 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Prathyaya Ramesh | + 2401 word(s) | 2401 | 2021-03-22 07:27:24 | | | |
| 2 | Lily Guo | -8 word(s) | 2393 | 2021-04-02 08:45:34 | | |
Video Upload Options
Cytokines are key molecules within the tumor microenvironment (TME) that can be used as biomarkers to predict the magnitude of anti-tumor immune responses. During immune monitoring, it has been customary to predict outcomes based on the abundance of a single cytokine, in particular IFN-γ or TGF-β, as a readout of ongoing anti-cancer immunity. However, individual cytokines within the TME can exhibit dual opposing roles. For example, both IFN-γ and TGF-β have been associated with pro- and anti-tumor functions. Moreover, cytokines originating from different cellular sources influence the crosstalk between CD4+ and CD8+ T cells, while the array of cytokines expressed by T cells is also instrumental in defining the mechanisms of action and efficacy of treatments. Thus, it becomes increasingly clear that a reliable readout of ongoing immunity within the TME will have to include more than the measurement of a single cytokine.
1. Introduction
Cytokines are intercellular signaling molecules that control critical biological functions such as cell proliferation, differentiation, survival, cell cycle progression, and immune-cell activity [1]. Based on their function, cytokines can be divided into subgroups: lymphokines, monokines, chemokines, interleukins (IL), and interferons (IFNs) [2]. These secreted proteins act at short range as humoral regulators, attuning the functional processes of individual cells to maintain cellular homeostasis [1]. Importantly, cytokines are key mediators of anti-tumor immune responses by controlling activation of immune cells [2][3]. As such, the abundance and type of cytokines within the tumor microenvironment (TME) is an important biomarker to predict tumor progression [1]. However, further studies are required to fully elucidate the different biological functions induced by the presence of a single versus multiple cytokines, and by the “cross-talk” between cytokine-mediated signaling pathways [1].
Typically, the TME is characterized by presenting high levels of the anti-inflammatory, immunosuppressive cytokine IL-10, which facilitates tumor expansion [3]. Meanwhile, IFNs hold anti-tumor, anti-viral, and immunomodulatory properties [4], with IFN-gamma (IFN-γ) playing a critical role in defining a pro- versus anti-TME. IFN-γ has long been studied as the ultimate pro-inflammatory cytokine, with the responsibility to regulate anti-inflammatory responses. It also inhibits differentiation of immunosuppressive regulatory T cells (Tregs), and balances tissue destruction in chronic inflammation conditions [5][6]. Interestingly, depending on the biological context, IFN-γ can enhance or diminish the expression of tumor antigens, thereby determining the extent of anti-tumor immune responses [7]. Thus, some pro-tumor consequences of IFN-γ exposure exist, and its role may depend on the length of exposure and absolute abundance of the cytokine within the TME [5]. Additionally, while necessary, this cytokine does not seem to be sufficient on its own to halt the development of tumors [6].
Transforming growth factor-beta (TGF-β) operates in different capacities throughout disease progression. TGF-β can function as a tumor suppressor, inducing apoptosis and suppressing proliferation of pre-malignant and cancer cells [8]. However, as the name suggests, TGF-β can also mediate pro-metastatic and tumorigenic responses and support an immunosuppressive TME. TGF-β can help control inflammation and mediate immune tolerance [8]. The interchange of TGF-β with IFN-γ can be indicative of the effectiveness of cancer therapies and of an immune-active TME. However, the dual face of these cytokines is emblematic as to why a single cytokine is not satisfactory to adequately determine the ongoing immunity of the TME. Besides IL-10, TGF-β, and IFN-γ, it is the concerted action of multiple cytokines within the TME that mainly determines the extent and success of anti-tumor immune responses. Cytokines allow immune cells to transmit signals to each other and, under ideal conditions, promote a vigorous and harmonized response against target cells in both a paracrine and autocrine manner over short distances [9][10]. The anti-tumor activities of several cytokines have been established in animal models, serving as a premise for several cytokine-based immunotherapies [10]. Moreover, T cell engineering can be used for the expression of an antigen specific CAR or TCR to drive responses selectively towards tumor cells [11]. The pattern of cytokines expressed in response to immune cell therapy can then be used as a readout for anti-tumor control.
As important as cytokines are in predicting tumor progression and the success of cancer therapies, they can provide the identity and abundance of immune cells infiltrating the TME. Tumor infiltrating lymphocyte (TIL) populations include CD3+CD4+ and CD3+ CD8+ T cells, which can each be engaged in suppressing tumor growth [12][13]. B cells can also be present in the TME, and among them, regulatory B cells are known for the release of immunoregulatory cytokines that hinder anti-tumor responses by inhibiting natural killer (NK) cells and CD8+ cytotoxic T cells [14]. Inflammatory cells within the TME can either actively hinder tumor development or contribute to tumor progression [12]. More ambiguous roles have been assigned to T helper 2 (Th2) CD4+ T cell populations, which produce IL-4, IL-5, and IL-3 in support of B and T helper 17 (Th17) cell responses [15]. Thus, it will be imperative to expand our understanding of cytokine function to understand the true breadth and role that these biomarkers play in the activation of these immune cells and establishing anti-tumor immunity.
Identifying a defined palette of cytokines that can predict immune activation versus immunosuppression enables a better characterization of the TME. Here, we provide a magnified perspective of the cytokine patterns that represent the inner workings of immune cells, and specifically T cells responding within the TME. The observation of different secreted cytokine patterns can be associated with respective outcomes, and then used to accurately evaluate the effectiveness of immune-based therapies.
2. T Cell-Derived Cytokines as Biomarkers of Anti-Tumor Activity
T cells are a notable indicator for characterizing ongoing immunity within the tumor microenvironment. However, T cell abundance is not an adequate marker of whether tumors will regress. Figure 1 showcases a human melanoma tumor with an abundance of T cells. This exemplifies the need to example T cell function over T cell abundance to understand the nature of tumor progression.
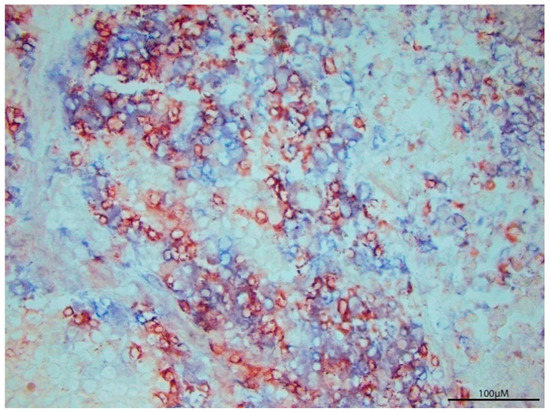
Figure 1. T cell Infiltration per se does not define tumor regression. P607 tumor tissue was surgically resected from a metastatic melanoma patient, revealing abundant T cell infiltration by immunoperoxidase labeled anti-CD8 (red) and alkaline phosphatase labeled anti-CD3 staining (blue). Cytokine analysis of tumor homogenates can reveal the prevailing immune response to better understand tumor expansion despite heavy T cell presence.
A combination of cytokine markers can provide reliable information on the capacity of T cells to act within the tumor. T cells are the primary source of each cytokine and these profiles are used to define T cell subsets [16]. Specific combinations of cytokines are associated with Th1, Th9, and Th17 T cells. As an additional parameter, most studies will include CD107A as a surrogate marker of cytotoxic activity.
In reference to the Th1 subset, the ratio of Th1 to Th2-associated cytokines provides information on the outcome of ongoing antitumor immune responses [17]. Representative cytokines for the Th1 subset, with classic anti-tumor activity include IFN-γ and IL-2 [18] and the Th1/Th2 ratio is critical for numerous immune responses [19]. Showing the ability of antigen specific Th1 cells through the use of adhesion molecules places the Th1 subset at the forefront of tumor rejection activity [17]. Th1 cells primarily stimulate CTL activation and differentiation and the secretion of cytokines such as IL-2, IL-12, IFN-γ, and TNF-α support CTL effector phenotype [20]. Additionally, Th1 therapy can induce lasting immunological memory to revive CTL generation upon tumor recurrence [17]. Critical to this work is the discovery that Th1 cells expressed adhesion molecules LFA-1/ICAM-1 [19], that allow transmigration into tumor tissues across tumor vessels, supporting tumor metastasis [17][19]. The cytokines secreted by Th1 T cells are commonly associated with a good prognosis for anti-tumor therapies [20]. Moreover, Th1 cytokines produced upon antigen stimulation directly induce the recruitment of effector cells such as CD8+ T cells, NKT cells, and NK cells to the tumor [17].
Th9 cytokines were designated as a separate T cell subset preferentially producing IL9 [18]. However, IL9 secretion per se is not restricted to Th9 cells, and can also be produced by Treg, Th17, or Th2 cells [18]. IRF4 expression combined with TGF-β and IL-4 exposure are critical for Th9 differentiation [18]. IL-9 is implicated in the anticancer effects of Th9 cells [18], whereas IL-2 supports the production of IL-9 [21]. The cytokine enhances dendritic cell survival and strengthens their capacity to induce and foster protective immunity [18]. Typically, Th9 cells produce low levels of IL-21, and also generate IL-25 and IL-1, but the anticancer effects of Th9 cells predominantly depend on IL-9 [18]. Originally assigned a function in combating parasitic infections, Th9 cells have been designated as anti-tumor T cells and their IL-9 expression can potentially induce bystander cell recruitment (such as NK cells and CD8 cells) [18]. Transcription factors, including STAT6, GATA3, PU.1, and IRF4, are central to Th9 polarization [18]. Of these, GATA3 and STAT6 have also been detected in Th2 cells. In other reports, IL-9 has been identified for targeted proliferation and activation of mast cells and enhancement of dendritic cell survival [18], highlighting IL-9 as a nominal marker for antitumor activity.
The Th17 cell subset does not exhibit stable differentiation and rather adorns a plasticity which allows it to adapt to its surroundings [22]. Th17 cells were identified as an individual subset based on their production of IL-17 [22]. Additionally, this subset produces IL-17F, IL-21, GM-CSF, and IL-22, whereas IL-23 is required for Th17 cell survival and pathogenesis [22]. Th17 cells have predominantly been associated with autoimmune and inflammatory diseases [22]. The plasticity of this subset places its role in cancer under debate, as Th17 can transdifferentiate into other subsets. Th1 conversion supports anti-tumor value, but Th17 can also morph into TR1, Th2, or TFH cells. Thus, Th17 are equipped to mediate several contradicting functions [22]. Within melanoma however, Th17 cells tend to spur protective immunity and establish tumor regression [22]. Within the TME, Th17 cells did not exhibit killing activity, but instead promoted dendritic cell and CD4+ and CD8+ T cell recruitment [22]. Th17 T cell function is thus difficult to classify due to its plasticity.
CD107a (also known as LAMP-1) is not a cytokine, but serves as a substitute marker of cytotoxic activity expressed by natural killer (NK), γδ T cells, and cytotoxic T cells, briefly found on the cell surface upon degranulation [23][24]. NK cells help mediate tumor cell clearance and viral infections [25]. Expression of CD107a is aligned with IFN-γ and TNF-α expression and cytotoxicity by T cells, and is likewise upregulated on activated NK cells, thus functioning as a general NK functional activity marker [25].
Besides T cell derived cytokines themselves, the cytokines that induce T cell activity can also be informative of the tumor microenvironment and can be used as treatments as well. Therapeutic cytokines for tumor patients can be subclassified into the IL-1, IL-2, and IL-12 families [26]. IL-1 is categorized as a pro-inflammatory cytokine and works as a T cell co-stimulator along with its role in inducing effector cell proliferation [26]. IL-2 was originally discovered as T cell growth factor and is key in promoting CD8+ mediated antitumor immunity [26], stimulating T cell growth, differentiation of effector T cells and Th1 cells, and activation of NK cells. In fact, it is the most used cytokine in cancer therapy [26]. Finally, IL-12 is capable of inducing Th1 and IFN-γ responses (activates JAK-STAT pathway after binding to its receptor) within T cells [26]. IL-12 is produced by dendritic cells and macrophages and like the other Th1 cytokines, plays a role in NK cell activation and Th1 differentiation [26]. IL-12 tends to display limited therapeutic potential, though IL-12-producing chimeric antigen receptor (CAR)-modified T cells potently destroy existing tumors [26].
Among T cell-derived biomarkers, Th1 cytokines IFN-γ, IL-2, and TNF-α present themselves as the most notable markers of responses to therapy [18][25]. The synergy of these and other cytokines (in a polyfunctional manner) as outlined in Table 1, can produce positive, potent, and efficient responses to cytokine therapies. Table 1 serves to highlight the most commonly measured cytokine responses across different tumor types with T cell-based therapeutics. By relating these secondary outcome measures to treatment outcomes, we create a foundation for a palette of cytokines that can better serve to relate therapeutic outcomes. From this table, we conclude that detecting a combination of IFN-γ, TNF-α and IL-2 is more informative than measuring a single cytokine such as IFN-γ alone. Moreover, the function of a cytokine within the tumor environment can be ambivalent and several factors can affect their in vivo functionality.
Table 1. Correlating cytokine profiles and T cell responses.
| Study | Cytokines Probed and Monitoring Methods | Patients/Source of T Cells | Cancer Type and Treatment | T cell Treatment Outcome Response and Correlated Cytokine |
|---|---|---|---|---|
| #1 [27] | IFN-γ, IL-2, TNF-α FACS analysis, ICS, ELISpot |
Healthy donors | CD3+ T cells isolated from peripheral blood | IL-2 (mostly among CD8− T cells) and IFN-γ secreting cells increased, TNF-α secreting cells decreased. IFN-γ and IL-2 secreting cytokines showed functional state persistence. |
| #2 [28] | TNF-α IFN-γ, IL-10, IL-17, IL-2 Intracellular cytokine staining of CD4+ and CD8+ T cells, in renal parenchyma tissues |
Peripheral blood, fresh tumor, and autologous renal parenchyma | Renal cell carcinoma PBMC and TIL thawed and analyzed for cytokine release. |
IL-10 increased among CD4+ and CD8+ subsets; TNF-α (CD4+ and CD8+), IFN-γ (CD8+) increased after activation. Some patients had increased IL-17 in CD4+ TIL. CD107a surface expression found in CD8+ and some CD4+ cells post- activation. Cytokine secretion pattern of responders: TNF-α, IFN-γ, IL-2 with little IL-5. |
| #3 [29] | TNF, IFN-γ, CD107a (cytotoxicity marker). FACS analysis, cytotoxicity assays, phenotype analysis, flow cytometry |
Serial blood sample obtained from TIL treated patients | Melanoma IL-2 based TIL therapy |
CD8+ T cells expressing CD107a were fewer than cytokine producing cells. Most CD107a + cells also produced one cytokine. |
| #4 [30] | IFN-γ, TNF-α, CCL3 IFNγ ELISPOT assay, FlowJo |
22 CMV seropositive patients | Glioblastoma In vitro generation of (CMV) pp65 T cells and CMV pp65- DCs from PBMCs |
Patients receiving CMV pp65 T cells had more IFNγ+, TNFα+ CCL3+ pp65 specific CD8+ T cells. Survival in treated patients correlated with expression of IFNγ, TNFα and CCL3. |
| #5 [31]. | IFN-γ2,TNF-α1,2, IL-2, IL-12, IL-18, IL-21 CCL41,2, CD107a2(cytotoxicity marker) Flow cytometry, ELISA, Bio-Plex |
Bulk ascites cell preparations from high-grade serous EOC patients | Epithelial ovarian cancer (EOC) Exogenous cytokine therapy and introduction of EOC ascites environment on T—cell polyfunctionality |
IL-1+, IL-12+ IL-18 enhanced IFNγ (by CD8+ cells), TNF-α, and CCL4 expression Cytokine combination synergistically induced polyfunctional responses and decreased cytokine negative or monofunctional T cells. |
| #6 [32] | IFN-γ3, TNF-α3 IL-23 Flow cytometry, immunohistochemistry |
25 treatment- naïve NSCLC patients with clinical stage I-Iva tumors. | Non-small cell lung cancers (NSCLC) TIL therapy |
CD4+/CD8+ cells producing either 2 or 3 of the cytokines were most informative. TNFα and IL-2 were crucial to T cell mediated immunity. |
References
- Foster, J.R. The functions of cytokines and their uses in toxicology. Int. J. Exp. Pathol. 2001, 82, 171–192.
- Bonini, C.; Mondino, A. Adoptive T-cell therapy for cancer: The era of engineered T cells. Eur. J. Immunol. 2015, 45, 2457–2469.
- Germano, G.; Allavena, P.; Mantovani, A. Cytokines as a key component of cancer-related inflammation. Cytokine 2008, 43, 374–379.
- Castro, F.; Cardoso, A.P.; Goncalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-Gamma at the Crossroads of Tumor Immune Surveillance or Evasion. Front. Immunol. 2018, 9, 847.
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-γ in tumor progression and regression: A review. Biomark. Res. 2020, 8, 49.
- Jennifer Gillary Segal, N.C.L.; Tsung, Y.L.; Jeffrey, A.; Norton, K.T. The Role of IFN-y in Rejection of Established Tumors by IL-12: Source of Productiona and Target. Cancer Res. 2002, 62, 4696–4703.
- Le Poole, I.C.; Riker, A.I.; Quevedo, M.E.; Stennett, L.S.; Wang, E.; Marincola, F.M.; Kast, W.M.; Robinson, J.K.; Nickoloff, B.J. Interferon-gamma reduces melanosomal antigen expression and recognition of melanoma cells by cytotoxic T cells. Am. J. Pathol. 2002, 160, 521–528.
- Batlle, E.; Massague, J. Transforming Growth Factor-beta Signaling in Immunity and Cancer. Immunity 2019, 50, 924–940.
- Conlon, K.C.; Miljkovic, M.D.; Waldmann, T.A. Cytokines in the Treatment of Cancer. J. Interferon Cytokine Res. 2019, 39, 6–21.
- Lee, S.; Margolin, K. Cytokines in cancer immunotherapy. Cancers 2011, 3, 3856–3893.
- Anderson, K.G.; Stromnes, I.M.; Greenberg, P.D. Obstacles Posed by the Tumor Microenvironment to T cell Activity: A Case for Synergistic Therapies. Cancer Cell 2017, 31, 311–325.
- Whiteside, T.L. The tumor microenvironment and its role in promoting tumor growth. Oncogene 2008, 27, 5904–5912.
- Pennock, N.D.; White, J.T.; Cross, E.W.; Cheney, E.E.; Tamburini, B.A.; Kedl, R.M. T cell responses: Naive to memory and everything in between. Adv. Physiol. Educ. 2013, 37, 273–283.
- Yuen, G.J.; Demissie, E.; Pillai, S. B lymphocytes and cancer: A love-hate relationship. Trends Cancer 2016, 2, 747–757.
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The tumor microenvironment at a glance. J. Cell Sci. 2012, 125, 5591–5596.
- Raphael, I.; Nalawade, S.; Eagar, T.N.; Forsthuber, T.G. T cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cytokine 2015, 74, 5–17.
- Knutson, K.L.; Disis, M.L. Tumor antigen-specific T helper cells in cancer immunity and immunotherapy. Cancer Immunol. Immunother. 2005, 54, 721–728.
- Vegran, F.; Apetoh, L.; Ghiringhelli, F. Th9 cells: A novel CD4 T-cell subset in the immune war against cancer. Cancer Res. 2015, 75, 475–479.
- Nishimura, T.; Iwakabe, K.; Sekimoto, M.; Ohmi, Y.; Yahata, T.; Nakui, M.; Sato, T.; Habu, S.; Tashiro, H.; Sato, M.; et al. Distinct Role of Antigen-specific T Helper Type 1 (Th1) and Th2 Cells in Tumor Eradication in Vivo. Rockefeller Univ. Press 1999, 190, 617–627.
- van Belzen, I.; Kesmir, C. Immune biomarkers for predicting response to adoptive cell transfer as cancer treatment. Immunogenetics 2019, 71, 71–86.
- Goswami, R.; Kaplan, M.H. A brief history of IL-9. J. Immunol. 2011, 186, 3283–3288.
- Guery, L.; Hugues, S. Th17 Cell Plasticity and Functions in Cancer Immunity. Biomed. Res. Int 2015, 2015, 314620.
- Badri, K.R.; Gao, L.; Hyjek, E.; Schuger, N.; Schuger, L.; Qin, W.; Chekaluk, Y.; Kwiatkowski, D.J.; Zhe, X. Exonic mutations of TSC2/TSC1 are common but not seen in all sporadic pulmonary lymphangioleiomyomatosis. Am. J. Respir. Crit. Care Med. 2013, 187, 663–665.
- Silva-Santos, B.; Serre, K.; Norell, H. γδ T cells in cancer. Nat. Rev. Immunol. 2015, 15, 683–691.
- Alter, G.; Malenfant, J.M.; Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 2004, 294, 15–22.
- Xu, H.-M. Th1 cytokine-based immunotherapy for cancer. Hepatobiliary Pancreat. Dis. Int. 2014, 13, 482–494.
- Han, Q.; Bagheri, N.; Bradshaw, E.M.; Hafler, D.A.; Lauffenburger, D.A.; Love, J.C. Polyfunctional responses by human T cells result from sequential release of cytokines. Proc. Natl. Acad. Sci. USA 2012, 109, 1607–1612.
- Attig, S.; Hennenlotter, J.; Pawelec, G.; Klein, G.; Koch, S.D.; Pircher, H.; Feyerabend, S.; Wernet, D.; Stenzl, A.; Rammensee, H.G.; et al. Simultaneous infiltration of polyfunctional effector and suppressor T cells into renal cell carcinomas. Cancer Res. 2009, 69, 8412–8419.
- Donia, M.; Kjeldsen, J.W.; Andersen, R.; Westergaard, M.C.W.; Bianchi, V.; Legut, M.; Attaf, M.; Szomolay, B.; Ott, S.; Dolton, G.; et al. PD-1(+) Polyfunctional T Cells Dominate the Periphery after Tumor-Infiltrating Lymphocyte Therapy for Cancer. Clin. Cancer Res. 2017, 23, 5779–5788.
- Reap, E.A.; Suryadevara, C.M.; Batich, K.A.; Sanchez-Perez, L.; Archer, G.E.; Schmittling, R.J.; Norberg, P.K.; Herndon, J.E., 2nd; Healy, P.; Congdon, K.L.; et al. Dendritic Cells Enhance Polyfunctionality of Adoptively Transferred T Cells That Target Cytomegalovirus in Glioblastoma. Cancer Res. 2018, 78, 256–264.
- Tran, E.; Nielsen, J.S.; Wick, D.A.; Ng, A.V.; Johnson, L.D.; Nesslinger, N.J.; McMurtrie, E.; Webb, J.R.; Nelson, B.H. Polyfunctional T-cell responses are disrupted by the ovarian cancer ascites environment and only partially restored by clinically relevant cytokines. PLoS ONE 2010, 5, e15625.
- De Groot, R.; Van Loenen, M.M.; Guislain, A.; Nicolet, B.P.; Freen-Van Heeren, J.J.; Verhagen, O.; Van Den Heuvel, M.M.; De Jong, J.; Burger, P.; Van Der Schoot, C.E.; et al. Polyfunctional tumor-reactive T cells are effectively expanded from non-small cell lung cancers, and correlate with an immune-engaged T cell profile. Oncoimmunology 2019, 8, e1648170.

