Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Hans V. Westerhoff | -- | 4801 | 2024-01-25 21:42:53 | | | |
| 2 | Rita Xu | -1266 word(s) | 3535 | 2024-01-26 02:37:27 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Hassan, Z.; Westerhoff, H.V. Arsenic Contamination of Groundwater. Encyclopedia. Available online: https://encyclopedia.pub/entry/54365 (accessed on 16 July 2026).
Hassan Z, Westerhoff HV. Arsenic Contamination of Groundwater. Encyclopedia. Available at: https://encyclopedia.pub/entry/54365. Accessed July 16, 2026.
Hassan, Zahid, Hans V. Westerhoff. "Arsenic Contamination of Groundwater" Encyclopedia, https://encyclopedia.pub/entry/54365 (accessed July 16, 2026).
Hassan, Z., & Westerhoff, H.V. (2024, January 25). Arsenic Contamination of Groundwater. In Encyclopedia. https://encyclopedia.pub/entry/54365
Hassan, Zahid and Hans V. Westerhoff. "Arsenic Contamination of Groundwater." Encyclopedia. Web. 25 January, 2024.
Copy Citation
At a great many locations worldwide, the safety of drinking water is endangered by pollution with arsenic. Arsenic toxicity is a matter of both systems chemistry and systems biology: it is determined by complex and intertwined networks of chemical reactions in the inanimate environment, in microbes in that environment, and in the human body.
systems biology
systems chemistry
arsenic toxicity
subsurface arsenic removal
arsenic microbial ecology
1. Introduction
Chemical pollution of groundwater poses a serious threat to public health. The pollution can either be due to industrial discharges and other anthropogenic activities or it can occur naturally. Arsenic is known as the “king of poison” or “hidden killer” [1]. The leaching of soil, weathering of rocks, and agricultural runoff all introduce arsenic into groundwater [2]. Natural sources further include seawater, arsenic-bearing minerals, volcanic emissions, and rivers originating in the Himalayas [3][4].
Inorganic arsenic is naturally present at high levels in groundwater in many countries across the globe, including the Americas (e.g., Argentina, Chile, Mexico, USA) [5], Asia (e.g., Bangladesh, India, Pakistan, Nepal, China, Taiwan), Southeast Asia (e.g., Indonesia, Thailand, Vietnam, Cambodia, Myanmar) [5][6][7][8][9][10][11][12][13], and Europe (e.g., Hungary, France, Germany, Romania, Italy) [14][15][16][17][18] (Figure 1). Nearly 200 million people worldwide are at risk of arsenic poisoning, including 180 million in Asia [19][20][21]. In nearly 108 countries, arsenic in groundwater exceeds the maximum of 10 µg/L recommended by the WHO. In total, 1 person out of every 60 people lives in a region where the concentration of arsenic in groundwater is 50 µg/L or above [22]. Bangladesh is the country that is worst off (Figure 1): groundwater is its major source of pathogen-free drinking water [23][24]. Ineffective water purification and sewage systems as well as periodic monsoons, cyclones, flooding, drought, and salinity complicate access to reliable drinking water. Approximately, 80 million inhabitants of Bangladesh are exposed to groundwater with concentrations above 50 µg/L and 35 million are potentially exposed to even higher concentrations of arsenic (50–300 μg/L) in drinking water [25][26]. The arsenic concentration in some tube wells is as high as 4.7 mg/L [27]. The WHO called this the largest mass poisoning of a population in history [28] yet increased its guideline for maximal arsenic in drinking water in Bangladesh to 50 μg/L [29][30]. These numbers are extreme for this country, but many other countries are also troubled by arsenic in groundwater [8].
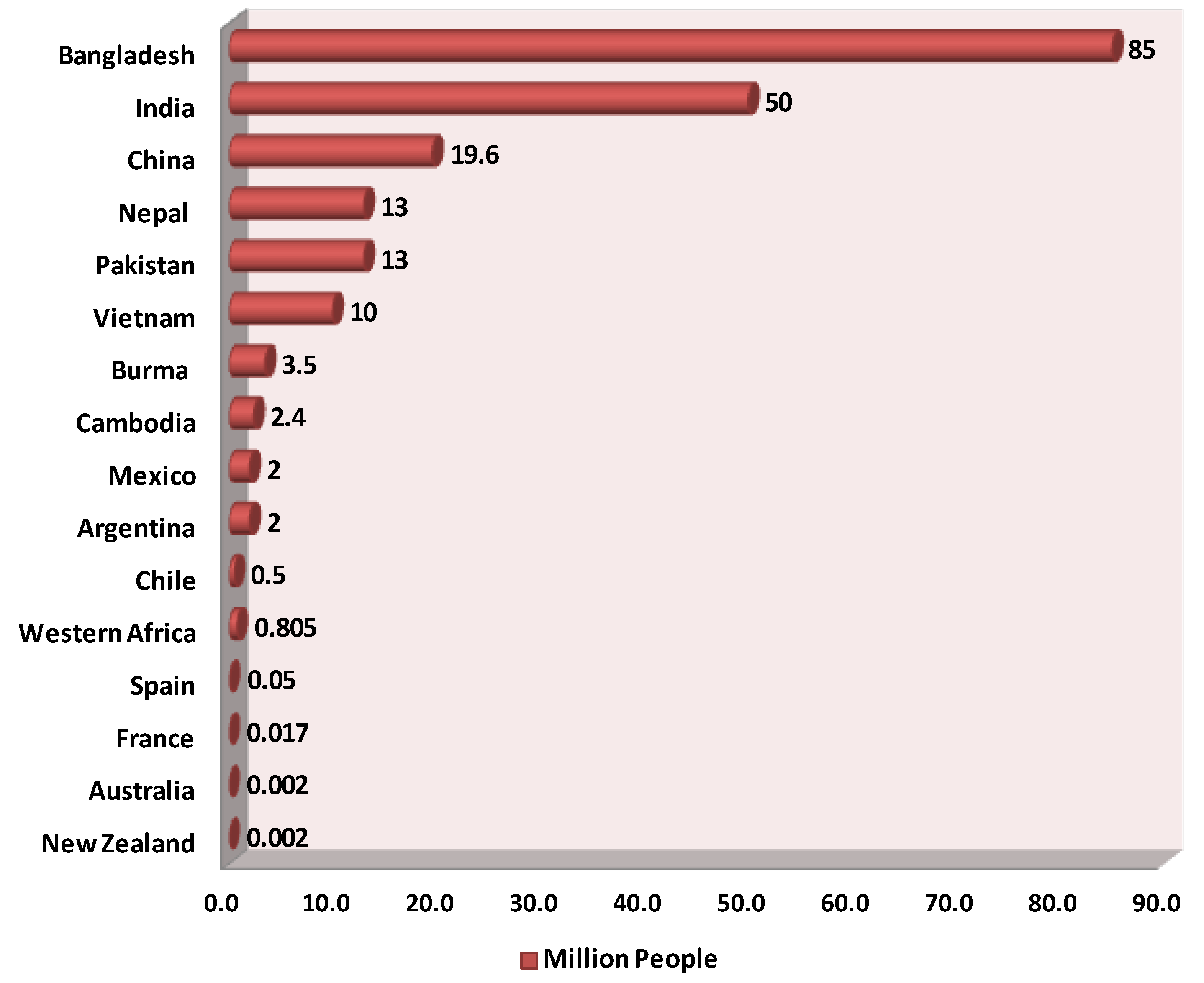
Figure 1. Estimated numbers of people (in millions) per country exposed to arsenic-contaminated groundwater.
The toxicity of arsenic in groundwater depends on a multitude of factors. These include chemical aspects like the conversion of the arsenic between its multiple chemical forms; the precipitation and subsequent adsorption of these forms with various metal oxides, particularly of manganese, iron, or aluminium [31]; the interaction of different arsenic species with various chemical (redox) forms of iron; the biochemical reactions of the arsenicals in the human; the interactions of arsenicals with metabolism; various anthropogenic activities that occur through mining and subsequent chemical modifications that occur during smelting for industrial, pharmaceutical, and agricultural uses; and various other groundwater characteristics such as ambient redox potential and pH, as well as nitrate, organic carbon, sulfur, and many other chemical concentrations [15].
2. The Water Chemistry of As and Fe Connects to Microbiology: Biology Does Matter
At atmospheric oxygen pressure, the oxidation of aqueous ferrous iron to ferric iron and the precipitation of the latter occur at substantial rates in the absence of living matter. We know this as rust formation. At consequently reduced dissolved oxygen concentrations in groundwater, the rate of these abiotic processes is reduced proportionally however. The accompanying paper (Hassan and Westerhoff (2024) reviews much of the thermodynamics and systems chemistry before turning to the roles that living organisms play. In the environment, microorganisms (i.e., bacteria, archaea or fungi) catalyze these processes, which strongly increases their rates and extents. Some microorganisms can harvest (Gibbs) energy from the oxidation of ferrous iron, which means that they have energy to grow. Provided that other elements required for their growth (like nitrogen and phosphorous) are present, these microorganisms can amplify to high abundance so that the iron oxidation at reduced oxygen levels again becomes substantial until the oxygen activity becomes really low. Other microorganisms can similarly oxidize arsenite to arsenate, reduce arsenate back to arsenite, reduce ferric iron to ferrous iron, or alter ambient redox potential (oxygen concentration) and pH in processes that are all very slow in the absence of microorganisms. Thus, arsenic and iron chemistry in natural waters is not determined just by iron and arsenic chemistry itself but also to a considerable extent by the microbial activities that occur and develop in those waters, i.e., by the Biology.
Conversely, the abundance of the microorganisms depends on the availability of the various forms of iron and arsenic, first as sources of Gibbs energy (e.g., through the oxidation of ferrous iron or arsenic by molecular oxygen catalyzed by these organisms) for their growth and, second, in terms of the toxicity of arsenite and arsenate, which causes death or growth inhibition. Precipitated ferric oxides and associated arsenic species may further enable microorganisms to attach and profit from a stable source of these materials or from the possibility to deposit toxic compounds outside their cells onto such material.
2.1. Organic Matter
The co-occurrence of very high concentrations of dissolved organic matter (DOM) with elevated concentrations of dissolved arsenic and iron in 'reductive' groundwater has often been observed [32]. DOM significantly influences arsenic biogeochemistry, and reactive organic matter facilitates the microbial release of arsenic from sediment to groundwater [33]. Sedimentary organic matter may provide carbon sources for microorganisms. Moreover, shallow groundwater is usually recharged by surface water, importing reactive organic carbon and accelerating microbial processes. Autotrophic microbial growth (i.e. growth that uses inorganic CO2 from the atmosphere to synthesize its organic carbon) may further increase the organic matter density. Such changes in the organic matter potentially influence both the spatial and the temporal evolution of groundwater arsenic geochemistry.
2.2. Microbiology of Arsenic: What Can Microorganisms Do?
Microorganisms cannot perform miracles. What they do must be consistent with thermodynamics, i.e., the processes that they catalyze must run downhill in terms of Gibbs energy. They can, however, escape from this limitation by coupling a thermodynamically uphill reaction to a different, thermodynamically downhill reaction. The most important example is the coupling of the often thermodynamically uphill reaction of microbial growth [34] to a process delivering Gibbs energy. The latter process is photon absorption in photoautotrophic microorganisms (which use light absorption for their input energy), the catabolism of organic material in heterotrophs, and inorganic reactions such as arsenite or ferrous iron oxidation by oxygen or nitrate in lithoautotrophs. This example is most important because microbial growth leads to the autoamplification of the chemical activities. The catalyst of the process, i.e., the microorganism, can thereby become tremendously active. Limitations here are the time it takes for the microbes to replicate and the other chemicals they require for growth. The elements carbon, nitrogen, phosphorous, and sulfur are minimally required, and this is an important issue as some ecosystems are lacking in one or more of these and, in other ecosystems, competing microorganisms strongly reduce their levels. It is an important observation that the elements and Gibbs energies may be limiting, but not the catalytic activities; the microbial geosphere is rich and dispersed enough to catalyze virtually anything that is possible in terms of thermodynamics and element conservation. And it will augment itself through proliferation.
Accordingly, microbial communities drive much of the global biogeochemical cycling of arsenic, and they do this through diverse metabolic functions [31][35][36]. In addition to Gibbs energy harvesting and coupling, microorganisms have evolved a variety of mechanisms to overcome the effect of metal(loid) toxicity. These include (i) mechanisms that restrict arsenic entry into the cell or enable active extrusion, (ii) enzymatic detoxification through redox transformations, and (iii) chelation or precipitation [37][38][39]. Frequently metal(loid) resistance genes are located on mobile genetic elements and are readily transferred between different bacteria via horizontal gene transfer [40]. Taxonomically diverse bacterial populations viz. Alpha-, Beta-, Gamma-proteobacteria, Firmicutes (Bacillus and relatives), Actinobacteria, Bacteroidetes, etc., play roles in the bio-geochemistry of arsenic-rich groundwater [41][42][43][44][45][46][47][48]. Chemolithotrophic and heterotrophic arsenic-transforming bacteria deploy an array of metabolic routes in arsenic speciation, distribution, and cycling in aquatic systems [35][49][50][51]. Diverse microbial genes encode metabolic processes involved in arsenic-oxidation, -reduction, and -methylation [52][53][54][55] and thereby affect arsenic speciation, mobilization, and availability as well as ecotoxicity [56][57]. Levels of mobile arsenic in groundwater depend on the balance between all the biochemical processes mentioned. These therefore need to be evaluated in any particular case of arsenic contamination in groundwater.
Microorganisms contribute to arsenic removal, in particular, ferrous iron oxidizers and arsenite oxidizers.
2.3. Microbial Oxidation of Ferrous Iron
Ferrous iron [Fe(II)] is an electron donor to a wide range of iron-oxidizing bacteria, and such iron oxidation can be operated at both acidic and neutral extracellular pHs, under either oxic or anoxic conditions (Figure 2). Fortin et al. noted that microbial iron oxidation is accelerated through a variety of mechanisms [58]. At pH = 4, the apparent negative midpoint potential of the oxygen–water couple amounts to −E0′ = −1.0 V, i.e., 0.23 V lower than that of the ferrous/aqueous ferric iron couple (−E0′ = −0.77). This implies that respiration with molecular oxygen as an electron acceptor, ferrous iron as an electron donor, and aqueous (i.e., soluble) ferric iron as a product is thermodynamically feasible in environments with an acidic pH, i.e., for acidophilic organisms; precipitation into goethite or hematite is not required for these energetics. Thus, ferrous iron constitutes a good source of Gibbs free energy for aerobic acidophilic prokaryotes [59][60]. Acidithiobacillus spp., (β-Proteobacteria) are by far the most studied group of bacteria capable of gaining Gibbs energy from the oxidation of ferrous to ferric iron at a very low pH.
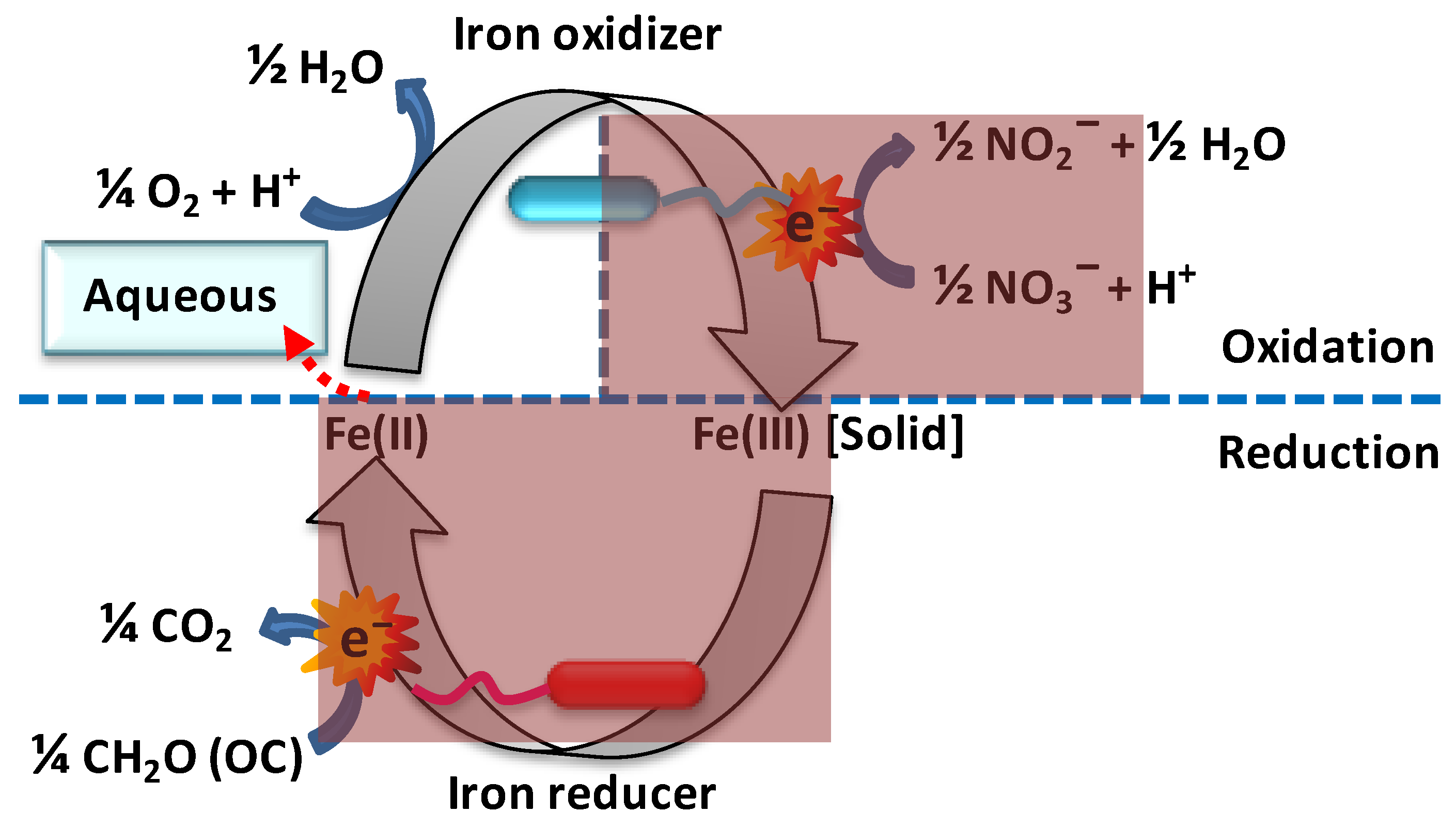
Figure 2. Microbial redox cycling of iron. In microaerophilic conditions, chemolithoautotrophic bacteria use Fe(II) as a source of electrons and couple the reduction of nitrate (or oxygen) to the oxidation of Fe(II) to Fe(III), which then precipitates (Fe(III)[solid] is equivalent to ferrihydrite precipitates such as hematite and goethite). In the bottom half of the cycle, heterotrophic Fe(III)-reducing bacteria couple the reduction of Fe(III) to the oxidation of organic carbon (OC), whereupon Fe(II) is released to the water as Fe2+(aq). The overall reaction is the oxidation of organic carbon by nitrate or oxygen to carbon dioxide and water, nitrite or nitric oxides, and even nitrogen, which yields Gibbs energy in the form of ATP, which drives the biosynthesis and replication of the microbes. The shaded dark red areas indicate the absence of oxygen.
This explains the first of the four physiological groups of bacteria that oxidize ferrous iron, i.e., (i) acidophilic, aerobic iron oxidizers; (ii) anaerobic, photosynthetic iron oxidizers [61]; (iii) neutrophilic, microaerophilic iron oxidizers; and (iv) neutrophilic, anaerobic (nitrate-dependent) iron oxidizers.
Fully anaerobic ferrous iron oxidation is conducted by anoxygenic, phototrophic, purple, non-sulfur bacteria utilizing ferrous iron as a reductant for thermodynamically uphill carbon dioxide fixation, with light as a Gibbs energy source [62]. It is clear where the photosynthetic iron oxidizers get their Gibbs energy from, but how can we understand the energetics of the two remaining groups?
The aerobic oxidation of ferrous iron by neutrophilic microorganisms (Figure 2) may seem paradoxical as at pH = 7 the midpoint potentials of the ferrous/ferric iron couple and the oxygen–water couple are too close to allow for Gibbs energy to emerge in the process. However, the precipitation of the ferric iron as ferrihydrites such as goethite and hematite reduces the effective electron potential (−E0′) to (−E0′ = 0.2 V at pH = 7, much lower than the electron potential −E0′ = 0.8 V of the oxygen–water couple, so that the oxidation of ferrous iron by oxygen can still occur. Microaerophilic conditions are required because only then can aerobic, ferrous iron-oxidizing bacteria compete effectively with the abiotic oxidation of ferrous iron by oxygen that would dominate at atmospheric oxygen tensions [63]. Neutrophilic, microaerophilic conditions are indeed common where iron-rich waters meet an oxic-anoxic interface due to low mixing rates and the limited molecular diffusion of oxygen in water [64]. Microaerophilic, iron-oxidizing bacteria thrive in wetland soils, plant rhizospheres [65][66], places where iron seeps into groundwater supplying freshwater [67][68], and drinking water distribution systems [69]. Chemolithoautotrophic bacteria (e.g., Gallionella spp. and Sideroxydans spp.) extract their metabolic energy from iron oxidation under these conditions [70][71][72], but this is not the case for obligate heterotrophs such as the Sphaerotilus-Leptothrix groups [73]. Among the four recognized species of Leptothrix, L. ochracea is the only species for which there is circumstantial evidence of autotrophic growth using Gibbs energy derived from iron oxidation [61]. Aerobic, chemolithotrophic, magnetite-oxidizing bacteria may contribute significantly to ferrous iron oxidation at a circumneutral pH [64][67][68][71][74][75]. Currently, all known oxygen-dependent, neutrophilic, chemolithotrophic iron oxidizers belong to the Proteobacteria group, with Gallionella as the best known representative [72], belonging to the β-proteobacteria group. Gallionella sp. can also grow on organic compounds such as glucose, fructose, and sucrose [76], and sulfur (electron potential −E0′ = 0.06 V) or sulfide (−E0′ = 0.10 V) [77][78] may serve as better electron donors compared to the ferrous iron in cases where ferrihydrite precipitation is slow or problematic.
A signature of iron-oxidizing bacteria is the unique morphological structures they produce, such as sheaths (in heterotrophic species) and helical, stalk-like filaments (in autotrophic species, although autotrophic Siderooxydans spp. form neither sheaths nor stalks [61]). Excreted from the cell surface, the stalk of Gallionella acts as an organic matrix for the deposition of the ferrihydrites produced (e.g., as hematite Fe2O3) [79]. In view of the thermodynamic importance of Fe(II) oxidation to ferrihydrite (see above), these unique structures are essential for the energetics. Moreover, arsenate may be trapped by Fe(III) that binds to the stalk or other extracellular polymeric substances (EPS) on the surface of bacteria to form As(III)-Fe(III)-EPS complexes [80][81], or just by Fe(III) in magnetite.
Twisted stalks of Gallionella ferruginea may further act as a protective mechanism against precipitated ferric iron or oxygen toxicity [82]. The metals may also bind as cations to the cell surface in a passive process [58], perhaps with tighter binding of the triply charged ferric iron, thereby again increasing the iron’s electron (negative redox) potential −E0′. Many neutrophilic, iron-oxidizing bacteria can form ferric iron minerals that can co-precipitate with arsenic [83]. Also, heterotrophic Leptothrix strains are able to deposit iron oxyhydroxides onto their cell surface [84].
The biological oxidation of ferrous iron in the absence of oxygen and in dark subsurface waters is also possible by light-independent chemoautotrophic microbial activity using nitrate as the electron acceptor [85] (Figure 2). Indeed, nitrate-reducing, iron-oxidizing bacteria are the most important catalysts for the generation of ferric oxides under anaerobic conditions [63]. Nitrate-dependent, iron-oxidizing microorganisms are able to oxidize both soluble and insoluble ferrous iron minerals [86]. For the thermodynamic reasons given above, the ferric iron must occur in complexes (such as with hydroxide in ferrihydrite/magnetite): nitrate/nitrite −E0′ = −0.41 V at pH = 7 and −E0′ = −0.60 at pH = 4 are both too high in electron potential for reduction by free ferrous iron transiting to free ferric iron (−E0′ = −0.77). Even Escherichia coli is capable of nitrate-dependent iron oxidation [87].
In a further demonstration of how interactions between various inanimate and animate processes may accomplish processes that are otherwise thermodynamically impossible, there are at least three further solutions to the small Gibbs energy yield of iron respiration with nitrate as electron acceptor. One is the use of complexed ferrous iron as a substrate: Thiobacillus denitrificans oxidizes ferrous sulfide (FeS; electron potential at pH 7 of approximately −E0′ = −0.25 V, i.e., higher than the −E0′ = −0.42 V of the nitrate/nitrite couple. This nitrate-dependent iron sulfide oxidation has been demonstrated for the hyperthermophilic archaeon Ferroglobus placidus [88], the mesophilic Proteobacteria Chromobacterium violacens [89], and the Paracoccus ferrooxidans strain BDN-1 [90]. A further alternative is the nitrate reduction (−E0′ = −0.42 V) or chlorate reduction (−E0′ = −0.79 V) [91] coupled to ferrous iron oxidation in the presence of carbon and a Gibbs energy source, This has been documented for the heterotrophic Dechlorosoma suillum strain PS [92] as well as for the Acidovorax strain BoFeN1 [93].
2.4. Microbial Oxidation of Arsenite
- Oxygen as an electron acceptor
The biological oxidation of arsenite (Figure 3) is an attractive alternative to its rather slow abiotic oxidation due to its potential specificity for arsenite, efficiency, effectiveness at lower oxygen tensions (through a low KMfor oxygen), and cost effectiveness in addition to environmental friendliness [94]. In environments where significant amounts of arsenite are oxidized to arsenate within a short period of time, this oxidation can be attributed to arsenite-oxidizing bacteria [95].
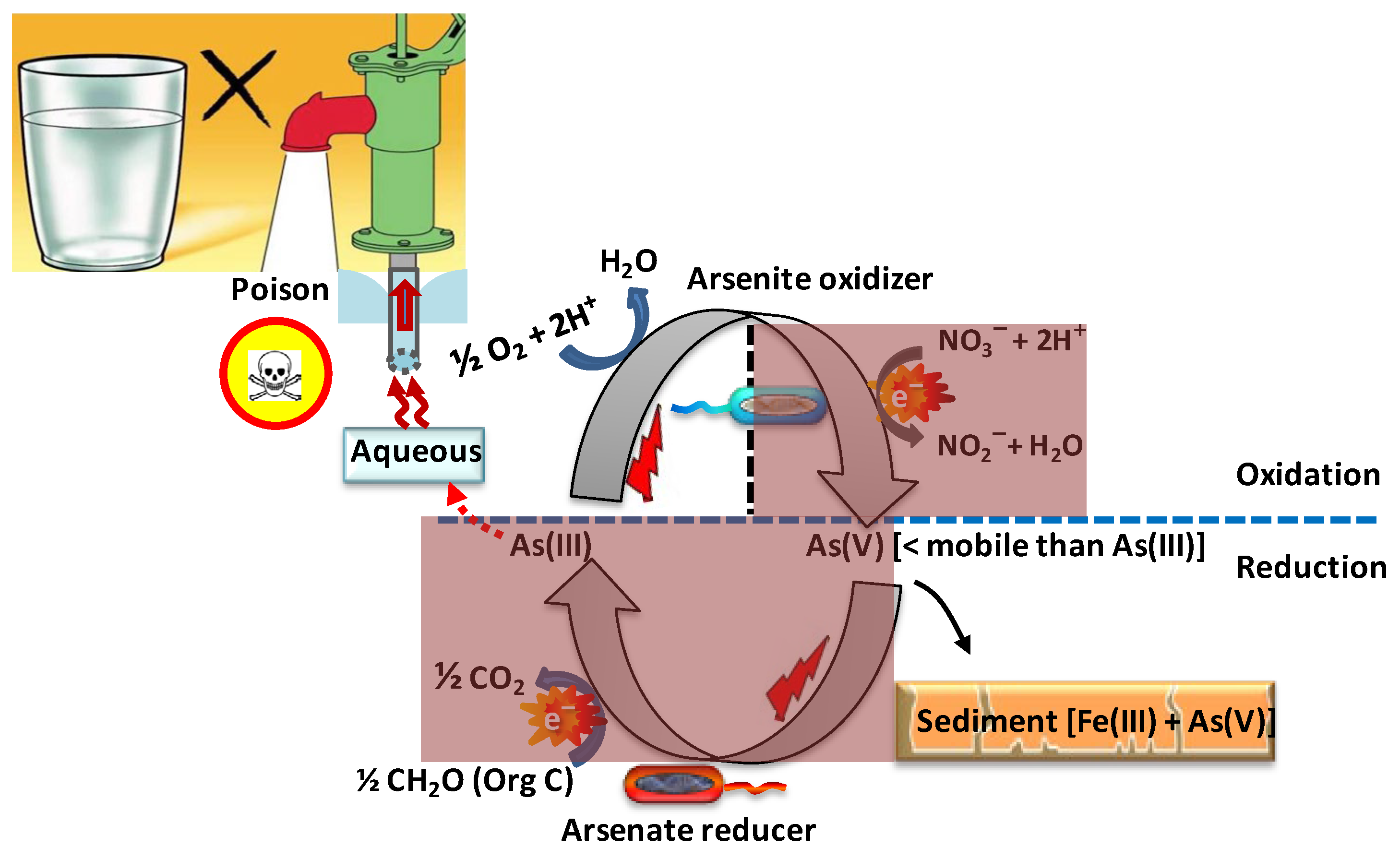
Figure 3. Scheme of putative microbial arsenic cycling in aquifers and consequences for drinking water. Microbial arsenite oxidation (above the dashed line) is mediated by a number of chemolithoautotrophs, under aerobic conditions at the surface or anaerobic conditions below that surface, using oxygen or nitrate, respectively, as the terminal electron acceptor. Microbial arsenate reduction (below the dashed line) is mediated by dissimilatory, arsenate-respiring bacteria coupling arsenate reduction under anaerobic conditions to the oxidation of organic carbon, the resulting arsenite entering into aqueous solution. The shaded dark red areas indicate the absence of oxygen.
Despite or perhaps precisely because of its biochemical toxicity (see above), arsenite is readily converted by a diversity of prokaryotes. Arsenite-oxidizing bacteria are classified into heterotrophic (HAO) and chemolithoautotrophic (CAO) arsenic oxidizers [54][96]. Heterotrophic arsenite oxidation may serve primarily as a detoxification reaction, rather than as a Gibbs energy source (in heterotrophs, other catabolic reactions readily provide the Gibbs energy required for growth): it converts arsenite encountered in the cell’s periplasmic space into the less toxic arsenate, perhaps making it less likely for the arsenic to enter the cell [35].
More than 50 phylogenetically diverse, arsenite-oxidizing (auto- and heterotrophic) species, distributed over 25 genera, have been isolated from various environments, especially mesophilic ecosystems [97]. Green, for instance, reported arsenite-oxidizing bacteria stemming from cattle dipping baths [98] and Battaglia-Brunet et al. isolated a Leptothrix sp. strain S1.1 from the settling pond sediments of mine drainage that was able to oxidize 0.1 g/L of As(III) in 1 week at 12 °C [99]. Phylogenetically, arsenite oxidizers are dispersed within the Alpha-, Beta-, and Gamma-proteobacteria; Actinobacteria; Firmicutes; and Deinococcus-Thermus. Green sulfur bacteria (e.g., Chlorobium limnicola and Chlorobium phaeobacteroides) and filamentous green non-sulfur bacteria (e.g., Chloroflexus aurantiacus) may also be capable of arsenite oxidation, as homologs of the gene-encoding arsenite oxidase (see below) have been identified in their genomes [100][101][102][103][104]. The most extensively studied heterotrophic arsenite oxidizer is Alcaligenes fecalis [54]. Little is known regarding the role of archaea in the oxidation of arsenite.
Heterotrophic Alcaligens faecalis [105] and Pseudomonas pudia [106] have not been shown to extract Gibbs energy from the oxidation of arsenite during heterotrophic growth. There is one known exception: Hydrogenophaga sp. str. NT-14, a β-proteobacterium, can oxidize arsenite whilst it grows heterotrophically, its arsenite oxidation still being coupled to the reduction of oxygen and yielding extra Gibbs energy for growth [55]. Gihring and Banfield (2001) isolated a peculiar thermophilic species of Thermus (strain HR 13) from an arsenic-rich hot spring. Under aerobic conditions, it was able to oxidize arsenite apparently for detoxification purposes, i.e., without conserving Gibbs energy. However, under anaerobic conditions, strain HR 13 can grow on lactate using arsenate as its electron acceptor, reducing it to arsenite [107].
Arsenite oxidase, located on the outer surface of the inner bacterial membrane, has been identified in both autotrophic and heterotrophic bacteria [35][54]. The enzyme is the first component of an electron transport chain that enables arsenite to reduce oxygen to water in a process coupled to proton pumping and the subsequent generation of ATP from ADP and phosphate. The genes encoding arsenite oxidase (aio genes) show considerable divergence; the aioA sequences of CAOs are phylogenetically distinct from those of HAOs [97][108]. Only two putative arsenite oxidase genes have been identified in Aeropyrum pernix and Sulfolobus tokodaii by sequence homology searches in their published genomes [109].
- Alternative electron acceptors
Molecular oxygen is poorly soluble in water (up to some 0.25 mM only, and also the rate at which it dissolves is small whenever the surface-to-volume ratio is small). Aerobic microbes in the upper oxic layers of aquifers consume dissolved oxygen, maintaining anaerobic zones below them. Anaerobic or facultative anaerobic microbes thereby become dominant in the underlying anoxic environment [110]. Alternative oxidants (e.g., nitrate; −E0′ = −0.42 V) then have the potential to support growth through the microbial oxidation of arsenite −E0′ = −0.06 V. Several studies have indeed demonstrated that anaerobic microorganisms can engage in nitrate-dependent arsenite oxidation to gain Gibbs energy [96][111]. Such arsenite-oxidizing, denitrifying bacteria have been isolated from various environments and enriched [96][112][113][114]. As their electron potential is also far below that of the arsenite/arsenate couple, besides nitrate also chlorate (ClO3−; −E0′ = −0.79 V) [115] can be an oxidant (electron acceptor) for the anaerobic microbial oxidation of arsenite. Dechloromonas sp. strain ECC1-pb1 and Azospira sp. strain ECC1-pb2 constitute examples [111].
Most arsenite-oxidizing, denitrifying organisms are Alpha, Beta, or Gamma-proteobacteria. The first identified anoxic, arsenite-oxidizing bacterium was Alkalilimnicola ehrlichii strain MLHE-1, a haloalkaliphilic facultative chemolithoautotroph: it is also able to grow heterotrophically with acetate (−E0′ = 0.29 V for CO2/acetate couple; [116]) as its electron donor, either aerobically, or anaerobically with nitrate as an electron acceptor. A novel type of arsenite oxidase gene (arxA) was identified in the genome of this extremophile, which fills a phylogenetic gap between the arsenate reductase (arrA) and arsenite oxidase (aioA) clades of arsenic-metabolizing enzymes [117]. Anoxic, chemolithoautotrophic, arsenite-oxidizing strains DAO1 and DAO10 (closely related to Sinorhizobium and Azoarcus sp., respectively) living under “normal” environmental conditions are also able to oxidize arsenite to arsenate with complete denitrification of nitrate (see above for the energetics) [112].
References
- ATSDR. Toxicological Profile for Arsenic; Report No.: 7440-38-2; Agency for Toxic Substances and Disease Registry, United States Department of Health and Human Services: Atlanta, GA, USA, 2007.
- Kumar, R.; Kumar, S.; Ghosh, A. Current scenario of groundwater arsenic contamination in West Bengal and its mitigation approach. In Arsenic Toxicity: Challenges and Solutions; Springer: Singapore, 2021; pp. 193–216.
- National Research Council. Arsenic: Medical and Biologic Effects of Environmental Pollutants; National Academies Press: Washington, DC, USA, 1977.
- Satyapal, G.K.; Kumar, N. Arsenic: Source, distribution, toxicity and bioremediation. In Arsenic Toxicity: Challenges and Solutions; Springer: Singapore, 2021; pp. 153–163.
- Herath, I.; Vithanage, M.; Bundschuh, J.; Maity, J.P.; Bhattacharya, P. Natural arsenic in global groundwaters: Distribution and geochemical triggers for mobilization. Curr. Pollut. Rep. 2016, 2, 68–89.
- Jat Baloch, M.Y.; Zhang, W.; Zhang, D.; Al Shoumik, B.A.; Iqbal, J.; Li, S.; Chai, J.; Farooq, M.A.; Parkash, A. Evolution mechanism of arsenic enrichment in groundwater and associated health risks in Southern Punjab, Pakistan. Int. J. Environ. Res. Public Health 2022, 19, 13325.
- Rehman, F.; Azeem, T.; Hashmi, R.A.; Siddique, J.; Shahab, A.; Mustafa, S. Drinking and irrigation quality of groundwater and health risk assessment due to arsenic exposure in Sheikhupura district, Punjab, Pakistan. Kuwait J. Sci. 2023, 50, 368–375.
- Kanel, S.R.; Das, T.K.; Varma, R.S.; Kurwadkar, S.; Chakraborty, S.; Joshi, T.P.; Bezbaruah, A.N.; Nadagouda, M.N. Arsenic contamination in groundwater: Geochemical basis of treatment technologies. ACS Environ. Au 2023, 3, 135–152.
- Khosravi-Darani, K.; Rehman, Y.; Katsoyiannis, I.A.; Kokkinos, E.; Zouboulis, A.I. Arsenic exposure via contaminated water and food sources. Water 2022, 14, 1884.
- Mueller, B. Ground water contamination by arsenic in Nepal: Lessons to be learned from geology. Austin Chem. Eng. 2019, 6, 1064.
- Rahman, M.S.; Reza, A.S.; Ahsan, M.A.; Siddique, M.A.B. Arsenic in groundwater from Southwest Bangladesh: Sources, water quality, and potential health concern. HydroResearch 2023, 6, 1–15.
- Chen, X.; Liu, S.; Luo, Y. Spatiotemporal distribution and probabilistic health risk assessment of arsenic in drinking water and wheat in Northwest China. Ecotoxicol. Environ. Saf. 2023, 256, 114880.
- Santha, N.; Sangkajan, S.; Saenton, S. Arsenic contamination in groundwater and potential health risk in western lampang basin, Northern Thailand. Water 2022, 14, 465.
- Zsámbok, I.; Andó, A.; Kuti, L.; Sebők, A. Toxic elements in the groundwater of Budapest, Hungary. Cent. Eur. Geol. 2014, 57, 297–305.
- Butaciu, S.; Senila, M.; Sarbu, C.; Ponta, M.; Tanaselia, C.; Cadar, O.; Roman, M.; Radu, E.; Sima, M.; Frentiu, T. Chemical modeling of groundwater in the Banat Plain, southwestern Romania, with elevated As content and co-occurring species by combining diagrams and unsupervised multivariate statistical approaches. Chemosphere 2017, 172, 127–137.
- Senila, M.; Levei, E.; Cadar, O.; Senila, L.R.; Roman, M.; Puskas, F.; Sima, M. Assessment of availability and human health risk posed by arsenic contaminated well waters from Timis-Bega area, Romania. J. Anal. Methods Chem. 2017, 2017, 3037651.
- Banning, A. Geogenic arsenic and uranium in Germany: Large-scale distribution control in sediments and groundwater. J. Hazard. Mater. 2021, 405, 124186.
- Ghezzi, L.; Arrighi, S.; Petrini, R.; Bini, M.; Vittori Antisari, L.; Franceschini, F.; Franchi, M.L.; Giannecchini, R. Arsenic contamination in groundwater, soil and the food-chain: Risk management in a densely populated area (Versilia Plain, Italy). Appl. Sci. 2023, 13, 5446.
- Shaji, E.; Santosh, M.; Sarath, K.; Prakash, P.; Deepchand, V.; Divya, B. Arsenic contamination of groundwater: A global synopsis with focus on the Indian Peninsula. Geosci. Front. 2021, 12, 101079.
- Patel, K.S.; Pandey, P.K.; Martín-Ramos, P.; Corns, W.T.; Varol, S.; Bhattacharya, P.; Zhu, Y. A review on arsenic in the environment: Contamination, mobility, sources, and exposure. RSC Adv. 2023, 13, 8803–8821.
- Podgorski, J.; Berg, M. Global threat of arsenic in groundwater. Science 2020, 368, 845–850.
- Anjum, N.A.; Gill, S.S.; Tuteja, N. Enhancing Cleanup of Environmental Pollutants; Springer: Cham, Switzerland, 2017.
- Nickson, R.; McArthur, J.; Burgess, W.; Ahmed, K.M.; Ravenscroft, P.; Rahmanñ, M. Arsenic poisoning of Bangladesh groundwater. Nature 1998, 395, 338.
- Yu, W.H.; Harvey, C.M.; Harvey, C.F. Arsenic in groundwater in Bangladesh: A geostatistical and epidemiological framework for evaluating health effects and potential remedies. Water Resour. Res. 2003, 39, 1146–1163.
- Argos, M.; Kalra, T.; Rathouz, P.J.; Chen, Y.; Pierce, B.; Parvez, F.; Islam, T.; Ahmed, A.; Rakibuz-Zaman, M.; Hasan, R.; et al. Arsenic exposure from drinking water, and all-cause and chronic-disease mortalities in Bangladesh (HEALS): A prospective cohort study. Lancet 2010, 376, 252–258.
- Nicomel, N.R.; Leus, K.; Folens, K.; Van Der Voort, P.; Du Laing, G. Technologies for arsenic removal from water: Current status and future perspectives. Int. J. Environ. Res. Public Health 2016, 13, 62.
- Chakraborti, D.; Rahman, M.M.; Das, B.; Murrill, M.; Dey, S.; Chandra Mukherjee, S.; Dhar, R.K.; Biswas, B.K.; Chowdhury, U.K.; Roy, S. Status of groundwater arsenic contamination in Bangladesh: A 14-year study report. Water Res. 2010, 44, 5789–5802.
- Smith, A.H.; Lingas, E.O.; Rahman, M. Contamination of drinking-water by arsenic in Bangladesh: A public health emergency. Bull. World Health Organ. 2000, 78, 1093–1103.
- WHO. Arsenic in Drinking-Water. Background Document for Preparation of WHO Guidelines for Drinking-Water Quality; WHO/SDE/WSH/03.04/75/Rev/1; World Health Organization: Geneva, Switzerland, 2011; Volume 1.
- Flanagan, S.V.; Johnston, R.B.; Zheng, Y. Arsenic in tube well water in Bangladesh: Health and economic impacts and implications for arsenic mitigation. Bull. World Health Organ. 2012, 90, 839–846.
- Barral-Fraga, L.; Barral, M.T.; MacNeill, K.L.; Martiñá-Prieto, D.; Morin, S.; Rodríguez-Castro, M.C.; Tuulaikhuu, B.-A.; Guasch, H. Biotic and abiotic factors influencing arsenic biogeochemistry and toxicity in fluvial ecosystems: A review. Int. J. Environ. Res. Public Health 2020, 17, 2331.
- Mladenov, N.; Zheng, Y.; Simone, B.; Bilinski, T.M.; McKnight, D.M.; Nemergut, D.; Radloff, K.A.; Rahman, M.M.; Ahmed, K.M. Dissolved organic matter quality in a shallow aquifer of Bangladesh: Implications for arsenic mobility. Environ. Sci. Technol. 2015, 49, 10815–10824.
- Cui, J.; Jing, C. A review of arsenic interfacial geochemistry in groundwater and the role of organic matter. Ecotoxicol. Environ. Saf. 2019, 183, 109550.
- Westerhoff, H.V.; Van Dam, K. Thermodynamics and Control of Biological Free-Energy Transduction; Elsevier: Amsterdam, The Netherlands; New York, NY, USA, 1987.
- Oremland, R.S.; Stolz, J.F. The ecology of arsenic. Science 2003, 300, 939–944.
- Oremland, R.S.; Stolz, J.F. Arsenic, microbes and contaminated aquifers. Trends Microbiol. 2005, 13, 45–49.
- Walter, T.; Klim, J.; Jurkowski, M.; Gawor, J.; Köhling, I.; Słodownik, M.; Zielenkiewicz, U. Plasmidome of an environmental Acinetobacter lwoffii strain originating from a former gold and arsenic mine. Plasmid 2020, 110, 102505.
- Das, S.; Liu, C.-C.; Jean, J.-S.; Lee, C.-C.; Yang, H.-J. Effects of microbially induced transformations and shift in bacterial community on arsenic mobility in arsenic-rich deep aquifer sediments. J. Hazard. Mater. 2016, 310, 11–19.
- Gadd, G.M. Metals, minerals and microbes: Geomicrobiology and bioremediation. Microbiology 2010, 156, 609–643.
- Sobecky, P.A.; Coombs, J.M. Horizontal gene transfer in metal and radionuclide contaminated soils. In Horizontal Gene Transfer; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2009; pp. 455–472.
- Hassan, Z.; Braster, M.; Röling, W.F.; Westerhoff, H.V. Microbes-assisted arsenate reduction activity in Bangladesh drinking water wells as revealed by enrichment cultivation. Geomicrobiol. J. 2023, 40, 161–171.
- Hassan, Z.; Sultana, M.; Khan, S.I.; Braster, M.; Röling, W.F.; Westerhoff, H.V. Ample arsenite bio-oxidation activity in Bangladesh drinking water wells: A bonanza for bioremediation? Microorganisms 2019, 7, 246.
- Hassan, Z.; Sultana, M.; van Breukelen, B.M.; Khan, S.I.; Röling, W.F. Diverse arsenic-and iron-cycling microbial communities in arsenic-contaminated aquifers used for drinking water in Bangladesh. FEMS Microbiol. Ecol. 2015, 91, fiv026.
- Ghosh, S.; Mohapatra, B.; Satyanarayana, T.; Sar, P. Molecular and taxonomic characterization of arsenic (As) transforming Bacillus sp. Strain IIIJ3–1 isolated from As-contaminated groundwater of Brahmaputra river basin, India. BMC Microbiol. 2020, 20, 1–20.
- Paul, D.; Kazy, S.K.; Gupta, A.K.; Pal, T.; Sar, P. Diversity, metabolic properties and arsenic mobilization potential of indigenous bacteria in arsenic contaminated groundwater of West Bengal, India. PLoS ONE 2015, 10, e0118735.
- Corsini, A.; Zaccheo, P.; Muyzer, G.; Andreoni, V.; Cavalca, L. Arsenic transforming abilities of groundwater bacteria and the combined use of Aliihoeflea sp. Strain 2WW and goethite in metalloid removal. J. Hazard. Mater. 2014, 269, 89–97.
- Fazi, S.; Amalfitano, S.; Casentini, B.; Davolos, D.; Pietrangeli, B.; Crognale, S.; Lotti, F.; Rossetti, S. Arsenic removal from naturally contaminated waters: A review of methods combining chemical and biological treatments. Rend. Lincei 2016, 27, 51–58.
- Andres, J.; Bertin, P.N. The microbial genomics of arsenic. FEMS Mcrobiol. Rev. 2016, 40, 299–322.
- Andres, J.; Arsène-Ploetze, F.; Barbe, V.; Brochier-Armanet, C.; Cleiss-Arnold, J.; Coppée, J.-Y.; Dillies, M.-A.; Geist, L.; Joublin, A.; Koechler, S. Life in an arsenic-containing gold mine: Genome and physiology of the autotrophic arsenite-oxidizing bacterium Rhizobium sp. NT-26. Genome Biol. Evol. 2013, 5, 934–953.
- Huang, J.-H. Impact of microorganisms on arsenic biogeochemistry: A review. Water Air Soil. Pollut. 2014, 225, 1–25.
- Páez-Espino, D.; Tamames, J.; de Lorenzo, V.; Cánovas, D. Microbial responses to environmental arsenic. Biometals 2009, 22, 117–130.
- Bentley, R.; Chasteen, T.G. Microbial methylation of metalloids: Arsenic, Antimony, and Bismuth. Microbiol. Mol. Biol. Rev. 2002, 66, 250–271.
- Osborne, T.H.; McArthur, J.M.; Sikdar, P.K.; Santini, J.M. Isolation of an arsenate-respiring bacterium from a redox front in an arsenic-polluted aquifer in West Bengal, Bengal Basin. Environ. Sci. Technol. 2015, 49, 4193–4199.
- Santini, J.M.; Sly, L.I.; Schnagl, R.D.; Macy, J.M. A new chemolithoautotrophic arsenite–oxidizing bacterium isolated from a gold mine: Phylogenetic, physiological, and preliminary biochemical studies. Appl. Environ. Microbiol. 2000, 66, 92–97.
- Vanden Hoven, R.N.; Santini, J.M. Arsenite oxidation by the heterotroph Hydrogenophaga sp. Str. NT-14: The arsenite oxidase and its physiological electron acceptor. Biochim. Biophys. Acta 2004, 1656, 148–155.
- Dunivin, T.K.; Yeh, S.Y.; Shade, A. A global survey of arsenic-related genes in soil microbiomes. BMC Biol. 2019, 17, 1–17.
- Mohsin, H.; Shafique, M.; Rehman, Y. Genes and biochemical pathways involved in microbial transformation of arsenic. In Arsenic Toxicity: Challenges and Solutions; Kumar, N., Ed.; Springer: Singapore, 2021; p. 391.
- Fortin, D.; Langley, S. Formation and occurrence of biogenic iron-rich minerals. Earth Sci. Rev. 2005, 72, 1–19.
- Edwards, K.J.; Bond, P.L.; Gihring, T.M.; Banfield, J.F. An archaeal iron-oxidizing extreme acidophile important in acid mine drainage. Science 2000, 287, 1796–1799.
- Baker, B.J.; Banfield, J.F. Microbial communities in acid mine drainage. FEMS Microbiol. Ecol. 2003, 44, 139–152.
- Hedrich, S.; Schlömann, M.; Johnson, D.B. The iron-oxidizing proteobacteria. Microbiology 2011, 157, 1551–1564.
- Widdel, F.; Schnell, S.; Heising, S.; Ehrenreich, A.; Assmus, B.; Schink, B. Ferrous iron oxidation by anoxygenic phototrophic bacteria. Nature 1993, 362, 834–836.
- Hohmann, C.; Winkler, E.; Morin, G.; Kappler, A. Anaerobic Fe(II)-oxidizing bacteria show as resistance and immobilize as during Fe(III) mineral precipitation. Environ. Sci. Technol. 2009, 44, 94–101.
- Emerson, D.; Weiss, J.V. Bacterial iron oxidation in circumneutral freshwater habitats: Findings from the field and the laboratory. Geomicrobiol. J. 2004, 21, 405–414.
- Emerson, D.; Weiss, J.V.; Megonigal, J.P. Iron-oxidizing bacteria are associated with ferric hydroxide precipitates (Fe-plaque) on the roots of wetland plants. Appl. Environ. Microbiol. 1999, 65, 2758–2761.
- Wang, J.; Muyzer, G.; Bodelier, P.L.; Laanbroek, H.J. Diversity of iron oxidizers in wetland soils revealed by novel 16S rRNA primers targeting Gallionella-related bacteria. ISME J. 2009, 3, 715–725.
- James, R.; Ferris, F. Evidence for microbial-mediated iron oxidation at a neutrophilic groundwater spring. Chem. Geol. 2004, 212, 301–311.
- Blöthe, M.; Roden, E.E. Microbial iron redox cycling in a circumneutral-pH groundwater seep. Appl. Environ. Microbiol. 2009, 75, 468–473.
- Li, D.; Li, Z.; Yu, J.; Cao, N.; Liu, R.; Yang, M. Characterization of bacterial community structure in a drinking water distribution system during an occurrence of red water. Appl. Environ. Microbiol. 2010, 76, 7171–7180.
- Gault, A.G.; Ibrahim, A.; Langley, S.; Renaud, R.; Takahashi, Y.; Boothman, C.; Lloyd, J.R.; Clark, I.D.; Ferris, F.G.; Fortin, D. Microbial and geochemical features suggest iron redox cycling within bacteriogenic iron oxide-rich sediments. Chem. Geol. 2011, 281, 41–51.
- Druschel, G.K.; Emerson, D.; Sutka, R.; Suchecki, P.; Luther, G.W. Low-oxygen and chemical kinetic constraints on the geochemical niche of neutrophilic iron (II) oxidizing microorganisms. Geochim. Cosmochim. Acta 2008, 72, 3358–3370.
- Emerson, D.; Fleming, E.J.; McBeth, J.M. Iron-oxidizing bacteria: An environmental and genomic perspective. Annu. Rev. Microbiol. 2010, 64, 561–583.
- Johnson, D.B.; Ghauri, M.A.; Said, M.F. Isolation and characterization of an acidophilic, heterotrophic bacterium capable of oxidizing ferrous iron. Appl. Environ. Microbiol. 1992, 58, 1423–1428.
- Neubauer, S.C.; Emerson, D.; Megonigal, J.P. Life at the energetic edge: Kinetics of circumneutral iron oxidation by lithotrophic iron-oxidizing bacteria isolated from the wetland-plant rhizosphere. Appl. Environ. Microbiol. 2002, 68, 3988–3995.
- Sobolev, D.; Roden, E.E. Evidence for rapid microscale bacterial redox cycling of iron in circumneutral environments. Antonie Van. Leeuwenhoek 2002, 81, 587–597.
- Hallbeck, L.; Pedersen, K. Autotrophic and mixotrophic growth of Gallionella ferruginea. J. Gen. Microbiol. 1991, 137, 2657–2661.
- Ghiorse, W. Biology of iron-and manganese-depositing bacteria. Annu. Rev. Microbiol. 1984, 38, 515–550.
- Lütters-Czekalla, S. Lithoautotrophic growth of the iron bacterium Gallionella ferruginea with thiosulfate or sulfide as energy source. Arch. Microbiol. 1990, 154, 417–421.
- Emerson, D.; Moyer, C.L. Neutrophilic Fe-oxidizing bacteria are abundant at the Loihi seamount hydrothermal vents and play a major role in Fe oxide deposition. Appl. Environ. Microbiol. 2002, 68, 3085–3093.
- Gehrke, T.; Hallmann, R.; Kinzler, K.; Sand, W. The EPS of Acidithiobacillus ferrooxidans—A model for structure-function relationships of attached bacteria and their physiology. Water Sci. Technol. 2001, 43, 159–167.
- Duquesne, K.; Lebrun, S.; Casiot, C.; Bruneel, O.; Personné, J.-C.; Leblanc, M.; Elbaz-Poulichet, F.; Morin, G.; Bonnefoy, V. Immobilization of arsenite and ferric iron by Acidithiobacillus ferrooxidans and its relevance to acid mine drainage. Appl. Environ. Microbiol. 2003, 69, 6165–6173.
- Hallbeck, L.; Pedersen, K. Benefits associated with the stalk of Gallionella ferruginea, evaluated by comparison of a stalk-forming and a non-stalk-forming strain and biofilm studies in situ. Microb. Ecol. 1995, 30, 257–268.
- Kappler, A.; Straub, K.L. Geomicrobiological cycling of iron. Rev. Miner. Geochem. 2005, 59, 85–108.
- Pokhrel, D.; Viraraghavan, T. Biological filtration for removal of arsenic from drinking water. J. Environ. Manag. 2009, 90, 1956–1961.
- Straub, K.L.; Benz, M.; Schink, B.; Widdel, F. Anaerobic, nitrate-dependent microbial oxidation of ferrous iron. Appl. Environ. Microbiol. 1996, 62, 1458–1460.
- Carlson, H.K.; Clark, I.C.; Blazewicz, S.J.; Iavarone, A.T.; Coates, J.D. Fe(II) oxidation is an innate capability of nitrate-reducing bacteria that involves abiotic and biotic reactions. J. Bacteriol. 2013, 195, 3260–3268.
- Brons, H.J.; Hagen, W.R.; Zehnder, A.J. Ferrous iron dependent nitric oxide production in nitrate reducing cultures of Escherichia coli. Arch. Microbiol. 1991, 155, 341–347.
- Hafenbradl, D.; Keller, M.; Dirmeier, R.; Rachel, R.; Roßnagel, P.; Burggraf, S.; Huber, H.; Stetter, K.O. Ferroglobus placidus gen. nov., sp. nov., a novel hyperthermophilic archaeum that oxidizes Fe2+ at neutral pH under anoxic conditions. Arch. Microbiol. 1996, 166, 308–314.
- Weber, K.A.; Pollock, J.; Cole, K.A.; O’Connor, S.M.; Achenbach, L.A.; Coates, J.D. Anaerobic nitrate-dependent iron(II) bio-oxidation by a novel lithoautotrophic betaproteobacterium, strain 2002. Appl. Environ. Microbiol. 2006, 72, 686–694.
- Kumaraswamy, R.; Sjollema, K.; Kuenen, G.; Van Loosdrecht, M.; Muyzer, G. Nitrate-dependent 2− oxidation by Paracoccus ferrooxidans sp. nov., isolated from a denitrifying bioreactor. Syst. Appl. Microbiol. 2006, 29, 276–286.
- Liebensteiner, M.; Stams, A.; Lomans, B. (Per)chlorate reduction at high temperature: Physiological study of Archaeoglobus fulgidus and potential implications for novel souring mitigation strategies. Int. Biodeterior. Biodegrad. 2014, 96, 216–222.
- Chaudhuri, S.K.; Lack, J.G.; Coates, J.D. Biogenic magnetite formation through anaerobic biooxidation of Fe(II). Appl. Environ. Microbiol. 2001, 67, 2844–2848.
- Kappler, A.; Schink, B.; Newman, D.K. Fe(III) mineral formation and cell encrustation by the nitrate-dependent Fe (II)-oxidizer strain BoFeN1. Geobiology 2005, 3, 235–245.
- Le Nguyen, A.; Sato, A.; Inoue, D.; Sei, K.; Soda, S.; Ike, M. Bacterial community succession during the enrichment of chemolithoautotrophic arsenite oxidizing bacteria at high arsenic concentrations. J. Environ. Sci. 2012, 24, 2133–2140.
- Tamaki, S.; Frankenberger, W.T. Environmental biochemistry of arsenic. Rev. Environ. Contam. Toxicol. 1992, 124, 79–110.
- Oremland, R.S.; Hoeft, S.E.; Santini, J.M.; Bano, N.; Hollibaugh, R.A.; Hollibaugh, J.T. Anaerobic oxidation of arsenite in Mono Lake water and by a facultative, arsenite-oxidizing chemoautotroph, strain MLHE-1. Appl. Environ. Microbiol. 2002, 68, 4795–4802.
- Quéméneur, M.; Heinrich-Salmeron, A.; Muller, D.; Lièvremont, D.; Jauzein, M.; Bertin, P.N.; Garrido, F.; Joulian, C. Diversity surveys and evolutionary relationships of aoxB genes in aerobic arsenite-oxidizing bacteria. Appl. Environ. Microbiol. 2008, 74, 4567–4573.
- Green, H.H. Isolation and description of a bacterium causing oxidation of arsenite to arsenate in cattle dipping baths. Rep. Dir. Vet. S. Afr. 1919, 5, 593–599.
- Battaglia-Brunet, F.; Itard, Y.; Garrido, F.; Delorme, F.; Crouzet, C.; Greffié, C.; Joulian, C. A simple biogeochemical process removing arsenic from a mine drainage water. Geomicrobiol. J. 2006, 23, 201–211.
- Herrera, C.; Moraga, R.; Bustamante, B.; Vilo, C.; Aguayo, P.; Valenzuela, C.; Smith, C.T.; Yáñez, J.; Guzmán-Fierro, V.; Roeckel, M. Characterization of arsenite-oxidizing bacteria isolated from arsenic-rich sediments, atacama desert, Chile. Microorganisms 2021, 9, 483.
- Campos, V.L.; Valenzuela, C.; Yarza, P.; Kämpfer, P.; Vidal, R.; Zaror, C.; Mondaca, M.-A.; Lopez-Lopez, A.; Rosselló-Móra, R. Pseudomonas arsenicoxydans sp nov., an arsenite-oxidizing strain isolated from the Atacama desert. Syst. Appl. Microbiol. 2010, 33, 193–197.
- Stolz, J.F.; Basu, P.; Santini, J.M.; Oremland, R.S. Arsenic and selenium in microbial metabolism. Annu. Rev. Microbiol. 2006, 60, 107–130.
- Guo, H.; Liu, Z.; Ding, S.; Hao, C.; Xiu, W.; Hou, W. Arsenate reduction and mobilization in the presence of indigenous aerobic bacteria obtained from high arsenic aquifers of the Hetao basin, Inner Mongolia. Environ. Pollut. 2015, 203, 50–59.
- Koechler, S.; Cleiss-Arnold, J.; Proux, C.; Sismeiro, O.; Dillies, M.-A.; Goulhen-Chollet, F.; Hommais, F.; Lièvremont, D.; Arsène-Ploetze, F.; Coppée, J.-Y. Multiple controls affect arsenite oxidase gene expression in Herminiimonas arsenicoxydans. BMC Microbiol. 2010, 10, 1–13.
- Osborne, F.H.; Ehrlich, H.L. Oxidation of arsenite by a soil isolate of Alcaligenes. J. Appl. Microbiol. 1976, 41, 295–305.
- Abdrashitova, S.A.; Mynbaeva, B.N.; Ilialetdinov, A.N. Arsenic oxidation by the heterotrophic bacteria Pseudomonas putida and Alcaligenes eutrophus. Mikrobiologiia 1981, 50, 41–45.
- Gihring, T.M.; Banfield, J.F. Arsenite oxidation and arsenate respiration by a new Thermus isolate. FEMS Microbiol. Lett. 2001, 204, 335–340.
- Rhine, E.D.; Ni Chadhain, S.M.; Zylstra, G.J.; Young, L.Y. The arsenite oxidase genes (aroAB) in novel chemoautotrophic arsenite oxidizers. Biochem. Biophys. Res. Commun. 2007, 354, 662–667.
- Lebrun, E.; Brugna, M.; Baymann, F.; Muller, D.; Lièvremont, D.; Lett, M.-C.; Nitschke, W. Arsenite oxidase, an ancient bioenergetic enzyme. Mol. Biol. Evol. 2003, 20, 686–693.
- Dowling, C.B.; Poreda, R.J.; Basu, A.R.; Peters, S.L.; Aggarwal, P.K. Geochemical study of arsenic release mechanisms in the Bengal Basin groundwater. Water Resour. Res. 2002, 38, 1173.
- Sun, W.; Sierra-Alvarez, R.; Milner, L.; Field, J.A. Anaerobic oxidation of arsenite linked to chlorate reduction. Appl. Environ. Microbiol. 2010, 76, 6804–6811.
- Rhine, E.D.; Phelps, C.D.; Young, L.Y. Anaerobic arsenite oxidation by novel denitrifying isolates. Environ. Microbiol. 2006, 8, 899–908.
- Sun, W.; Sierra-Alvarez, R.; Fernandez, N.; Sanz, J.L.; Amils, R.; Legatzki, A.; Maier, R.M.; Field, J.A. Molecular characterization and in situ quantification of anoxic arsenite-oxidizing denitrifying enrichment cultures. FEMS Microbiol. Ecol. 2009, 68, 72–85.
- Sun, W.; Sierra, R.; Field, J.A. Anoxic oxidation of arsenite linked to denitrification in sludges and sediments. Water Res. 2008, 42, 4569–4577.
- Liebensteiner, M.G.; Tsesmetzis, N.; Stams, A.J.; Lomans, B.P. Microbial redox processes in deep subsurface environments and the potential application of (per)chlorate in oil reservoirs. Front. Microbiol. 2014, 5, 428.
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 1977, 41, 100–180.
- Zargar, K.; Hoeft, S.; Oremland, R.; Saltikov, C.W. Identification of a novel arsenite oxidase gene, arxA, in the haloalkaliphilic, arsenite-oxidizing bacterium Alkalilimnicola ehrlichii strain MLHE-1. J. Bacteriol. 2010, 192, 3755–3762.
More
Information
Subjects:
Ecology; Water Resources; Thermodynamics
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.2K
Revisions:
2 times
(View History)
Update Date:
31 Jan 2024
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No