoculation from Trichoderma significantly modulated the structure of rhizosphere eukaryotic populations by providing considerable impacts with suboptimal N in contrast with N fertilized treatments. Furthermore, bacterial inoculants promotes the nutrients availability and their mobilization by the plant. Koskey et al. [18] identified 42 rhizobial isolates from the soil for characterization of biochemical, morpho-cultural, and genetic traits in the modified root nodules of climbing bean cultivars. They identified the bacterial strains with apparent biofertilizer properties capable of performing under stress conditions. The application of complex microbial inoculants comprising of genera, viz., Azospirillum, Streptomyces, Bacillus, Trichoderma, Pseudomonas, and R. irregularis were considered effective in wheat production in comparison to chemical fertilizers and the commercial minerals at the recommendation dose for field application distinguished by marginal P and N deficient soils [20]. The solubilization of zinc (Zn) by PGPR is practically a recent method to validate this method, a researcher team expertise not only screened but also analyzed the Zn-solubilizing rhizobacteria from sugarcane and wheat [21]. They found that microbial inoculants (Enterobacter cloacae, Pseudomonas fragi, Pantoea sp.) has tremendous potential as a microbe-based biostimulants to minimize the Zn deficiency under mineral deficit soil conditions.
References
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; et al. A vegetal biopolymer-based biostimulant promoted root growth in melon while triggering brassinosteroids and stress-related compounds. Front. Plant Sci. 2018, 9, 472.
- Singh, U.B.; Malviya, D.; Khan, W.; Singh, S.; Karthikeyan, N.; Imran, M.; Rai, J.P.; Sarma, B.K.; Manna, M.C.; Chaurasia, R.; et al. Earthworm grazed-Trichoderma harzianum biofortified spent mushroom substrates modulate accumulation of natural antioxidants and bio-fortification of mineral nutrients in tomato. Front. Plant Sci. 2018, 9, 1017.
- Mahnert, A.; Haratani, M.; Schmuck, M.; Berg, G. Enriching beneficial microbial diversity of indoor plants and their surrounding built environment with biostimulants. Front. Microbiol. 2018, 9, 2985.
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655.
- Ertani, A.; Francioso, O.; Tinti, A.; Schiavon, M.; Pizzeghello, D.; Nardi, S. Evaluation of seaweed extracts from Laminaria and Ascophyllum nodosum spp. as biostimulants in Zea mays L. using a combination of chemical, biochemical and morphological approaches. Front. Plant Sci. 2018, 9, 428.
- Palumbo, G.; Schiavon, M.; Nardi, S.; Ertani, A.; Celano, G.; Colombo, C.M. Biostimulant potential of humic acids extracted from an amendment obtained via combination of Olive Mill Wastewaters (OMW) and a pre-treated organic material derived from Municipal Solid Waste (MSW). Front. Plant Sci. 2018, 9, 1028.
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5.
- Li, Z.T.; Janisiewicz, W.J.; Liu, Z.; Callahan, A.M.; Evans, B.E.; Jurick, W.M.I.; Dardick, C. Exposure in vitro to an Environmentally Isolated Strain TC09 of Cladosporium sphaerospermum triggers plant growth promotion, early flowering, and fruit yield increase. Front. Plant Sci. 2019, 9, 1959.
- Cristiano, G.; Pallozzi, E.; Conversa, G.; Tufarelli, V.; De Lucia, B. Effects of an animal-derived biostimulant on the growth and physiological parameters of potted snapdragon (Antirrhinum majus L.). Front. Plant Sci. 2018, 9, 861.
- Sharma, A.; Kumar, V.; Yuan, H.; Kanwar, M.K.; Bhardwaj, R.; Thukral, A.K.; Zheng, B. Jasmonic acid seed treatment stimulates insecticide detoxification in Brassica juncea L. Front. Plant Sci. 2018, 9, 1609.
- Aamir, M.; Kashyap, S.P.; Zehra, A.; Dubey, M.K.; Singh, V.K.; Ansari, W.A.; Upadhyay, R.S.; Singh, S. Trichoderma erinaceum bio-priming modulates the wrkys defense programming in tomato against the Fusarium oxysporum f. sp. lycopersici (Fol) challenged condition. Front. Plant Sci. 2019, 10, 911.
- Cramer, G.R.; Urano, K.; Velrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163.
- De Vasconcelos, A.C.F.; Chaves, L.H.G. Biostimulants and their role in improving plant growth under abiotic stresses. In Biostimulants in Plant Science; Mirmajlessi, S.M., Radhakrishnan, R., Eds.; IntechOpen: London, UK, 2020.
- Cortivo, C.D.; Conselvan, G.B.; Carletti, P.; Barion, G.; Sella, L.; Vamerali, T. Biostimulant effects of seed-applied sedaxane fungicide: Morphological and physiological changes in maize seedlings. Front. Plant Sci. 2017, 8, 8.
- Huang, J.; Levine, A.; Wang, Z. Plant abiotic stress. Sci. World J. 2013, 2013, 432836.
- Rouphael, Y.; Colla, G.; Graziani, G.; Ritieni, A.; Cardarelli, M.; De Pascale, S. Phenolic composition, antioxidant activity and mineral profile in two seed-propagated artichoke cultivars as affected by microbial inoculants and planting time. Food Chem. 2017, 234, 10–19.
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in agriculture. Front. Plant Sci. 2020, 11, 40.
- Trejo-Téllez, L.I.; Estrada-Ortiz, E.; Gómez-Merino, F.C.; Becker, C.; Krumbein, A.; Schwarz, D. Flavonoid, nitrate and glucosinolate concentrations in Brassica species are differentially affected by photosynthetically active radiation, phosphate and phosphite. Front. Plant Sci. 2019, 10, 371.
- Kamran, S.; Shahid, I.; Baig, D.N.; Rizwan, M.; Malik, K.A.; Mehnaz, S. Contribution of zinc solubilizing bacteria in growth promotion and zinc content of wheat. Front. Microbiol. 2017, 8, 2593.
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393.
- Soppelsa, S.; Kelderer, M.; Casera, C.; Bassi, M.; Robatscher, P.; Andreotti, C. Use of biostimulants for organic apple production: Effects on tree growth, yield, and fruit quality at harvest and during storage. Front. Plant Sci. 2018, 9, 1342.

