 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Tiziana Persico | -- | 2180 | 2024-01-17 08:26:54 | | | |
| 2 | Lindsay Dong | Meta information modification | 2180 | 2024-01-19 02:16:08 | | |
Video Upload Options
Primary ovarian failure (POF) is caused by follicle exhaustion and is associated with menstrual irregularities and elevated gonadotropin levels, which lead to infertility before the age of 40 years. The etiology of POI is mostly unknown, but a heterogeneous genetic and familial background can be identified in a subset of cases. Abnormalities in the fragile X mental retardation 1 gene (FMR1) are among the most prevalent monogenic causes of POI. These abnormalities are caused by the expansion of an unstable CGG repeat in the 5′ untranslated region of FMR1. Expansions over 200 repeats cause fragile X syndrome (FXS), whereas expansions between 55 and 200 CGG repeats, which are defined as a fragile X premutation, have been associated with premature ovarian failure type 1 (POF1) in heterozygous females. Preimplantation genetic testing for monogenic diseases (PGT-M) can be proposed when the female carries a premutation or a full mutation.
1. Introduction
2. Premature Ovarian Failure
3. FMR1
4. POF1
5. FMR1 Premutation and Ovarian Damage
6. Reproductive Counseling and Fertility Preservation
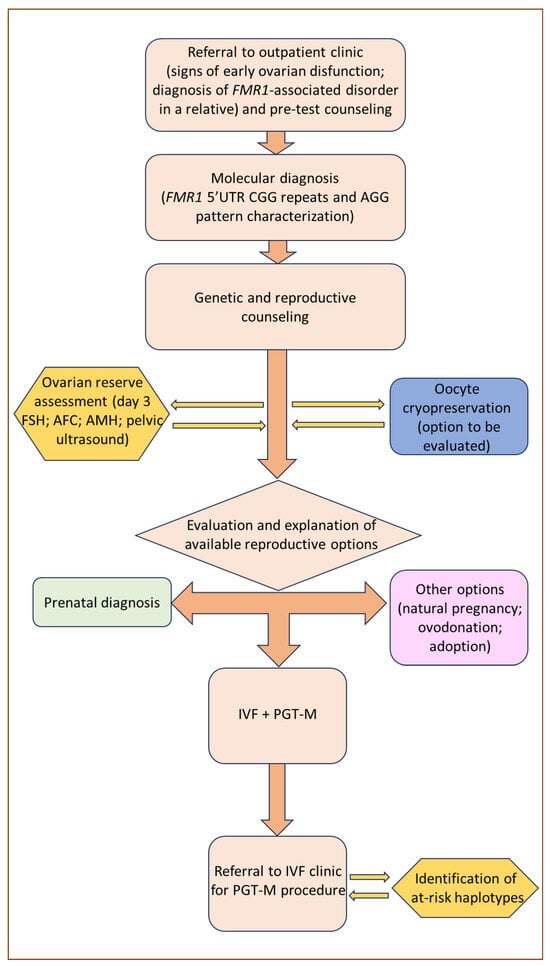
7. PGT-M Strategies and Limitations in FMR1-Related Disorders
8. POF1 in the PGT-M Context
9. Conclusions
References
- American College of Obstetricians and Gynecologists. Primary ovarian insufficiency in adolescents and young women. Obs. Gynecol. 2014, 124, 193–197.
- European Society for Human Reproduction and Embryology (ESHRE) Guideline Group on POI; Webber, L.; Davies, M.; Anderson, R.; Bartlett, J.; Braat, D.; Cartwright, B.; Cifkova, R.; de Muinck Keizer-Schrama, S.; Hogervorst, E.; et al. Management of women with premature ovarian insufficiency. Hum. Reprod. 2016, 31, 926–937.
- Ishizuka, B. Current Understanding of the Etiology, Symptomatology, and Treatment Options in Premature Ovarian Insufficiency (POI). Front. Endocrinol. 2021, 12, 626924.
- Cheng, M.H.; Nelson, L.M. Mechanisms and models of immune tolerance breakdown in the ovary. Semin. Reprod. Med. 2011, 29, 308–316.
- Qin, Y.; Jiao, X.; Simpson, J.L.; Chen, Z.-J. Genetics of primary ovarian insufficiency: New developments and opportunities. Hum. Reprod. Update 2015, 21, 787–808.
- Aittomäki, K.; Lucena, J.L.; Pakarinen, P.; Sistonen, P.; Tapanainen, J.; Gromoll, J.; Kaskikari, R.; Sankila, E.M.; Lehväslaiho, H.; Engel, A.R.; et al. Mutation in the follicle-stimulating hormone receptor gene causes hereditary hypergonadotropic ovarian failure. Cell 1995, 82, 959–968.
- Verkerk, A.J.M.H.; Pieretti, M.; Sutcliffe, J.S.; Fu, Y.-H.; Kuhl, D.P.P.A.; Pizzuti, A.; Reiner, O.; Richards, S.; Victoria, M.F.; Zhang, F.; et al. Identification of a gene (FMR-1) containing a CGG repeat coincident with a breakpoint cluster region exhibiting length variation in fragile X syndrome. Cell 1991, 65, 905–914.
- Lubs, H.A.; Stevenson, R.E.; Schwartz, C.E. Fragile X and X-linked intellectual disability: Four decades of discovery. Am. J. Hum. Genet. 2012, 90, 579–590.
- Vincent, A.; Heitz, D.; Petit, C.; Kretz, C.; Oberlé, I.; Mandel, J.L. Abnormal pattern detected in fragile-X patients by pulsed-field gel electrophoresis. Nature 1991, 349, 624–626.
- Bell, M.V.; Hirst, M.C.; Nakahori, Y.; MacKinnon, R.N.; Roche, A.; Flint, T.J.; Jacobs, P.A.; Tommerup, N.; Tranebjaerg, L.; Froster-Iskenius, U.; et al. Physical mapping across the fragile X: Hypermethylation and clinical expression of the fragile X syndrome. Cell 1991, 64, 861–866.
- Kooy, R.F.; Willemsen, R.; Oosatra, B.A. Fragile X syndrome at turn of the century. Mol. Med. Today 2000, 6, 193–198.
- Nolin, S.L.; Glicksman, A.; Ersalesi, N.; Dobkin, C.; Brown, W.T.; Cao, R.; Blatt, E.; Sah, S.; Latham, G.J.; Hadd, A.G. Fragile X full mutation expansions are inhibited by one or more AGG interruptions in premutation carriers. Genet. Med. 2014, 17, 358–364.
- Eichler, E.E.; Holden, J.J.; Popovich, B.W.; Reiss, A.L.; Snow, K.; Thibodeau, S.N.; Richards, C.S.; Ward, P.A.; Nelson, D.L. Length of uninterrupted CGG repeats determines instability in the FMR1 gene. Nat. Genet. 1994, 8, 88–94.
- Yrigollen, C.M.; Durbin-Johnson, B.; Gane, L.; Nelson, D.L.; Hagerman, R.; Hagerman, P.J.; Tassone, F. AGG interruptions within the maternal FMR1 gene reduce the risk of offspring with fragile X syndrome. Genet. Med. 2012, 14, 729–736.
- Nelson, L.M. Clinical practice. Primary ovarian insufficiency. N. Engl. J. Med. 2009, 360, 606–614.
- Sullivan, A.; Marcus, M.; Epstein, M.; Allen, E.; Anido, A.; Paquin, J.; Yadav-Shah, M.; Sherman, S. Association of FMR1 repeat size with ovarian dysfunction. Hum. Reprod. 2005, 20, 402–412.
- Allingham-Hawkins, D.J.; Babul-Hirji, R.; Chitayat, D.; Holden, J.J.A.; Yang, K.T.; Lee, C.; Hudson, R.; Gorwill, H.; Nolin, S.R.; Glicksman, A.; et al. Fragile X Premutation Is a Significant Risk Factor for Premature Ovarian Failure. Am. J. Med. Genet. 1999, 83, 322–325.
- Cronister, A.; Schreiner, R.; Wittenberger, M.; Amiri, K.; Harris, K.; Hagerman, R.J. Heterozygous fragile X female: Historical, physical, cognitive, and cytogenetic features. Am. J. Med. Genet. 1991, 38, 269–274.
- Welt, C.K. Primary ovarian insufficiency: A more accurate term for premature ovarian failure. Clin. Endocrinol. 2008, 68, 499–509.
- Bodega, B.; Bione, S.; Dalprà, L.; Toniolo, D.; Ornaghi, F.; Vegetti, W.; Ginelli, E.; Marozzi, A. Influence of intermediate and uninterrupted FMR1 CGG expansions in premature ovarian failure manifestation. Hum. Reprod. 2006, 21, 952–957.
- Ennis, S.; Ward, D.; Murray, A. Nonlinear association between CGG repeat number and age of menopause in FMR1 premutation carriers. Eur. J. Hum. Genet. 2006, 14, 253–255.
- Allen, E.S.; Charen, K.; Hipp, H.S.; Shubeck, L.; Amin, A.; He, W.; Nolin, S.L.; Glicksman, A.; Tortora, N.; McKinnon, B.; et al. Refining the risk for fragile X-associated primary ovarian insufficiency (POF1) by FMR1 CGG repeat size. Genet. Med. 2021, 23, 1648–1655.
- Tassone, F.; Iong, K.P.; Tong, T.H.; Lo, J.; Gane, L.W.; Berry-Kravis, E.; Nguyen, D.; Mu, L.Y.; Laffin, J.; Bailey, D.B.; et al. FMR1 CGG allele size and prevalence ascertained through newborn screening in the United States. Genome Med. 2012, 4, 100.
- Schwartz, C.E.; Dean, J.; Howard-Peebles, P.N.; Bugge, M.; Mikkelsen, M.; Tommerup, N.; Hull, C.; Hagerman, R.; Holden, J.J.; Stevenson, R.E. Obstetrical and gynecological complication in fragile X carriers: A multicenter study. Am. J. Med. Genet. 1994, 51, 400–402.
- Streuli, I.; Fraisse, T.; Ibecheole, V.; Moix, I.; Morris, M.A.; de Ziegler, D. Intermediate and premutation FMR1 alleles in women with occult primary ovarian insufficiency. Fertil. Steril. 2009, 92, 464–470.
- Ishizuka, B.; Okamoto, N.; Hamada, N.; Sugishita, Y.; Saito, J.; Takahashi, N.; Ogata, T.; Itho, M.T. Number of CGG repeats in the FMR1 gene of Japanese patients with primary ovarian insufficiency. Fertil. Steril. 2011, 96, 1170–1174.
- Gleicher, N.; Weghofer, A.; Barad, D.H. Apilot study of premature ovarian senescence: Correlation of triple CGG repeats on the FMR1 gene to ovarian reserve parameters FSH and anti-Mullerian hormone. Fertil. Steril. 2009, 91, 1700–1706.
- Barasoain, M.; Barrenetxea, G.; Huerta, I.; Télez, M.; Criado, B.; Arrieta, I. Study of the Genetic Etiology of Primary Ovarian Insufficiency: FMR1 Gene. Genes 2016, 7, 123.
- Wittenberger, M.D.; Hagerman, R.J.; Sherman, S.L.; McConkie-Rosell, A.; Welt, C.K.; Rebar, R.W.; Corrigan, E.C.; Simpson, J.L.; Nelson, L.M. The FMR1 premutation and reproduction. Fertil. Steril. 2007, 87, 456–465.
- Sherman, S.L. Premature ovarian failure among fragile X premutation carriers: Parent of origin effect? Am. J. Hum. Genet. 2000, 67, 11–13.
- Vianna-Morgante, A.M.; Costa, S.S.; Pavanello, R.C.; Otto, P.A.M.; Mingroni-Netto, R.C. Premature ovarian failure (POF) in Brazilian fragile X carriers. Genet. Mol. Biol. 1999, 22, 471–474.
- Murray, A.; Ennis, S.; MacSwiney, F.; Webb, J.; Morton, N.E. Reproductive and menstrual history of females with fragile X expansions. Eur. J. Hum. Genet. 2000, 8, 247–252.
- Mallolas, J.; Duran, M.; Sánchez, A.; Jimenez, D.; Castellvi-Bel, S.; Rife, M.; Milá, M. Implications of the FMR1 gene in menopause: Study of 147 Spanish women. Menopause 2001, 8, 106–110.
- Hundscheid, R.D.; Smits, A.O.; Thomas, C.M.; Kiemeney, L.A.; Braat, D.D. Female carriers of fragile X premutations have no increased risk for additional diseases other than premature ovarian failure. Am. J. Med. Genet. 2003, 117A, 6–9.
- Allen, E.G.; He, W.; Yadav-Shah, M.; Sherman, S.L. A Study of the distributional characteristics of FMR1 transcript levels in 238 individuals. Hum. Genet. 2004, 114, 439–447.
- Tassone, F.; Bellina, A.; Carosi, C.; Albertosi, S.; Bagni, C.; Li, L.; Glover, K.; Bentley, D.; Hanerman, P.J. Elevated FMR1 mRNA in premutation carriers is due to increased transcription. RNA 2007, 13, 555–562.
- Kenneson, A.; Zhang, F.; Hagedorn, C.H.; Warren, S.T. Reduced FMRP and increased FMR1 transcription is proportionally associated with CGG repeat number in intermediate-length and premutation carriers. Hum. Mol. Genet. 2001, 10, 1449–1454.
- Peprah, E.; He, W.; Allen, E.; Oliver, T.; Boyne, A.; Sherman, S.L. Examination of FMR1 transcript and protein levels among 74 premutation carriers. J. Hum. Genet. 2010, 55, 66–68.
- Garcia-Alegria, E.; Ibanez, B.; Minguez, M.; Poch, M.; Valiente, A.; Sanz-Parra, A.; Martinez-Bouzas, C.; Beristain, E.; Tejada, M.I. Analysis of FMR1 gene expression in female premutation carriers using robust segmented linear regression models. RNA 2007, 13, 756–762.
- Hagerman, R.J.; Leehey, M.; Heinrichs, W.; Tassone, F.; Wilson, R.; Hills, J.; Grigsby, J.; Gage, B.; Hagerman, P.J. Intention tremor, parkinsonism, and generalized brain atrophy in male carriers of fragile X. Neurology 2001, 57, 127–130.
- Elizur, S.E.; Lebovitz, O.; Derech-Haim, S.; Dratviman-Storobinsky, O.; Feldman, B.; Dor, J.; Orvieto, T.; Cohen, Y. Elevated levels of FMR1 mRNA in granulosa cells are associated with low ovarian reserve in FMR1 premutation carriers. PLoS ONE 2014, 9, e105121.
- Allen, E.G.; Sullivan, A.K.; Marcus, M.; Small, C.; Dominguez, C.; Epstein, M.P.; Charen, K.; He, W.; Taylor, K.C.; Sherman, S.L. Examination of reproductive aging milestones among women who carry the FMR1 premutation. Hum. Reprod. 2007, 22, 2142–2152.
- Napierala, M.; Michalowski, D.; de Mezer, M.; Krzyzosiak, W.J. Facile FMR1 mRNA structure regulation by interruptions in CGG repeats. Nucleic Acids Res. 2005, 33, 451–463.
- Sonigo, C.; Mayeur, A.; Sadoun, M.; Pinto, M.; Benguigui, J.; Frydman, N.; Monnot, S.; Benachi, A.; Steffann, J.; Grynberg, M. What is the threshold of mature oocytes to obtain at least one healthy transferable cleavage-stage embryo after preimplantation genetic testing for fragile X syndrome? Hum. Reprod. 2021, 36, 3003–3013.
- Murray, A. Premature ovarian failure and the FMR1 gene. Semin. Reprod. Med. 2000, 18, 59–66.
- La Marca, A.; Mastellari, E. Fertility preservation for genetic diseases leading to premature ovarian insufficiency (POI). J. Assist. Reprod. Genet. 2021, 38, 759–777.
- Vandervorst, M.; Liebaers, I.; Sermon, K.; Staessen, C.; De Vos, A.; Van de Velde, H.; Van Assche, E.; Joris, H.; Van Steirteghem, A.; Devroey, P. Successful preimplantation genetic diagnosis is related to the number of available cumulus-oocyte complexes. Hum. Reprod. 1998, 13, 3169–3176.
- Wheeler, A.C.; Raspa, M.; Green, A.; Bishop, E.; Bann, C.; Edwards, A.; Bailey, D.B., Jr. Health and reproductive experiences of women with an FMR1 premutation with and without fragile X premature ovarian insufficiency. Front. Genet. 2014, 5, 300.
- Sullivan, N.; de Faoite, E. Psychological Impact of Abortion due to Fetal Anomaly: A Review of Published Research. Issues Law. Med. 2017, 32, 19–30.
- Ranisavljevic, N.; Hess, M.; Castelli, C.; Willems, M.; Ferrieres-Hoa, A.; Girardet, A.; Anahory, T. Are ovarian response and pregnancy rates similar in selected FMR1 premutated and mutated patients undergoing preimplantation genetic testing? J. Assist. Reprod. Genet. 2020, 37, 1675–1683.
- Platteau, P.; Sermon, K.; Seneca, S.; Van Steirteghem, A.; Devroey, P.; Liebaers, I. Preimplantation genetic diagnosis for fragile X syndrome: Difficult but not impossible. Hum. Reprod. 2002, 17, 2807–2812.
- Chen, M.; Zhao, M.; Lee, C.G.; Chong, S.S. Identification of microsatellite markers <1 Mb from the FMR1 CGG repeat and development of a single-tube tetradecaplex PCR panel of highly polymorphic markers for preimplantation genetic diagnosis of fragile X syndrome. Genet. Med. 2016, 18, 869–875.
- Dreesen, J.C.; Jacobs, L.J.; Bras, M.; Herbergs, J.; Dumoulin, J.C.; Geraedts, J.P.; Evers, J.L.; Smeets, H.J. Multiplex PCR of polymorphic markers flanking the CFTR gene; a general approach for preimplantation genetic diagnosis of cystic fibrosis. Mol. Hum. Reprod. 2000, 6, 391–396.
- Apessos, A.; Abou-Sleiman, P.M.; Harper, J.C.; Delhanty, J.D.A. Preimplantation genetic diagnosis of the fragile X syndrome by use of linked polymorphic markers. Prenat. Diagn. 2001, 21, 504–511.
- Malcov, M.; Naiman, T.; Ben Yosef, D.; Carmon, A.; Mey-Raz, N.; Amit, A.; Vagman, I.; Yaron, Y. Preimplantation genetic diagnosis for fragile X syndrome using multiplex nested PCR. Reprod. Biomed. Online 2007, 14, 515–521.
- Kieffer, E.; Nicod, J.C.; Gardes, N.; Kastner, C.; Becker, N.; Celebi, C.; Pirrello, O.; Rongières, C.; Koscinski, I.; Gosset, P.; et al. Improving preimplantation genetic diagnosis for Fragile X syndrome: Two new powerful single-round multiplex indirect and direct tests. Eur. J. Hum. Genet. 2016, 24, 221–227.
- Hutchinson, A.P.; Pereira, N.; Lilienthal, D.P.; Coveney, S.; Lekovich, J.P.; Elias, R.T.; Rosenwaks, Z. Impact of FMR1 Pre-Mutation Status on Blastocyst Development in Patients Undergoing Pre-Implantation Genetic Diagnosis. Gynecol. Obs. Investig. 2018, 83, 23–28.
- Avraham, S.; Almog, B.; Reches, A.; Zakar, L.; Malcov, M.; Sokolov, A.; Alpern, S.; Azem, F. The ovarian response in fragile X patients and premutation carriers undergoing IVF-PGD: Reappraisal. Hum. Reprod. 2017, 32, 1508–1511.
- Pastore, L.M.; Young, S.L.; Baker, V.L.; Karns, L.B.; Williams, C.D.; Silverman, L.M. Elevated prevalence of 35-44 FMR1 trinucleotide repeats in women with diminished ovarian reserve. Reprod. Sci. 2012, 19, 1226–1231.
- Nayot, D.; Chung, J.T.; Son, W.Y.; Ao, A.; Hughes, M.; Dahan, M.H. Live birth following serial vitrification of embryos and PGD for fragile X syndrome in a patient with the premutation and decreased ovarian reserve. J. Assist. Reprod. Genet. 2013, 30, 1439–1444.


