 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Hasan Mehraj | + 1014 word(s) | 1014 | 2020-06-16 10:00:02 | | | |
| 2 | Catherine Yang | + 71 word(s) | 1085 | 2021-01-13 04:00:09 | | |
Video Upload Options
Clubroot is a major disease for the brassica crops, and it can not completely control by any other means except the resistant caultivars. MAS breeding helps us for the development of new resistant cultivars
1. Introduction
MAS is an indirect selection process where a trait of interest is selected based on a marker (morphological, biochemical, or DNA/RNA variation) linked to that trait. Selecting individuals with disease resistance using MAS involves identifying a marker allele that is linked to disease resistance rather than to the level of disease resistance. There are several types of DNA markers that have been used to identify disease resistance genes [1][2][3][4][5][6][7][8].
The complexity of plant–pathogen interaction is a problematic in the case of CR breeding due to the appearance of multiple races of the pathogen [9]. Combinations of different CR genes exhibit higher resistance to the disease [10][11][12]. Though CR cultivars have been used widely for major production areas, field isolates of P. brassicae show variation, and different resistance sources from either B. rapa or B. oleracea vegetables were attained by P. brassicae. This suggests a serious risk that a resistance gene can be overcome by pathogen variants [13]. For example, seven CR canola cultivars were characterized for virulence in 106 P. brassicae population, and 61 of 106 P. brassicae population overcame the resistance in at least one of the seven CR cultivars [14]. There are many reports that CR genes show different reactions against the variable virulence of P. brassicae [15][16][17][18][19][10][20][21][22], but heterozygous CR loci are less resistant than the homozygous state [23]. B. rapa possesses several major CR loci, which may confer differential (pathotype-specific) resistance to particular isolates of P. brassicae, and sometimes this may have a large effect on resistance [9][15][24][25]. The NARO Institute of Vegetable and Tea Science (NIVTS) has developed a high CR Chinese cabbage cultivar, ‘Akimeki’, by the accumulation of Crr1, Crr2, and CRb genes. It was proven that the accumulation of CR genes through MAS strengthened resistance and, consequently, it can be resistant to the multiple races of P. brassicae in B. rapa. Three CR genes, CRa, CRk, and CRc, were accumulated in Chinese cabbage through MAS [9] and the homozygous lines for the CR genes exhibited exceedingly high resistance against all six field isolates of P. brassicae. The effect of accumulation of different CR genes could be controlled by the dose-dependent accumulation of CR proteins [26][27]. In B. oleracea, resistance in genotypes has generally been identified less frequently than in the genotypes of B. rapa and the level of resistance is low [28]. This might be due to the polygenic nature of resistance in B. oleracea [11]. B. oleracea progeny were developed by accumulating major and minor QTLs to evaluate its effectiveness to the clubroot disease [29]. Three QTLs in the F2/F3 population from the cross between cabbage and kale line K269 were identified [10]. The accumulation of those three CR genes showed broad resistance to three isolates. It was observed that only one major QTL PbBo(Anju)1 showed moderate resistance, whereas three minor QTLs without the major one showed distinct susceptibility [29]. Later, it was proven that PbBo(Anju)1 and three minor QTLs PbBo(Anju)2, PbBo(Anju)4, and PbBo(GC)1 play a critical role in the acquisition of resistance to clubroot disease [11][12]. Here, PbBo(Anju)1 plays a crucial role in the expression of clubroot resistance, and pyramiding minor CR genes are also essential for achieving higher resistance [11][12]. Their effectiveness was verified for controlling disease involving various isolates of P. brassicae [11]. Recently, two CR genes, CRb and PbBa8.1, were combined through MAS and CR homozygous lines in developed B. napus. The homozygous lines demonstrated a higher resistance than the heterozygous lines [30].
The Type A resistance to Fusarium wilt disease controlled by a single dominant gene has been successfully mapped and molecular markers have been developed: SSR marker KBrS003O1N10 [31], InDel markers M10 and A1 [6], Indel markers Bra012688m and Bra012689m [32][7], and DNA marker sets [8][33][34], which are used to generate a series of resistance cultivars (Figure 1).
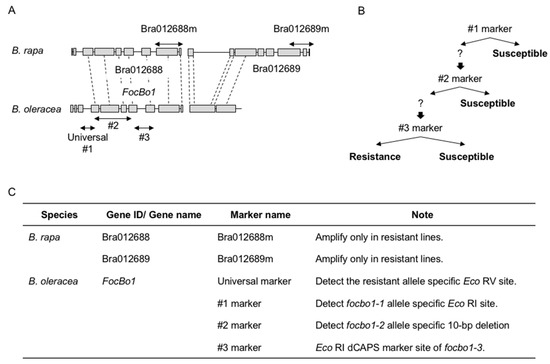
Figure 1. Schematic view of the alignment of resistance genes of Fusarium wilt disease. (A). DNA marker positions of resistance genes in B. rapa and B. oleracea. Arrows indicate marker positions. (B). Scheme of marker assisted selection in B. oleracea. (C). DNA marker list for marker assisted selection in B. rapa and B. oleracea.
2. Influence and Results
Breeding cultivars that have resistance to both clubroot and Fusarium wilt is desired. However, inoculation tests against multiple pathogens or multiple races are difficult to perform on the same individual plant. Thus, DNA marker-based selection is useful for the identification of plants that have one Fusarium wilt resistance gene and multiple clubroot resistance genes. Furthermore, it is necessary to confirm whether these resistance genes are linked. In B. rapa, a Fusarium wilt resistance gene is located on chromosome 3, and CRa/CRb, Crr3, and CRk are located near this Fusarium wilt resistance gene. The CRa/CRb gene is the closest, being approximately 2 Mb in physical distance to the Fusarium wilt resistance gene (Figure 2). Since recombination between these two genes can occur [7], it is possible to inherit both resistance genes. In B. oleracea, a Fusarium wilt resistance gene is located on chromosome 7, and there is a minor QTL for clubroot resistance, PbBo(Anju)4, nearby this Fusarium wilt resistance gene. However, these loci are not completely linked to each other [8][32][35]. Therefore, it is possible to have both resistance genes. In B. napus, the association between susceptibility to Fusarium wilt and clubroot resistance against pathotype 3 was found, and these two resistance genes are located about 10 cM apart [36]. However, recombination between these two genes has been reported [36], suggesting that it is possible to inherit both resistance genes and identify them by DNA marker-based selection.
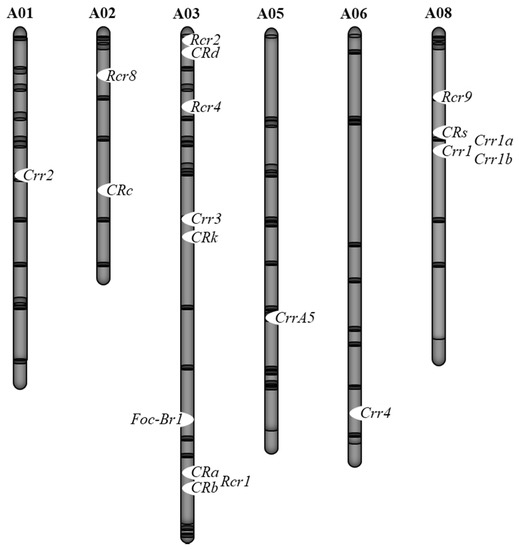
Figure 2. Chromosomal locations of clubroot resistant (CR) and Fusarium wilt resistant loci in B. rapa.
From the results from various researchers, it has been demonstrated that the DNA markers developed can select for the genes that are required for the acquisition of resistance, and these markers could be a powerful tool for resistance breeding in Brassica species. The novel breeding method developed can reinforce resistance by pyramiding R genes through MAS. For the genetic accumulation of R genes corresponding to wide pathogenicity, MAS is indispensable because it allows a precise identification of how many R genes are involved in a cultivar, and can monitor the accumulation of R genes in the progeny in the breeding program. To increase the durability of resistant cultivars to a broader spectrum of pathogen races, the combination of different R genes into a single line will be indispensable.
References
- Lv, H.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Q.; Liu, Y.; Zhuang, M.; Yang, Y.; Xie, B.; Liu, B.; et al. Mapping and analysis of a novel candidate Fusarium wilt resistance gene FOC1 in Brassica oleracea. BMC Genom. 2014, 15, 1094.
- Piao, Z.Y.; Deng, Y.Q.; Choi, S.R.; Park, Y.J.; Lim, Y.P. SCAR and CAPS mapping of CRb, a gene conferring resistance to Plasmodiophora brassicae in Chinese cabbage (Brassica rapa ssp. pekinensis). Theor. Appl. Genet. 2004, 108, 1458–1465.
- Kato, T.; Hatakeyama, K.; Fukino, N.; Matsumoto, S. Identification of a clubroot resistance locus conferring resistance to a Plasmodiophora brassicae classified into pathotype group 3 in Chinese cabbage (Brassica rapa L.). Breed. Sci. 2012, 62, 282–287.
- Zhang, T.; Zhao, Z.; Zhang, C.; Pang, W.; Choi, S.R.; Lim, Y.P.; Piao, Z. Fine genetic and physical mapping of CRb gene conferring resistance to clubroot disease in Brassica rapa. Mol. Breed. 2014, 34, 1173–1183.
- Grandclemant, C.; Laurens, F.; Thomas, G. Genetic analysis of resistance to clubroot (Plasmodiophora brassicae Woron) in two Brassica oleracea groups (spp. acephala and spp. botrytis) through diallel analysis. Plant Breed. 1996, 115, 152–156.
- Lv, H.; Yang, L.; Kang, J.; Wang, Q.; Wang, X.; Fang, Z.; Liu, Y.; Zhuang, M.; Zhang, Y.; Lin, Y.; et al. Development of InDel markers linked to Fusarium wilt resistance in cabbage. Mol. Breed. 2013, 32, 961–967.
- Kawamura, K.; Kawanabe, T.; Shimizu, M.; Nagano, A.J.; Saeki, N.; Okazaki, K.; Kaji, M.; Dennis, E.S.; Osabe, K.; Fujimoto, R. Genetic characterization of inbred lines of Chinese cabbage by DNA markers; towards the application of DNA markers to breeding of F1 hybrid cultivars. Data Brief 2016, 6, 229–237.
- Kawamura, K.; Shimizu, M.; Kawanabe, T.; Pu, Z.; Kodama, T.; Kaji, M.; Kenji, O.; Fujimoto, R.; Keiichi, O. Assessment of DNA markers for seed contamination testing and selection of disease resistance in cabbage. Euphytica 2017, 213, 28.
- Matsumoto, E.; Ueno, H.; Aruga, D.; Sakamoto, K.; Hayashida, N. Accumulation of three clubroot resistance genes through marker-assisted selection in Chinese cabbage (Brassica rapa spp. pekinensis). J. Jpn. Soc. Hortic. Sci. 2012, 81, 184–190.
- Nomura, K.; Minegishi, Y.; Kimizuka-Takagi, C.; Fujioka, T.; Moriguchi, K.; Shishido, R.; Ikehashi, H. Evaluation of F2 and F3 plants introgressed with QTLs for clubroot resistance in cabbage developed by using SCAR markers. Plant Breed. 2005, 124, 371–375.
- Tomita, H.; Shimizu, M.; Doullah, M.A.U.; Fujimoto, R.; Okazaki, K. Accumulation of quantitative trait loci conferring broad-spectrum clubroot resistance in Brassica oleracea. Mol. Breed. 2013, 32, 889–900.
- Doullah, M.A.U.; Tomita, H.; Shimizu, M.; Matsumoto, S.; Fujimoto, R.; Okazaki, K. Recent progress of clubroot resistance breeding through marker assisted selection in Brassica rapa and Brassica oleracea. J. Sylhet Agric. Univ. 2014, 1, 139–146.
- Neik, T.X.; Barbetti, M.J.; Batley, J. Current status and challenges in identifying disease resistance genes in Brassica napus. Front. Plant Sci. 2017, 8, 1788.
- Strelkov, S.E.; Hwang, S.F.; Manolii, V.P.; Cao, T.; Fredua-Agyeman, R.; Harding, M.W.; Peng, G.; Gossen, B.D.; Mcdonald, M.R.; Feindel, D. Virulence and pathotype classification of Plasmodiophora brassicae populations collected from clubroot resistant canola (Brassica napus) in Canada. Can. J. Plant Pathol. 2018, 40, 284–298.
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Matsumoto, S.; Hirai, M. Identification of two loci for resistance to clubroot (Plasmodiophora brassicae Woronin) in Brassica rapa L. Theor. Appl. Genet. 2003, 107, 997–1002.
- Yu, F.; Zhang, X.; Peng, G.; Falk, K.C.; Strelkov, S.E.; Gossen, B.D. Genotyping-by-sequencing reveals three QTL for clubroot resistance to six pathotypes of Plasmodiophora brassicae in Brassica rapa. Sci. Rep. 2017, 7, 4516.
- Chen, J.; Jing, J.; Zhan, Z.; Zhang, T.; Zhang, C.; Piao, Z. Identification of novel QTLs for isolate-specific partial resistance to Plasmodiophora brassicae in Brassica rapa. PLoS ONE 2013, 8, e85307.
- Huang, Z.; Peng, G.; Liu, X.; Deora, A.; Falk, K.C.; Gossen, B.D.; McDonald, M.R.; Yu, F. Fine mapping of a clubroot resistance gene in Chinese cabbage using SNP markers identified from bulked segregant RNA Sequencing. Front. Plant Sci. 2017, 8, 1448.
- Moriguchi, K.; Kimizuka-Takagi, C.; Ishii, K.; Nomura, K. A genetic map based on RAPD, RFLP, isozyme, morphological markers and QTL analysis for clubroot resistance in Brassica oleracea. Breed. Sci. 1999, 49, 257–265.
- Rocherieux, J.; Glory, P.; Giboulot, A.; Boury, S.; Barbeyron, G.; Thomas, G.; Manzanares-Dauleux, M.J. Isolate-specific and broad-spectrum QTLs are involved in the control of clubroot in Brassica oleracea. Theor. Appl. Genet. 2004, 108, 1555–1563.
- Peng, L.; Zhou, L.; Li, Q.; Wei, D.; Ren, X.; Song, H.; Mei, J.; Si, J.; Qian, W. Identification of quantitative trait loci for clubroot resistance in Brassica oleracea with the use of Brassica SNP microarray. Front. Plant Sci. 2018, 9, 822.
- Hasan, M.J.; Rahman, R. Genetics and molecular mapping of resistance to Plasmodiophora brassicae pathotypes 2, 3, 5, 6 and 8 in rutabaga (Brassica napus var. napobrassica). Genome 2016, 59, 805–815.
- Hirai, M. Genetic analysis of clubroot resistance in Brassica crops. Breed. Sci. 2006, 56, 223–229.
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Kondo, M.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Hirai, M.; Matsumoto, S. Simple sequence repeat-based comparative genomic between Brassica rapa and Arabidopsis thaliana: The genetic origin of clubroot resistance. Genetics 2006, 173, 309–319.
- Kuginuki, Y.; Yoshikawa, H.; Hirai, M. Variation in virulence of Plasmodiophora brassicae in Japan tested with clubroot-resistant cultivars of Chinese cabbage (Brassica rapa L. spp. pekinensis). Eur. J. Plant Pathol. 1999, 105, 327–332.
- Hatakeyama, K.; Suwabe, K.; Tomita, R.N.; Kato, T.; Nunome, T.; Fukuoka, H.; Matsumoto, S. Identification and characterization of Crr1a, a gene for resistance to clubroot disease (Plasmodiophora brassicae Woronin) in Brassica rapa L. PLoS ONE 2013, 8, e54745.
- Kou, Y.; Wang, S. Broad-spectrum and durability: Understanding of quantitative disease resistance. Curr. Opin. Plant Biol. 2010, 13, 181–185.
- Crisp, P.; Crute, I.R.; Sutherland, R.A.; Angell, S.M.; Bloor, K.; Burgess, H.; Gordon, P.L. The exploitation of genetic resources of Brassica oleracea in breeding for resistance to clubroot (Plasmodiophora brassicae). Euphytica 1889, 42, 215–226.
- Nagaoka, T.; Doullah, M.A.U.; Matsumoto, S.; Kawasaki, S.; Ishikawa, T.; Hori, H.; Okazaki, K. Identification of QTLs that control clubroot resistance in Brassica oleracea and comparative analysis of clubroot resistance genes between B. rapa and B. oleracea. Theor. Appl. Genet. 2010, 120, 1335–1346.
- Shah, N.; Sun, J.; Yu, S.; Yang, Z.; Wang, Z.; Huang, F.; Dun, B.; Gong, J.; Liu, Y.; Li, Y.; et al. Genetic variation analysis of field isolates of clubroot and their responses to Brassica napus lines containing resistant genes CRb and PbBa8.1 and their combination in homozygous and heterozygous state. Mol. Breed. 2019, 39, 153.
- Pu, Z.; Shimizu, M.; Zhang, Y.; Nagaoka, T.; Hayashi, T.; Hori, H.; Matsumoto, S.; Fujimoto, R.; Okazai, K. Genetic mapping of a Fusarium wilt resistance gene in Brassica oleracea. Mol. Breed. 2012, 30, 809–818.
- Shimizu, M.; Fujimoto, R.; Ying, H.; Pu, Z.; Ebe, Y.; Kawanabe, T.; Saeki, N.; Taylor, J.M.; Kaji, M.; Dennis, E.S.; et al. Identification of candidate genes for Fusarium yellows resistance in Chinese cabbage by differential expression analysis. Plant Mol. Biol. 2014, 85, 247–257.
- Shimizu, M.; Pu, Z.; Kawanabe, T.; Kitashiba, H.; Matsumoto, S.; Ebe, Y.; Sano, M.; Funaki, E.; Fujimoto, R.; Okazai, K. Map-based cloning of a candidate gene conferring Fusarium yellows resistance in Brassica oleracea. Theor. Appl. Genet. 2015, 128, 119–130.
- Sato, M.; Shimizu, M.; Shea, D.J.; Hoque, M.; Kawanabe, T.; Miyaji, N.; Fujimoto, R.; Fukai, E.; Okazaki, K. Allele specific DNA marker for fusarium resistance gene FocBo1 in Brassica oleracea. Breed. Sci. 2019, 69, 308–315.
- Pu, Z.; Ino, Y.; Kimura, Y.; Tago, A.; Shimizu, M.; Natsume, S.; Sano, Y.; Fujjimoto, R.; Kaneko, K.; Shea, D.J.; et al. Changes in the proteome of xylem sap in Brassica oleracea in response to Fusarium oxysporum stress. Front. Plant Sci. 2016, 7, 31.
- Rahman, H.; Franke, C. Association of fusarium wilt susceptibility with clubroot resistance derived from winter Brassica napus L. ‘Mendel’. Can. J. Plant Pathol. 2019, 41, 60–64.


