 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Eiji Takahashi | + 4925 word(s) | 4925 | 2020-04-22 10:45:28 | | | |
| 2 | Rita Xu | -931 word(s) | 3994 | 2020-04-29 06:24:48 | | | | |
| 3 | Rita Xu | -1141 word(s) | 2853 | 2020-05-13 04:41:59 | | | | |
| 4 | Rita Xu | -35 word(s) | 2818 | 2020-10-29 08:45:46 | | |
Video Upload Options
Hematogenous tumor metastasis begins with the invasion and spread of primary tumor cells in the local tissue leading to intravasation. Directional migration of primary cancer cells toward intratumor blood/lymphatic vessels should elevate the probability for intravasation and ultimate hematogenous metastasis. Many presume, on the analogy of chemotaxis by specific chemoattractants, that concentration gradients of energy substrates/metabolites in tumor tissue would be a guiding cue for directional cell migration, whereas strong experimental evidence is scarce at present. Here, using a novel microfluidic device, we clearly demonstrated that the gradient of extracellular pH is a cue for directional migration of MDA-MB-231 cells in vitro.
1. Introduction
The presence of metastasis is an important prognostic factor in the majority of tumor patients. Distant metastasis begins with the intravasation of primary tumor cells into intratumoral blood and lymphatic vessels. Local tumor cell proliferations elevate hydrostatic pressure in the interstitium, which tends to push tumor cells into microvessels. Tumor angiogenesis and lymphangiogenesis further enhance passive intravasation. In addition to passive cell movement, active cell migrations guided by chemokines are also occurring, in which cancer-associated fibroblasts play a role [1][2][3].
Considering the analogy of chemotaxis, where component movement depends on the concentration gradient of specific chemoattractants, it has long been postulated that the gradients of nutrients and metabolic waste might guide tumor cell migration. This prediction is reinforced given the metabolic characteristics peculiar to solid tumors. In solid tumors, the structures of blood vessels are immature and their topological configuration in tissues is generally irregular [4]. As a result, the convectional and diffusional transports of substances between individual cells and blood are considerably hindered and steep concentration gradients of nutrients and metabolic wastes are established within the tissue [4][5]. Although <0.1% of primary tumor cells succeed in intravasation in the local tissue [6], directional cell migration guided by these metabolic gradients, if any, could elevate the probability of intravasation and, ultimately, hematogenous metastasis. However, evidence indicating the status of extracellular nutrient/metabolite gradients as guiding cues for cancer cell migration is scarce at the present time.
Given the above, our initial hypothesis is that the metabolic gradients in the tissue might be a cue for guiding the tumor cells to nearby blood vessels. Because acidosis and hypoxia are the hallmark of an invasive hypoxic tumor [7][8], we focused, among numerous cues in tissues [9], on H+ and oxygen particularly their concentration gradients. Hereafter, we refer to the gradient of pH and oxygen concentration as metabolic gradients.
To test this hypothesis in vitro, we used a glassware product that we previously devised and call gap cover glass (GCG; Figure 1) [10] to apply in producing metabolic gradients in cultured monolayer cells. In fact, we demonstrated that gradients of extracellular pH with a magnitude of 0.2–0.3 units/mm are established at three hours after placing the GCG onto confluent monolayer MDA-MB-231 cells [11], while gradients of oxygen concentration under the GCG has not yet been demonstrated. Furthermore, we demonstrated a tendency that MDA-MB-231 cells migrate toward the opening of GCG; in other words, toward areas of higher pH and/or oxygen concentration. One of the serious drawbacks of our approach, however, was that migrating cells under the GCG frequently underwent collisions with each other and subsequently changed their direction of migration. Thus, we thought that our tentative conclusions regarding directional migration of MDA-MB-231 cells were far from promising. To overcome the collision artifacts, we refined our technique by combining our GCG with the conventional wound-healing assay for cell migration [12].
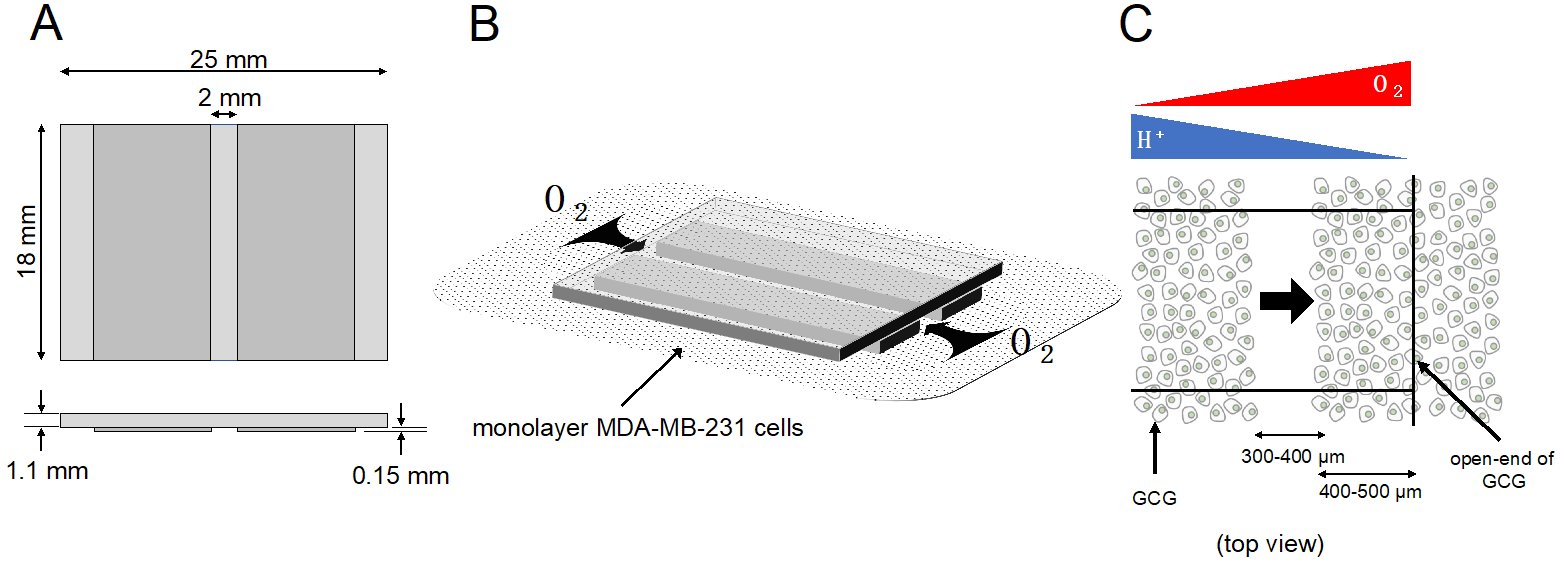
Figure 1. GCG. (A) Three thin glass plates were assembled into the GCG. (B) The GCG was gently placed onto the monolayer of MDA-MB-231 cells. The diffusional supply of energy substrates such as oxygen to cells in the narrow channel of the GCG is restricted. Thus, cellular oxygen consumption produces gradients of oxygen concentration in the extracellular medium in the narrow channel. Washout of metabolites such as CO2/H+ to the bulk medium is similarly restricted, and pH gradients are produced in the narrow channel. (C) GCG combined with wound-healing assay for cell migration. Gradients of oxygen and H+ concentration are also illustrated. In the present study, we demonstrated a vectorial movement of MDA-MB-231 cells toward the open-end of GCG (i.e., higher oxygen concentration or lower H+ concentration; indicated by the thick arrow).
We demonstrated with a new improved technique that MDA-MB-231 cells under GCG migrate toward higher pH/O2 regions while directional migration was abolished after eliminating gradients of pH. Thus, relatively small gradients of pH in the extracellular medium compared to those found in Na+/H+ exchanger-driven cell migration were sufficient to guide MDA-MB-231 cells in vitro.
2. Results
2.1. Extracellular Gradients of Oxygen Concentration under GCG
We successfully visualized the oxygen concentration gradient under GCG. As shown in Figure 2A, immediately after placing the GCG, the oxygen concentration along the oxygen diffusion path was constant up to 1200 µm inside the GCG. Subsequently, the oxygen concentration gradients developed slowly and, at six hours after placement of the GCG, a −6.5% O2/mm linear gradient was established under GCG. Magnitude of the oxygen concentration gradient increased in proportion to the number of cells per unit volume (Figure 2B).
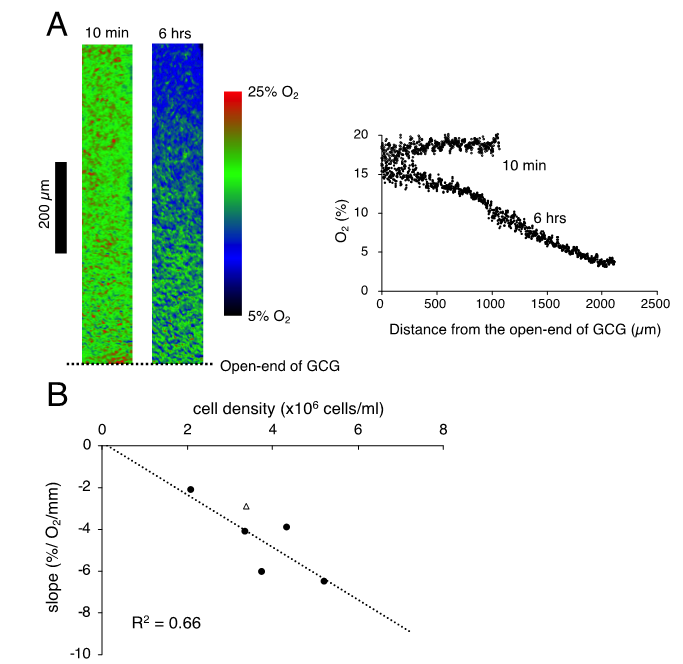
Figure 2. (A). Representative data indicating changes in oxygen concentration in the extracellular medium along the oxygen diffusion path as measured by the oxygen sensor foil. The ratio of red and green fluorescence is represented in pseudo-color. At 10 min after placing the gap cover glass (GCG), no gradient was demonstrated underneath. At six hours after placing the GCG, a −6.2% O2/mm linear oxygen concentration gradient was produced under the GCG. The number of MDA-MB-231 cells in the culture dish was 5.2 × 106 cells/mL. (B) Relationship between the magnitude of the oxygen concentration gradient and cell density. The linear relationship indicates that the oxygen concentration gradient depends upon oxygen consumption of the cell per unit volume. The open triangle represents the slope in which L-15 medium was buffered with 15 mM hepes.
2.2. Effects of Hepes-buffered L-15 on pH and Oxygen Gradients under GCG
As reported elsewhere [11], extracellular pH gradients along the diffusion path slowly developed in GCG and reached the steady state in three hours. Gradients of pH with a magnitude of 0.2–0.3 units/mm were consistently demonstrated. Supplementing hepes into the L-15 medium abolished the pH gradient (Figure 3).
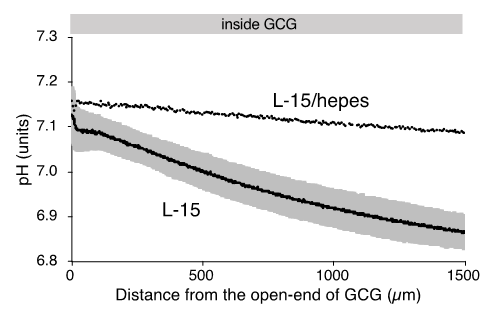
Figure 3. Gradients of extracellular pH under GCG in L-15 and in hepes (15 mM)-buffered L-15 medium. Average pH gradients in L-15 medium are shown where the range of ±standard deviation (SD) is represented in gray (n = 4). Data were collected three hours after placing the GCG.
2.3. Cell Migration
We completed five “without GCG” and five “with GCG” experiments, respectively, and representative data associated with these “without GCG” and “with GCG” experiments are shown in Supplementary Video S1 and Video S2, respectively. Figure 4 illustrates the trajectory of individual MDA-MB-231 cells for 24 h.
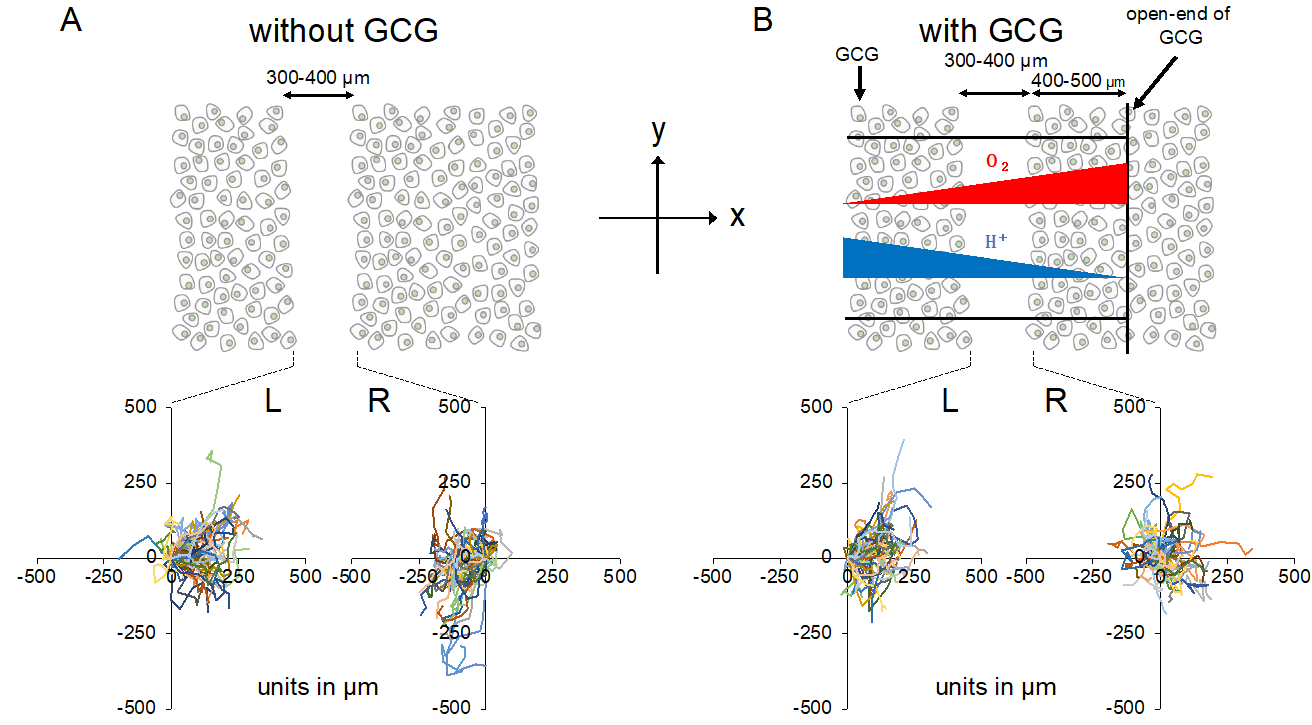
Figure 4. Trajectories of MDA-MB-231 cells measured for 24 h. The oxygen concentration in the microincubator was 21% (room air). Values presented in the graphs are in micrometers. (A) Without the GCG placed, cells migrated into the wound space equally from the right (R) and left (L) boundaries of the wound space. (B) With the GCG placed, the migration of the cells initially located at the right boundary (R) into the wound space was hindered and they even appeared to migrate in a direction opposite to that toward the wound space as if they were crawling into the crowd of cells. Trajectories of the 50 cells are superimposed.
Without involvement from the GCG, the L-cells (cells migrating into the wound space from the left boundary, see Figure 4) and R-cells (cells migrating into the wound space from the right boundary, see Figure 4) similarly migrated into the wound space (Figure 4A). In contrast, in the “with GCG” experiments, we demonstrated distinct differences in the direction of cell migration, particularly in the R-cells. With the inclusion of the GCG, the migration of R-cells into the wound space appeared to be considerably hindered (Figure 4B). These cells even migrated in the direction opposite of that toward the wound space, as if they were instead crawling into the crowd of cells. These data are consistent with that cells under the GCG demonstrate directional migration toward the open-end of the GCG (right side). Directional cell movement can be visually confirmed by observing Video S2.
We further checked the directional migration of the cells under GCG by counting the numbers of L-cells and R-cells in the wound space. As shown in Figure 5A, without the GCG in place, the numbers of L-cells and R-cells were the same throughout the experiment. In contrast, with the GCG in place, the numbers of R-cells were smaller than that of the L-cells (Figure 5B), reaching statistical significance after five hours. Thus, the numbers of L-cells and R-cells at 24 h were highly different from one another (Figure 5C).
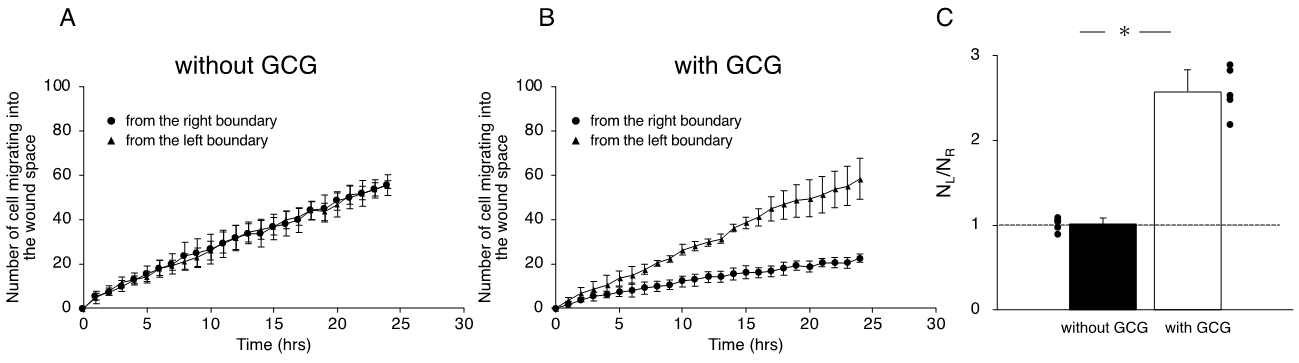
Figure 5. The number of cells that migrated into the wound space in 24 h. (A) Without the GCG, the numbers of cells migrating into the wound space from the left boundary and the right boundary were the same. (B) With the GCG, the numbers of cells migrating into the wound space from the left boundary and the right boundary were different, with a statistically significant difference demonstrated after five hours. (C) The ratio of the numbers of cells migrating into the wound space from the left boundary and the right boundary determined at 24 h (NL and NR, respectively). The directional migration of MDA-MB-231 cells was clearly demonstrated with involvement of the GCG. Error bars represent the SD. Data were accumulated from five independent experiments. * p < 0.05, as judged by Student’s t-test.
Next, we undertook another series of experiments in which the role of extracellular pH gradients in directional movements of MDA-MB-231 cells was examined. We completed five “L-15” and five “L-15/hepes” experiments, respectively. In the L-15 medium, the number of cells migrating from the left boundary into the wound space was significantly higher than that from the right boundary (Figure 6A,C). In comparison, in the L-15/hepes medium, such heterogeneity disappeared (Figure 6B,C). These results indicate that the extracellular pH gradient is the dominant cue for the directional migration of MDA-MB-231 cells under our GCG.
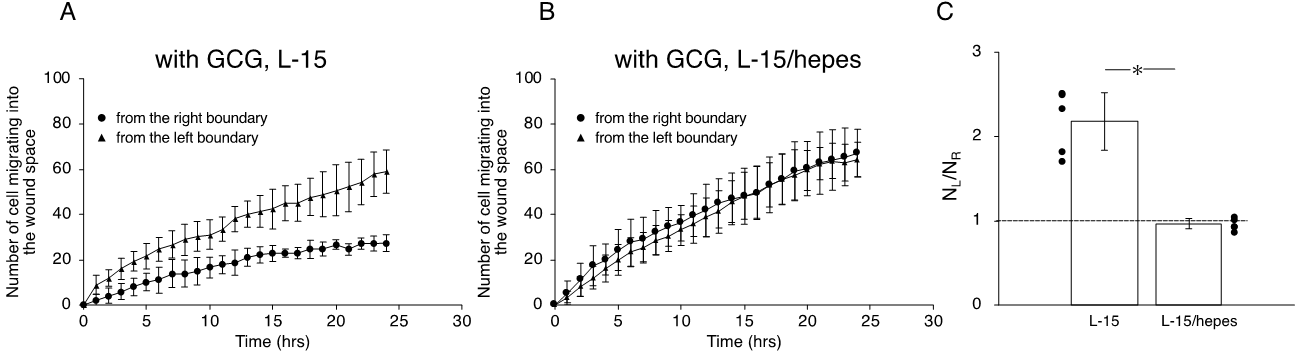
Figure 6. The number of cells that migrated into the wound space in 24 h. (A) In L-15 medium, NL and NR were different, consistent with what is observable in Figure 4B. (B) Conversely, in L-15/hepes medium, NL and NR were the same. (C) The heterogeneity in cell migration ultimately disappeared in L-15/hepes medium. Error bars represent the SD. Data were accumulated from five independent experiments. * p < 0.05, as judged by Student’s t-test.
3. Discussion
Directional migration of primary cancer cells toward intratumor blood/lymphatic vessels should elevate the probability for intravasation and ultimate hematogenous metastasis. On the analogy of chemotaxis, many presume that the gradients of nutrients and metabolic waste in the local tissue might guide tumor cells to nearby microvessels. However, at the present time, presence of such metabolic cues still remains an open question.
In the present study, we specifically focused on the gradients of H+ and oxygen as candidates for the metabolic cue. To monitor migratory behaviors of the cell in gradients of pH and/or oxygen in vitro, we previously proposed a simple microfluidic glassware, GCG, which is capable of producing gradients of energy substrates and metabolites, including H+ and oxygen in monolayer cultured cells [10]. Simultaneous changes in H+ and oxygen concentrations under the GCG are similar to those found in solid tumors and, therefore, experimentation using the GCG reflects a clinically relevant setting.
Previous in vitro studies demonstrated the possibility that oxygen concentration gradients may act as a guiding cue for cell migration. [13][14][15][16][17] Results of these in vitro studies appear conflicting in terms of the direction of cell migration in oxygen gradients. Indeed, effects of oxygen gradient on the cell migration should differ according to cell type, culture conditions (2D or 3D, culture media) and spatial profiles of oxygen concentration, including the magnitude and the local oxygen concentration at which cell is exposed. Furthermore, in most microfluidic devices, metabolic activities of the cell may also change spatial distribution of nutrients and metabolites that may affect the cellular migratory behavior in addition to the effect of the oxygen concentration gradient.
Initially, we hypothesized that the extracellular gradients of oxygen might be a cue for MDA-MB-231 cells to migrate directionally because steep gradients of oxygen concentration (~10 mmHg/100 µm) have been demonstrated in vivo [5]. Hypoxia-inducible factor 1α (HIF-1α) is an intracellular oxygen sensor that has been reported to affect the intracellular machinery for cell migration [18]. Therefore, it is likely that the HIF-1α pathway plays a role in directing cell migration in the steep gradient of oxygen concentration. However, in the present setting, the direct measurement of the oxygen concentration under the GCG achieved in the present study (Figure 2A) revealed oxygen gradients with relatively small magnitudes such that the oxygen concentration recorded at ~400 µm inside the GCG was much higher (~14%) than the oxygen level at which HIF-1α is responsive (5% or lower [19]). Thus, it is difficult to attribute the directional cell migration demonstrated in the present study specifically to HIF-1α dependent mechanisms. Although we do not exclude the possibility that oxygen gradients might direct cell migration in hypoxic microenvironment in tumor tissues in vivo, directional cell migration at unphysiologically high oxygen concentrations demonstrated in the present study prompted us to seek another possible cue for directional cell migration.
A few studies to date have addressed directional cell migration under gradients of extracellular pH in vitro. Paradise et al. [20] demonstrated using the Dunn chamber that both αVβ3 CHO-B2 cells and primary microvascular endothelial cells preferentially migrate toward acid in an extracellular pH gradient. In their research, the Dunn chamber produced a pH gradient of 6.0 to 7.5 over 1 mm. Elsewhere, Jagielska et al. [21] determined that oligodendrocyte precursor cells migrate toward areas of acidic extracellular pH produced by the Zigmond chamber. Here the pH gradient was set to 6.0 to 7.0 over a distance of 1 mm.
In the current study, we found that, first, a gradient of pH was in fact established in the extracellular medium under the GCG (0.2–0.3 units/mm); second, MDA-MB-231 cells under the GCG preferentially migrated toward the open-end of the GCG (i.e., higher pH/O2 regions); and third, such findings of directional cell migration completely disappeared when the extracellular pH gradient was abolished. Albeit, while gradients of various metabolic substances should exist under the GCG, these results strongly indicate that extracellular pH gradient is the predominant cue for the migration of MDA-MB-231 cells under the GCG.
Among migrating cells, gradients of intracellular and pericellular pH have been demonstrated at the level of the single cell. Martin et al. [22] observed intracellular pH gradients within single melanoma cells incubated in hepes-buffered Ringer solution at a pH of 7.0 and reported that the mean front-to-rear pHi gradients measured ~0.15 units over a 20-µm distance in migrating human (MV3) and murine (B16V) melanoma cells, where the front (leading edge) was more alkaline. Using a combination of pH-sensitive fluorescent dye and total internal reflection microscopy, Ludwig et al. [23] determined the pericellular pH on the surface of the plasma membrane in polarized MV3 cells, reporting significant gradients in single cells where front (at focal adhesions)-to-rear gradients were ~0.2 units (the cell front was more acidic). These subcellular heterogeneities in pH may arise from the heterogeneous distribution of the Na+/H+ exchanger isoform 1 (NHE1), a major plasma membrane protein that extrude protons from cytosol, where NHE1 accumulates in the leading edge of migrating cells [22][24][25]. Both intracellular and pericellular pHgradients may independently affect cell motility through effects on cytoskeletal machinery and cell-matrix interactions, respectively [26].
In the present experiment, we imposed 0.2–0.3 units/mm gradients of pH in the extracellular medium that correspond to a gradient of 0.02 units per single MDA-MB-231 cell. The value is far smaller, if compared at the single-cell level, than the NHE1-driven gradients of pH. This is also true in previous studies in which microfluidic devices produced ~1-unit/mm gradients of pH in the extracellular medium [20][21]. Therefore, it is unclear whether the relationship between cell migration and the NHE1-driven intracellular/pericellular heterogeneities would be directly applicable to the present and other experiments in which the pH of the extracellular bulk medium was manipulated.
In summary, the use of novel microfluidic devise GCG produced gradients of pH and oxygen concentration in the extracellular medium in monolayer MDA-MB-231 cells. We clearly demonstrated heterogeneous migration of the cells into the wound space in such a way that cells preferentially migrated in the direction of higher pH/oxygen concentration. Elimination of pH gradients also abolished the directional cell movement under the GCG thus indicating a possibility that extracellular pH gradients are the dominant guiding cue for migration of MDA-MB-231 in the present setting. Because, under GCG, extracellular oxygen concentration remained at unphysiologically high ranges despite the presence of significant gradients, the effect of oxygen concentration gradients on directional migration is still remain to be determined.
References
- Bockhorn, M.; Jain, R.K.; Munn, L.L. Active versus passive mechanisms in metastasis: Do cancer cells crawl into vessels, or are they pushed? Lancet Oncol. 2007, 8, 444–448.
- Elisabetta Marcuzzi; Roberta Angioni; Barbara Molon; B. Calì; Chemokines and Chemokine Receptors: Orchestrating Tumor Metastasization. International Journal of Molecular Sciences 2018, 20, 96, 10.3390/ijms20010096.
- Morgan O'hayre; Catherina L. Salanga; Tracy M. Handel; Samantha J. Allen; Chemokines and cancer: migration, intracellular signalling and intercellular communication in the microenvironment. Biochemical Journal 2008, 409, 635-649, 10.1042/bj20071493.
- Peter Vaupel; Tumor microenvironmental physiology and its implications for radiation oncology. Seminars in Radiation Oncology 2004, 14, 198-206, 10.1016/j.semradonc.2004.04.008.
- Gabriel Helmlinger; Fan Yuan; Marc Dellian; Rakesh K. Jain; Interstitial pH and pO2 gradients in solid tumors in vivo: High-resolution measurements reveal a lack of correlation. Nature Medicine 1997, 3, 177-182, 10.1038/nm0297-177.
- Franziska Van Zijl; Georg Krupitza; Wolfgang Mikulits; Initial steps of metastasis: Cell invasion and endothelial transmigration. Mutation Research/Reviews in Mutation Research 2011, 728, 23-34, 10.1016/j.mrrev.2011.05.002.
- Jennifer S. Fang; Robert D. Gillies; Robert A. Gatenby; Adaptation to hypoxia and acidosis in carcinogenesis and tumor progression. Seminars in Cancer Biology 2008, 18, 330-7, 10.1016/j.semcancer.2008.03.011.
- Calvin R. Justus; Edward J. Sanderlin; Li V. Yang; Molecular Connections between Cancer Cell Metabolism and the Tumor Microenvironment. International Journal of Molecular Sciences 2015, 16, 11055-11086, 10.3390/ijms160511055.
- Madeleine J. Oudin; Valerie M. Weaver; Physical and Chemical Gradients in the Tumor Microenvironment Regulate Tumor Cell Invasion, Migration, and Metastasis. Cold Spring Harbor Symposia on Quantitative Biology 2016, 81, 189-205, 10.1101/sqb.2016.81.030817.
- D. Yahara; T. Yoshida; Y. Enokida; Eiji Takahashi; Directional Migration of MDA-MB-231 Cells Under Oxygen Concentration Gradients. Single Molecule and Single Cell Sequencing 2016, 923, 129-134, 10.1007/978-3-319-38810-6_17.
- Y. Enokida; Y. Tsuruno; K. Okubo; Y. Yamaoka; Eiji Takahashi; Directional Migration of MDA-MB-231 Cells Under O2/pH Gradients. Advances in Experimental Medicine and Biology 2017, 977, 169-174, 10.1007/978-3-319-55231-6_23.
- Yusuke Tsuruno; Kaima Okubo; Takahiro Fujiwara; Yoshihisa Yamaoka; Eiji Takahashi; An In Vitro Model for Determining Tumor Cell Migration Under Metabolic Gradients. Advances in Experimental Medicine and Biology 2018, 1072, 201-205, 10.1007/978-3-319-91287-5_32.
- Chia-Wen Chang; Yung-Ju Cheng; Melissa Tu; Ying-Hua Chen; Chien-Chung Peng; Wei-Hao Liao; Yi-Chung Tung; A polydimethylsiloxane–polycarbonate hybrid microfluidic device capable of generating perpendicular chemical and oxygen gradients for cell culture studies. Lab on a Chip 2014, 14, 3762-3772, 10.1039/c4lc00732h.
- Hsiu-Chen Shih; Tse-Ang Lee; Hsiao-Mei Wu; Ping-Liang Ko; Wei-Hao Liao; Yi-Chung Tung; Microfluidic Collective Cell Migration Assay for Study of Endothelial Cell Proliferation and Migration under Combinations of Oxygen Gradients, Tensions, and Drug Treatments.. Scientific Reports 2019, 9, 8234, 10.1038/s41598-019-44594-5.
- Jelle J. F. Sleeboom; Jaap M.J. Den Toonder; Cecilia M. Sahlgren; MDA-MB-231 Breast Cancer Cells and Their CSC Population Migrate Towards Low Oxygen in a Microfluidic Gradient Device. International Journal of Molecular Sciences 2018, 19, 3047, 10.3390/ijms19103047.
- Bobak Mosadegh; Matthew R. Lockett; Kyaw Thu Minn; Karen A. Simon; Karl Gilbert; Shawn Hillier; David Newsome; Howard Li; Amy B. Hall; Diane M. Boucher; Brenda K. Eustace; George M. Whitesides; A paper-based invasion assay: Assessing chemotaxis of cancer cells in gradients of oxygen. Biomaterials 2015, 52, 262-271, 10.1016/j.biomaterials.2015.02.012.
- Daniel M. Lewis; Kyung Min Park; Vitor Tang; Yu Xu; Koreana Pak; T. S. Karin Eisinger-Mathason; M. Celeste Simon; Sharon Gerecht; Intratumoral oxygen gradients mediate sarcoma cell invasion. Proceedings of the National Academy of Sciences 2016, 113, 9292-9297, 10.1073/pnas.1605317113.
- Daniele M. Gilkes; Lisha Xiang; Sun Joo Lee; Pallavi Chaturvedi; Maimon E. Hubbi; Denis Wirtz; Gregg L. Semenza; Hypoxia-inducible factors mediate coordinated RhoA-ROCK1 expression and signaling in breast cancer cells. Proceedings of the National Academy of Sciences 2013, 111, E384-E393, 10.1073/pnas.1321510111.
- B H Jiang; G L Semenza; C Bauer; H H Marti; Hypoxia-inducible factor 1 levels vary exponentially over a physiologically relevant range of O2 tension. American Journal of Physiology-Legacy Content 1996, 271, C1172-1180.
- Ranjani K. Paradise; Matthew J. Whitfield; Uglas A. Lauffenburger; Krystyn J. Van Vliet; Directional cell migration in an extracellular pH gradient: A model study with an engineered cell line and primary microvascular endothelial cells. Experimental Cell Research 2013, 319, 487-497, 10.1016/j.yexcr.2012.11.006.
- Anna Jagielska; Kristen D. Wilhite; Krystyn J. Van Vliet; Extracellular Acidic pH Inhibits Oligodendrocyte Precursor Viability, Migration, and Differentiation. PLOS ONE 2013, 8, e76048, 10.1371/journal.pone.0076048.
- Martin, C.; Pedersen, S.F.; Schwab, A.; Stock, C; Intracellular pH gradients in migrating cells. Am. J. Physiol. Cell Physiol. 2011, 300, C490-495.
- Florian Timo Ludwig; Albrecht Schwab; Christian Stock; The Na+/H+-exchanger (NHE1) generates pH nanodomains at focal adhesions. Journal of Cellular Physiology 2013, 228, 1351-1358, 10.1002/jcp.24293.
- Sheryl P. Denker; Diane L. Barber; Cell migration requires both ion translocation and cytoskeletal anchoring by the Na-H exchanger NHE1. Journal of Cell Biology 2002, 159, 1087-1096, 10.1083/jcb.200208050.
- Helene Halkjær Jensen; Gitte A. Pedersen; Jeanette J. Morgen; Maddy Parsons; Stine F. Pedersen; Lene Niemann Nejsum; The Na + /H + exchanger NHE1 localizes as clusters to cryptic lamellipodia and accelerates collective epithelial cell migration. The Journal of Physiology 2019, 597, 849-867, 10.1113/jp277383.
- Stock, C.; Pedersen, S.F. Roles of pH and the Na+/H+ exchanger NHE1 in cancer: From cell biology and animal models to an emerging translational perspective? Semin. Cancer Biol. 2017, 43, 5–16.

