 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Karen Bohmwald | -- | 6896 | 2024-01-11 13:38:19 | | | |
| 2 | Catherine Yang | Meta information modification | 6896 | 2024-01-12 01:37:46 | | |
Video Upload Options
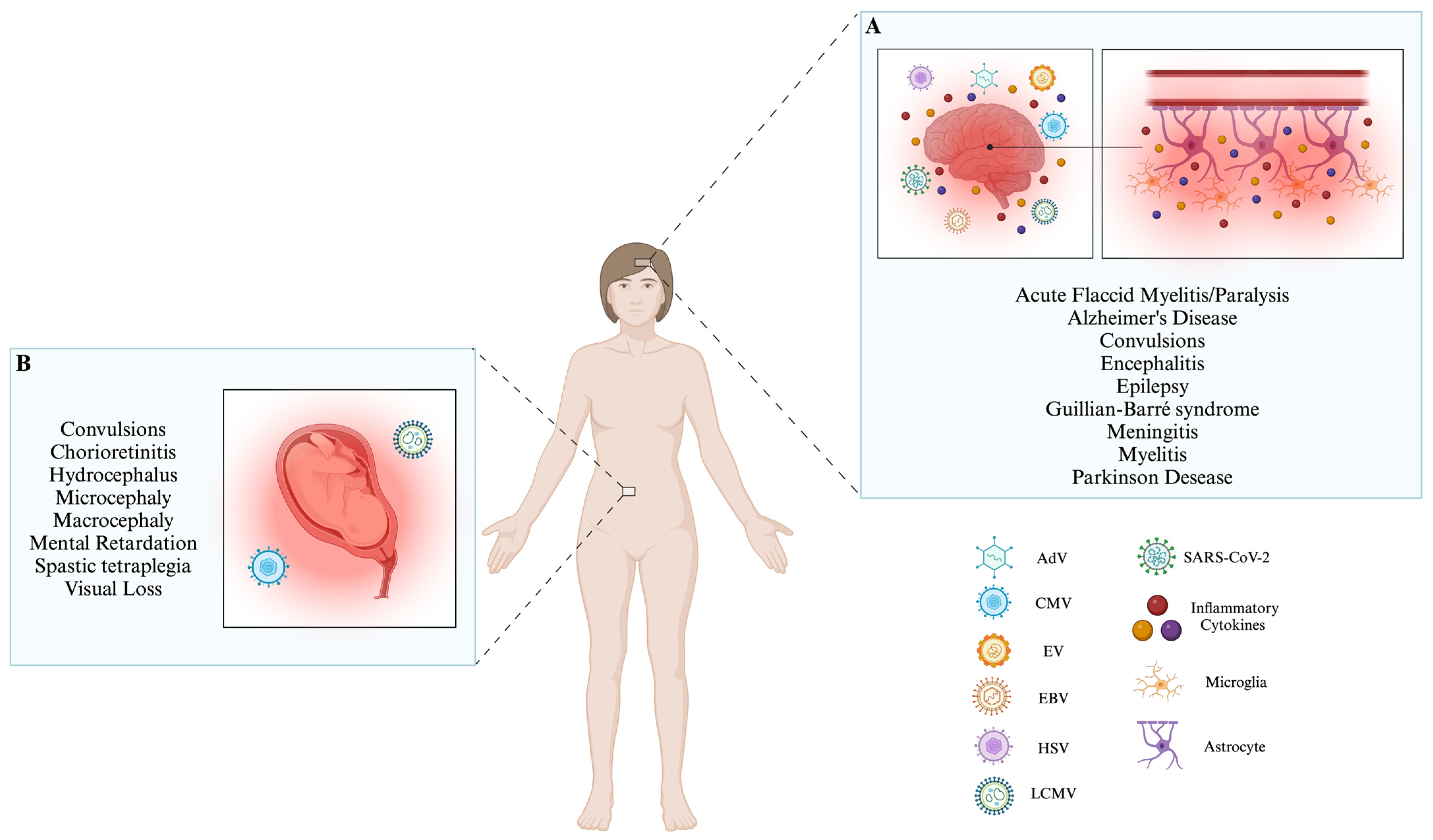
Neurotrophic viruses are pathogens that can cause alterations in the function of the central nervous system (CNS). These viruses can enter the CNS through the previously mentioned pathways. Additionally, it has been observed that viruses whose genome corresponds to RNA can be introduced into the CNS through nerves, such as the sciatic nerve. Some can even transport immune cells, allowing them to arrive at the CNS. On the other hand, double-stranded DNA (dsDNA) viruses can enter the CNS by penetrating the blood–brain barrier, utilizing brain microvascular endothelial cells, and even using sensory nerve endings and olfactory receptor neurons to enter the CNS. When the infection occurs, the main mechanism that the CNS possesses to defend itself is the activation of microglia, which induce the activation of the innate immune response and a subsequent adaptive immune response. Even the microglia can act with astrocytes to induce the chemotaxis of immune cells, like neutrophils, to combat infection generated in the CNS.
1. Adenovirus
| Virus | Genome | Invasion to CNS | Immune Response Evasion | Clinical Signs | References |
|---|---|---|---|---|---|
| AdV | dsDNA | BBB, and Olfactory nerves | IFN-I pathway modulates the innate and adaptive immune response, and affects antigen presentation processes |
Aseptic meningitis convulsions, encephalitis, and acute disseminated encephalomyelitis. | [1][8][9][10][11][12][13][14][15][16] |
| CMV | dsDNA | BBB | Affects antigen presentation processes module phenotypic and functional changes in macrophages. | Aseptic meningitis, polyradiculopathy, myelitis, ventriculitis, dementia, cranial nerve involvement, microcephaly, periventricular calcifications, convulsions, spastic tetraplegia, and hydrocephalus suppress the development of neural stem cells. | [8][9][17][18][19][20][21][22][23][24][25] |
| EV | ssRNA− | BBB, Myeloid cells infected, motoneurons. | IFN-I pathway regulates the cytosolic RIG-I pathway and NLRP3 inflammasome. | Aseptic meningitis, encephalitis, AFM, cognitive impairments in motor skills, problem-solving abilities, socialization, communication, neuronal development delay, ADHD, and epilepsy. | [8][9][26][27][28][29][30][31][32][33][34][35][36][37][38][39] |
| EBV | dsDNA | BBB | IFN-I pathway, MAVS, and NLRP3 inflammasome induce apoptosis and inhibit DC development, reprogramming B cells. | Aseptic meningitis, encephalitis, transverse myelitis, cerebellitis, ataxia, Guillian–Barré syndrome, Alzheimer’s Disease, and Parkinson’s disease. |
[8][9][40][41][42][43][44][45][46][47][48][49][50][51][52][53] |
| HSV | dsDNA | BBB, trigeminal or olfactory nerves, latent HSV-1 in situ in the brain, and reactivation of initial infection in the periphery. | IFN-I pathway hinders the complement activation, hinders the ability of IgG antibodies, induces DCs apoptosis, and affects antigen presentation processes. | Aseptic meningitis, acute encephalitis, and cognitive impairment. | [8][9][54][55][56][57][58][59][60][61][62][63][64][65][66][67][68][69] |
| LCMV | ssRNA− | BBB | IFN-I production. | Aseptic meningitis, hydrocephalus, chorioretinitis, macrocephaly, microcephaly, chorioretinopathy, spastic quadriparesis, seizures, visual loss, mental retardation, and neurodegenerative manifestations. | [8][9][70][71][72][73][74][75][76][77][78][79] |
| SARS-CoV-2 | ssRNA+ | BBB, and cranial nerves. | It affects dsRNA recognition via the RIG-I IFN pathway, induces MAVS autophagy, modules the immune chemotaxis, and affects antigen presentation processes. | Encephalitis, encephalopathy, seizures, epilepsy, and Alzheimer’s Disease. | [80][81][82][83][84][85][86][87][88][89][90][91][92][93][94][95] |
2. Cytomegalovirus
3. Enteroviruses
4. Epstein-Barr Virus
5. Herpes Simplex Virus
6. Lymphocytic Choriomeningitis Virus
7. Severe Acute Respiratory Syndrome Coronavirus-2
References
- Watanabe, M.; Nishikawaji, Y.; Kawakami, H.; Kosai, K. Adenovirus Biology, Recombinant Adenovirus, and Adenovirus Usage in Gene Therapy. Viruses 2021, 13, 2502.
- Saban, S.D.; Silvestry, M.; Nemerow, G.R.; Stewart, P.L. Visualization of α-Helices in a 6-Ångstrom Resolution Cryoelectron Microscopy Structure of Adenovirus Allows Refinement of Capsid Protein Assignments. J. Virol. 2006, 80, 12049–12059.
- Davison, A.J.; Benkő, M.; Harrach, B. Genetic Content and Evolution of Adenoviruses. J. Gen. Virol. 2003, 84, 2895–2908.
- Smith, J.G.; Wiethoff, C.M.; Stewart, P.L.; Nemerow, G.R. Adenovirus. In Current topics in microbiology and immunology; Springer: New York, NY, USA, 2010; Volume 343, pp. 195–224.
- Rosa-Calatrava, M.; Grave, L.; Puvion-Dutilleul, F.; Chatton, B.; Kedinger, C. Functional Analysis of Adenovirus Protein IX Identifies Domains Involved in Capsid Stability, Transcriptional Activity, and Nuclear Reorganization. J. Virol. 2001, 75, 7131–7141.
- Moyer, C.L.; Besser, E.S.; Nemerow, G.R. A Single Maturation Cleavage Site in Adenovirus Impacts Cell Entry and Capsid Assembly. J. Virol. 2016, 90, 521–532.
- San Martín, C. Latest Insights on Adenovirus Structure and Assembly. Viruses 2012, 4, 847–877.
- McBride, M.; Williman, J.; Best, E.; Walls, T.; Sadarangani, M.; Grant, C.C.; Martin, N.G. The Epidemiology of Aseptic Meningitis in New Zealand Children from 1991 to 2020. J Paediatr. Child Health 2022, 58, 1980–1989.
- Mathew, S.; Al Khatib, H.A.; Al Ansari, K.; Nader, J.; Nasrallah, G.K.; Younes, N.N.; Coyle, P.V.; Al Thani, A.A.; Al Maslamani, M.A.; Yassine, H.M. Epidemiology Profile of Viral Meningitis Infections Among Patients in Qatar (2015–2018). Front. Med. 2021, 8, 663694.
- van Riel, D.; Verdijk, R.; Kuiken, T. The Olfactory Nerve: A Shortcut for Influenza and Other Viral Diseases into the Central Nervous System. J. Pathol. 2015, 235, 277–287.
- Hou, J.; Baker, L.A.; Zhou, L.; Klein, R.S. Viral Interactions with the Blood-Brain Barrier: Old Dog, New Tricks. Tissue Barriers 2016, 4, e1142492.
- Huang, Y.-C.; Huang, S.-L.; Chen, S.-P.; Huang, Y.-L.; Huang, C.-G.; Tsao, K.-C.; Lin, T.-Y. Adenovirus Infection Associated with Central Nervous System Dysfunction in Children. J. Clin. Virol. 2013, 57, 300–304.
- Fonseca, G.J.; Thillainadesan, G.; Yousef, A.F.; Ablack, J.N.; Mossman, K.L.; Torchia, J.; Mymryk, J.S. Adenovirus Evasion of Interferon-Mediated Innate Immunity by Direct Antagonism of a Cellular Histone Posttranslational Modification. Cell Host Microbe 2012, 11, 597–606.
- Sohn, S.-Y.; Hearing, P. Adenovirus Sequesters Phosphorylated STAT1 at Viral Replication Centers and Inhibits STAT Dephosphorylation. J. Virol. 2011, 85, 7555–7562.
- Lu, Q.; Yu, D.-H.; Fang, C.; Liu, F.; Ye, X.; Zhao, Y.; Qin, J.; Guo, X.-K.; Liang, M.; Hu, F.; et al. Influence of E3 Region on Conditionally Replicative Adenovirus Mediated Cytotoxicity in Hepatocellular Carcinoma Cells. Cancer Biol. Ther. 2009, 8, 1125–1132.
- Sester, M.; Koebernick, K.; Owen, D.; Ao, M.; Bromberg, Y.; May, E.; Stock, E.; Andrews, L.; Groh, V.; Spies, T.; et al. Conserved Amino Acids within the Adenovirus 2 E3/19K Protein Differentially Affect Downregulation of MHC Class I and MICA/B Proteins. J. Immunol. 2010, 184, 255–267.
- Van Damme, E.; Van Loock, M. Functional Annotation of Human Cytomegalovirus Gene Products: An Update. Front. Microbiol. 2014, 5, 218.
- Zeng, J.; Cao, D.; Yang, S.; Jaijyan, D.K.; Liu, X.; Wu, S.; Cruz-Cosme, R.; Tang, Q.; Zhu, H. Insights into the Transcriptome of Human Cytomegalovirus: A Comprehensive Review. Viruses 2023, 15, 1703.
- Kawasaki, H.; Kosugi, I.; Sakao-Suzuki, M.; Meguro, S.; Arai, Y.; Tsutsui, Y.; Iwashita, T. Cytomegalovirus Initiates Infection Selectively from High-Level Β1 Integrin–Expressing Cells in the Brain. Am. J. Pathol. 2015, 185, 1304–1323.
- Zheng, H.; Savitz, J. Effect of Cytomegalovirus Infection on the Central Nervous System: Implications for Psychiatric Disorders. In Microorganisms and Mental Health; Springer International Publishing: Cham, Switzerland, 2022; pp. 215–241.
- Sharma, S.; Mohan, A.; Smith-Rohrberg, D.; Sethu, M. Cytomegalovirus Polyradiculopathy: A Rare Neurological Manifestation of Acquired Immunodeficiency Syndrome. Neurol. India 2008, 56, 493.
- Kosugi, I.; Shinmura, Y.; Kawasaki, H.; Arai, Y.; Li, R.-Y.; Baba, S.; Tsutsui, Y. Cytomegalovirus Infection of the Central Nervous System Stem Cells from Mouse Embryo: A Model for Developmental Brain Disorders Induced by Cytomegalovirus. Lab. Investig. 2000, 80, 1373–1383.
- Grigoleit, U.; Riegler, S.; Einsele, H.; Laib Sampaio, K.; Jahn, G.; Hebart, H.; Brossart, P.; Frank, F.; Sinzger, C. Human Cytomegalovirus Induces a Direct Inhibitory Effect on Antigen Presentation by Monocyte-Derived Immature Dendritic Cells. Br. J. Haematol. 2002, 119, 189–198.
- Baasch, S.; Giansanti, P.; Kolter, J.; Riedl, A.; Forde, A.J.; Runge, S.; Zenke, S.; Elling, R.; Halenius, A.; Brabletz, S.; et al. Cytomegalovirus Subverts Macrophage Identity. Cell 2021, 184, 3774–3793.e25.
- LoPiccolo, D.M.; Gold, M.C.; Kavanagh, D.G.; Wagner, M.; Koszinowski, U.H.; Hill, A.B. Effective Inhibition of Kb- and Db-Restricted Antigen Presentation in Primary Macrophages by Murine Cytomegalovirus. J. Virol. 2003, 77, 301–308.
- Tabor-Godwin, J.M.; Ruller, C.M.; Bagalso, N.; An, N.; Pagarigan, R.R.; Harkins, S.; Gilbert, P.E.; Kiosses, W.B.; Gude, N.A.; Cornell, C.T.; et al. A Novel Population of Myeloid Cells Responding to Coxsackievirus Infection Assists in the Dissemination of Virus within the Neonatal CNS. J. Neurosci. 2010, 30, 8676–8691.
- Baicus, A.; Baicus, C. Neuronal Infections; Singh, S.K., Ruzek, D., Eds.; CRC Press: Boca Raton, FL, USA, 2013.
- Lei, X.; Liu, X.; Ma, Y.; Sun, Z.; Yang, Y.; Jin, Q.; He, B.; Wang, J. The 3C Protein of Enterovirus 71 Inhibits Retinoid Acid-Inducible Gene I-Mediated Interferon Regulatory Factor 3 Activation and Type I Interferon Responses. J. Virol. 2010, 84, 8051–8061.
- Coyne, C.B.; Bozym, R.; Morosky, S.A.; Hanna, S.L.; Mukherjee, A.; Tudor, M.; Kim, K.S.; Cherry, S. Comparative RNAi Screening Reveals Host Factors Involved in Enterovirus Infection of Polarized Endothelial Monolayers. Cell Host Microbe 2011, 9, 70–82.
- Wang, W.; Sun, J.; Wang, N.; Sun, Z.; Ma, Q.; Li, J.; Zhang, M.; Xu, J. Enterovirus A71 Capsid Protein VP1 Increases Blood–Brain Barrier Permeability and Virus Receptor Vimentin on the Brain Endothelial Cells. J. Neurovirol. 2020, 26, 84–94.
- Lim, Z.Q.; Ng, Q.Y.; Oo, Y.; Chu, J.J.H.; Ng, S.Y.; Sze, S.K.; Alonso, S. Enterovirus-A71 Exploits Peripherin and Rac1 to Invade the Central Nervous System. EMBO Rep. 2021, 22, e51777.
- Piralla, A.; Pellegrinelli, L.; Giardina, F.; Galli, C.; Binda, S.; Pariani, E.; Baldanti, F. Contribution of Enteroviruses to Acute Central Nervous System or Systemic Infections in Northern Italy (2015–2017): Is It Time to Establish a National Laboratory-Based Surveillance System? BioMed Res. Int. 2020, 2020, 9393264.
- Lai, R.-H.; Chow, Y.-H.; Chung, N.-H.; Chen, T.-C.; Shie, F.-S.; Juang, J.-L. Neurotropic EV71 Causes Encephalitis by Engaging Intracellular TLR9 to Elicit Neurotoxic IL12-P40-INOS Signaling. Cell Death Dis. 2022, 13, 328.
- de Ceano-Vivas, M.; García, M.L.; Velázquez, A.; Martín del Valle, F.; Menasalvas, A.; Cilla, A.; Epalza, C.; Romero, M.P.; Cabrerizo, M.; Calvo, C. Neurodevelopmental Outcomes of Infants Younger Than 90 Days Old Following Enterovirus and Parechovirus Infections of the Central Nervous System. Front. Pediatr. 2021, 9, 719119.
- Chang, L.-Y.; Huang, L.-M.; Gau, S.S.-F.; Wu, Y.-Y.; Hsia, S.-H.; Fan, T.-Y.; Lin, K.-L.; Huang, Y.-C.; Lu, C.-Y.; Lin, T.-Y. Neurodevelopment and Cognition in Children after Enterovirus 71 Infection. N. Engl. J. Med. 2007, 356, 1226–1234.
- Tseng, J.-J.; Lin, C.-H.; Lin, M.-C. Long-Term Outcomes of Pediatric Enterovirus Infection in Taiwan: A Population-Based Cohort Study. Front. Pediatr. 2020, 8, 285.
- Wang, H.; Lei, X.; Xiao, X.; Yang, C.; Lu, W.; Huang, Z.; Leng, Q.; Jin, Q.; He, B.; Meng, G.; et al. Reciprocal Regulation between Enterovirus 71 and the NLRP3 Inflammasome. Cell Rep. 2015, 12, 42–48.
- Xiang, Z.; Liu, L.; Lei, X.; Zhou, Z.; He, B.; Wang, J. 3C Protease of Enterovirus D68 Inhibits Cellular Defense Mediated by Interferon Regulatory Factor 7. J. Virol. 2016, 90, 1613–1621.
- Kang, J.; Pang, Z.; Zhou, Z.; Li, X.; Liu, S.; Cheng, J.; Liu, P.; Tan, W.; Wang, Z.; Wang, T. Enterovirus D68 Protease 2A pro Targets TRAF3 To Subvert Host Innate Immune Responses. J. Virol. 2021, 95, 10–1128.
- Young, L.S. Epstein–Barr Virus: General Features. In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 2008; pp. 148–157.
- Indari, O.; Chandramohanadas, R.; Jha, H.C. Epstein–Barr Virus Infection Modulates Blood–Brain Barrier Cells and Its Co-Infection with Plasmodium falciparum Induces RBC Adhesion. Pathog. Dis. 2021, 79, ftaa080.
- Williams, M.V.; Cox, B.; Lafuse, W.P.; Ariza, M.E. Epstein-Barr Virus DUTPase Induces Neuroinflammatory Mediators: Implications for Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Clin. Ther. 2019, 41, 848–863.
- Wallace, L.A.; English, S.W.; Fugate, J.E.; Tosh, P.K. Acute Epstein-Barr Virus Infection Presenting as Guillain-Barre Syndrome. IDCases 2021, 25, e01196.
- Singhi, P.; Sharma, J.P.; Gautam, R.; Indra, R.M.; Rafli, A. Extensive Longitudinal Transverse Myelitis Associated with CSF Epstein-Barr Virus Infection: A Case Report. Child. Neurol. Open 2021, 8, 2329048X2110499.
- Tiwari, D.; Singh, V.K.; Baral, B.; Pathak, D.K.; Jayabalan, J.; Kumar, R.; Tapryal, S.; Jha, H.C. Indication of Neurodegenerative Cascade Initiation by Amyloid-like Aggregate-Forming EBV Proteins and Peptide in Alzheimer’s Disease. ACS Chem. Neurosci. 2021, 12, 3957–3967.
- Woulfe, J.M.; Gray, M.T.; Gray, D.A.; Munoz, D.G.; Middeldorp, J.M. Hypothesis: A Role for EBV-Induced Molecular Mimicry in Parkinson’s Disease. Park. Relat. Disord. 2014, 20, 685–694.
- Yiu, S.P.T.; Zerbe, C.; Vanderwall, D.; Huttlin, E.L.; Weekes, M.P.; Gewurz, B.E. An Epstein-Barr Virus Protein Interaction Map Reveals NLRP3 Inflammasome Evasion via MAVS UFMylation. Mol. Cell 2023, 83, 2367–2386.e15.
- Liu, X.; Sadaoka, T.; Krogmann, T.; Cohen, J.I. Epstein-Barr Virus (EBV) Tegument Protein BGLF2 Suppresses Type I Interferon Signaling To Promote EBV Reactivation. J. Virol. 2020, 94, 10–1128.
- Wang, P.; Deng, Y.; Guo, Y.; Xu, Z.; Li, Y.; Ou, X.; Xie, L.; Lu, M.; Zhong, J.; Li, B.; et al. Epstein-Barr Virus Early Protein BFRF1 Suppresses IFN-β Activity by Inhibiting the Activation of IRF3. Front. Immunol. 2020, 11, 513383.
- Li, L.; Liu, D.; Hutt-Fletcher, L.; Morgan, A.; Masucci, M.G.; Levitsky, V. Epstein-Barr Virus Inhibits the Development of Dendritic Cells by Promoting Apoptosis of Their Monocyte Precursors in the Presence of Granulocyte Macrophage–Colony-Stimulating Factor and Interleukin-4. Blood 2002, 99, 3725–3734.
- Mrozek-Gorska, P.; Buschle, A.; Pich, D.; Schwarzmayr, T.; Fechtner, R.; Scialdone, A.; Hammerschmidt, W. Epstein–Barr Virus Reprograms Human B Lymphocytes Immediately in the Prelatent Phase of Infection. Proc. Natl. Acad. Sci. USA 2019, 116, 16046–16055.
- Megyola, C.; Ye, J.; Bhaduri-McIntosh, S. Identification of a Sub-Population of B Cells That Proliferates after Infection with Epstein-Barr Virus. Virol. J. 2011, 8, 84.
- Ma, N.; Lu, J.; Pei, Y.; Robertson, E.S. Transcriptome Reprogramming of Epstein-Barr Virus Infected Epithelial and B Cells Reveals Distinct Host-Virus Interaction Profiles. Cell Death Dis. 2022, 13, 894.
- Lehman, I.R.; Boehmer, P.E. Replication of Herpes Simplex Virus DNA. J. Biol. Chem. 1999, 274, 28059–28062.
- Lubinski, J.M.; Lazear, H.M.; Awasthi, S.; Wang, F.; Friedman, H.M. The Herpes Simplex Virus 1 IgG Fc Receptor Blocks Antibody-Mediated Complement Activation and Antibody-Dependent Cellular Cytotoxicity In Vivo. J. Virol. 2011, 85, 3239–3249.
- Komala Sari, T.; Gianopulos, K.A.; Nicola, A.V. Glycoprotein C of Herpes Simplex Virus 1 Shields Glycoprotein B from Antibody Neutralization. J. Virol. 2020, 94, 10–1128.
- Bello-Morales, R.; Andreu, S.; López-Guerrero, J.A. The Role of Herpes Simplex Virus Type 1 Infection in Demyelination of the Central Nervous System. Int. J. Mol. Sci. 2020, 21, 5026.
- Kopp, S.J.; Ranaivo, H.R.; Wilcox, D.R.; Karaba, A.H.; Wainwright, M.S.; Muller, W.J. Herpes Simplex Virus Serotype and Entry Receptor Availability Alter CNS Disease in a Mouse Model of Neonatal HSV. Pediatr. Res. 2014, 76, 528–534.
- He, Q.; Liu, H.; Huang, C.; Wang, R.; Luo, M.; Lu, W. Herpes Simplex Virus 1-Induced Blood-Brain Barrier Damage Involves Apoptosis Associated With GM130-Mediated Golgi Stress. Front. Mol. Neurosci. 2020, 13, 2.
- Liu, H.; Qiu, K.; He, Q.; Lei, Q.; Lu, W. Mechanisms of Blood-Brain Barrier Disruption in Herpes Simplex Encephalitis. J. Neuroimmune Pharmacol. 2019, 14, 157–172.
- Rozenberg, F. Herpes Simplex Virus and Central Nervous System Infections: Encephalitis, Meningitis, Myelitis. Virologie 2020, 24, 283–294.
- Murphy, M.J.; Fani, L.; Ikram, M.K.; Ghanbari, M.; Ikram, M.A. Herpes Simplex Virus 1 and the Risk of Dementia: A Population-Based Study. Sci. Rep. 2021, 11, 8691.
- Ma, Y.; He, B. Recognition of Herpes Simplex Viruses: Toll-Like Receptors and Beyond. J. Mol. Biol. 2014, 426, 1133–1147.
- Finberg, R.W.; Knipe, D.M.; Kurt-Jones, E.A. Herpes Simplex Virus and Toll-Like Receptors. Viral Immunol. 2005, 18, 457–465.
- Stefanidou, M.; Ramos, I.; Mas Casullo, V.; Trépanier, J.B.; Rosenbaum, S.; Fernandez-Sesma, A.; Herold, B.C. Herpes Simplex Virus 2 (HSV-2) Prevents Dendritic Cell Maturation, Induces Apoptosis, and Triggers Release of Proinflammatory Cytokines: Potential Links to HSV-HIV Synergy. J. Virol. 2013, 87, 1443–1453.
- Smiley, J.R. Herpes Simplex Virus Virion Host Shutoff Protein: Immune Evasion Mediated by a Viral RNase? J. Virol. 2004, 78, 1063–1068.
- Koppers-Lalic, D.; Rijsewijk, F.A.M.; Verschuren, S.B.E.; van Gaans-van den Brink, J.A.M.; Neisig, A.; Ressing, M.E.; Neefjes, J.; Wiertz, E.J.H.J. The UL41-Encoded Virion Host Shutoff (Vhs) Protein and Vhs-Independent Mechanisms Are Responsible for down-Regulation of MHC Class I Molecules by Bovine Herpesvirus 1. J. Gen. Virol. 2001, 82, 2071–2081.
- Tigges, M.A.; Leng, S.; Johnson, D.C.; Burke, R.L. Human Herpes Simplex Virus (HSV)-Specific CD8+ CTL Clones Recognize HSV-2-Infected Fibroblasts after Treatment with IFN-Gamma or When Virion Host Shutoff Functions Are Disabled. J. Immunol. 1996, 156, 3901–3910.
- Koyanagi, N.; Kawaguchi, Y. Evasion of the Cell-Mediated Immune Response by Alphaherpesviruses. Viruses 2020, 12, 1354.
- Buchmeier, M.J.; de la Torre, J.C.; Peters, C.J. Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Williams & Wilkins: Philadelphia, PA, USA, 2013.
- Kang, S.S.; McGavern, D.B. Lymphocytic Choriomeningitis Infection of the Central Nervous System. Front. Biosci. 2008, 4529, 4529.
- Spindler, K.R.; Hsu, T.-H. Viral Disruption of the Blood–Brain Barrier. Trends Microbiol. 2012, 20, 282–290.
- Kim, J.V.; Kang, S.S.; Dustin, M.L.; McGavern, D.B. Myelomonocytic Cell Recruitment Causes Fatal CNS Vascular Injury during Acute Viral Meningitis. Nature 2009, 457, 191–195.
- Matullo, C.M.; O’Regan, K.J.; Hensley, H.; Curtis, M.; Rall, G.F. Lymphocytic Choriomeningitis Virus-Induced Mortality in Mice Is Triggered by Edema and Brain Herniation. J. Virol. 2010, 84, 312–320.
- Barton, L.L.; Mets, M.B.; Beauchamp, C.L. Lymphocytic Choriomeningitis Virus: Emerging Fetal Teratogen. Am. J. Obs. Gynecol. 2002, 187, 1715–1716.
- Sharma, K.; Singhapakdi, K.; Maertens, P. Echoencephalography of Encephalopathy Due to Congenital Lymphocytic Choriomeningitis Virus. J. Neuroimaging 2022, 32, 412–419.
- Wright, R.; Johnson, D.; Neumann, M.; Ksiazek, T.G.; Rollin, P.; Keech, R.V.; Bonthius, D.J.; Hitchon, P.; Grose, C.F.; Bell, W.E.; et al. Congenital Lymphocytic Choriomeningitis Virus Syndrome: A Disease That Mimics Congenital Toxoplasmosis or Cytomegalovirus Infection. Pediatrics 1997, 100, e9.
- Pythoud, C.; Rothenberger, S.; Martínez-Sobrido, L.; de la Torre, J.C.; Kunz, S. Lymphocytic Choriomeningitis Virus Differentially Affects the Virus-Induced Type I Interferon Response and Mitochondrial Apoptosis Mediated by RIG-I/MAVS. J. Virol. 2015, 89, 6240–6250.
- Pythoud, C.; Rodrigo, W.W.S.I.; Pasqual, G.; Rothenberger, S.; Martínez-Sobrido, L.; de la Torre, J.C.; Kunz, S. Arenavirus Nucleoprotein Targets Interferon Regulatory Factor-Activating Kinase IKKε. J. Virol. 2012, 86, 7728–7738.
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The Species Severe Acute Respiratory Syndrome-Related Coronavirus: Classifying 2019-NCoV and Naming It SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544.
- Pellett, P.E.; Mitra, S.; Holland, T.C. Basics of Virology. In Handbook of Clinical Neurology; Academic Press: Cambridge, MA, USA, 2014; pp. 45–66.
- Wang, W.; Zhou, Z.; Xiao, X.; Tian, Z.; Dong, X.; Wang, C.; Li, L.; Ren, L.; Lei, X.; Xiang, Z.; et al. SARS-CoV-2 Nsp12 Attenuates Type I Interferon Production by Inhibiting IRF3 Nuclear Translocation. Cell Mol. Immunol. 2021, 18, 945–953.
- Arshad, N.; Laurent-Rolle, M.; Ahmed, W.S.; Hsu, J.C.-C.; Mitchell, S.M.; Pawlak, J.; Sengupta, D.; Biswas, K.H.; Cresswell, P. SARS-CoV-2 Accessory Proteins ORF7a and ORF3a Use Distinct Mechanisms to down-Regulate MHC-I Surface Expression. Proc. Natl. Acad. Sci. USA 2023, 120, e2208525120.
- Li, X.; Hou, P.; Ma, W.; Wang, X.; Wang, H.; Yu, Z.; Chang, H.; Wang, T.; Jin, S.; Wang, X.; et al. SARS-CoV-2 ORF10 Suppresses the Antiviral Innate Immune Response by Degrading MAVS through Mitophagy. Cell Mol. Immunol. 2022, 19, 67–78.
- Zhang, L.; Zhou, L.; Bao, L.; Liu, J.; Zhu, H.; Lv, Q.; Liu, R.; Chen, W.; Tong, W.; Wei, Q.; et al. SARS-CoV-2 Crosses the Blood–Brain Barrier Accompanied with Basement Membrane Disruption without Tight Junctions Alteration. Signal Transduct. Target. Ther. 2021, 6, 337.
- Buzhdygan, T.P.; DeOre, B.J.; Baldwin-Leclair, A.; Bullock, T.A.; McGary, H.M.; Khan, J.A.; Razmpour, R.; Hale, J.F.; Galie, P.A.; Potula, R.; et al. The SARS-CoV-2 Spike Protein Alters Barrier Function in 2D Static and 3D Microfluidic in-Vitro Models of the Human Blood–Brain Barrier. Neurobiol. Dis. 2020, 146, 105131.
- Vitale-Cross, L.; Szalayova, I.; Scoggins, A.; Palkovits, M.; Mezey, E. SARS-CoV-2 Entry Sites Are Present in All Structural Elements of the Human Glossopharyngeal and Vagal Nerves: Clinical Implications. EBioMedicine 2022, 78, 103981.
- Green, R.; Mayilsamy, K.; McGill, A.R.; Martinez, T.E.; Chandran, B.; Blair, L.J.; Bickford, P.C.; Mohapatra, S.S.; Mohapatra, S. SARS-CoV-2 Infection Increases the Gene Expression Profile for Alzheimer’s Disease Risk. Mol. Ther. Methods Clin. Dev. 2022, 27, 217–229.
- Vossler, D.G. Does SARS-CoV-2 Cause Seizures and Epilepsy in COVID-19 via Inflammation or by Direct Infection? Epilepsy Curr. 2023, 23, 153–155.
- Zhang, P.-P.; He, Z.-C.; Yao, X.-H.; Tang, R.; Ma, J.; Luo, T.; Zhu, C.; Li, T.-R.; Liu, X.; Zhang, D.; et al. COVID-19-Associated Monocytic Encephalitis (CAME): Histological and Proteomic Evidence from Autopsy. Signal Transduct. Target. Ther. 2023, 8, 24.
- Abenza Abildúa, M.J.; Atienza, S.; Carvalho Monteiro, G.; Erro Aguirre, M.E.; Imaz Aguayo, L.; Freire Álvarez, E.; García-Azorín, D.; Gil-Olarte Montesinos, I.; Lara Lezama, L.B.; Navarro Pérez, M.P.; et al. Encephalopathy and Encephalitis during Acute SARS-CoV-2 Infection. Spanish Society of Neurology’s COVID-19 Registry. Neurología 2021, 36, 127–134.
- Zheng, Y.; Deng, J.; Han, L.; Zhuang, M.-W.; Xu, Y.; Zhang, J.; Nan, M.-L.; Xiao, Y.; Zhan, P.; Liu, X.; et al. SARS-CoV-2 NSP5 and N Protein Counteract the RIG-I Signaling Pathway by Suppressing the Formation of Stress Granules. Signal Transduct. Target. Ther. 2022, 7, 22.
- Lu, X.; Pan, J.; Tao, J.; Guo, D. SARS-CoV Nucleocapsid Protein Antagonizes IFN-β Response by Targeting Initial Step of IFN-β Induction Pathway, and Its C-Terminal Region Is Critical for the Antagonism. Virus Genes. 2011, 42, 37–45.
- Chen, J.; Li, Z.; Guo, J.; Xu, S.; Zhou, J.; Chen, Q.; Tong, X.; Wang, D.; Peng, G.; Fang, L.; et al. SARS-CoV-2 Nsp5 Exhibits Stronger Catalytic Activity and Interferon Antagonism than Its SARS-CoV Ortholog. J. Virol. 2022, 96, e00037-22.
- López-Muñoz, A.D.; Kosik, I.; Holly, J.; Yewdell, J.W. Cell Surface SARS-CoV-2 Nucleocapsid Protein Modulates Innate and Adaptive Immunity. Sci. Adv. 2022, 8, eabp9770.
- Lynch, J.; Kajon, A. Adenovirus: Epidemiology, Global Spread of Novel Serotypes, and Advances in Treatment and Prevention. Semin. Respir. Crit. Care Med. 2016, 37, 586–602.
- Zhang, X.-F.; Tan, C.-B.; Yao, Z.-X.; Jiang, L.; Hong, S.-Q. Adenovirus Infection-Associated Central Nervous System Disease in Children. Pediatr. Infect. Dis. J. 2021, 40, 205–208.
- Schwartz, K.L.; Richardson, S.E.; MacGregor, D.; Mahant, S.; Raghuram, K.; Bitnun, A. Adenovirus-Associated Central Nervous System Disease in Children. J. Pediatr. 2019, 205, 130–137.
- Yamada, M.; Nakamura, K.; Yoshii, M.; Kaku, Y.; Narita, M. Brain Lesions Induced by Experimental Intranasal Infection of Japanese Encephalitis Virus in Piglets. J. Comp. Pathol. 2009, 141, 156–162.
- Ginsberg, H.S.; Moldawer, L.L.; Sehgal, P.B.; Redington, M.; Kilian, P.L.; Chanock, R.M.; Prince, G.A. A Mouse Model for Investigating the Molecular Pathogenesis of Adenovirus Pneumonia. Proc. Natl. Acad. Sci. USA 1991, 88, 1651–1655.
- Carlin, C.R. New Insights to Adenovirus-Directed Innate Immunity in Respiratory Epithelial Cells. Microorganisms 2019, 7, 216.
- Kotha, P.L.N.; Sharma, P.; Kolawole, A.O.; Yan, R.; Alghamri, M.S.; Brockman, T.L.; Gomez-Cambronero, J.; Excoffon, K.J.D.A. Adenovirus Entry From the Apical Surface of Polarized Epithelia Is Facilitated by the Host Innate Immune Response. PLoS Pathog. 2015, 11, e1004696.
- Bhat, N.R.; Fan, F. Adenovirus Infection Induces Microglial Activation: Involvement of Mitogen-Activated Protein Kinase Pathways. Brain Res. 2002, 948, 93–101.
- Zemke, N.R.; Berk, A.J. The Adenovirus E1A C Terminus Suppresses a Delayed Antiviral Response and Modulates RAS Signaling. Cell Host Microbe 2017, 22, 789–800.e5.
- Yunis, J.; Farrell, H.E.; Bruce, K.; Lawler, C.; Wyer, O.; Davis-Poynter, N.; Brizić, I.; Jonjić, S.; Adler, B.; Stevenson, P.G. Murine Cytomegalovirus Glycoprotein O Promotes Epithelial Cell Infection In Vivo. J. Virol. 2019, 93, e01378-18.
- Lopper, M.; Compton, T. Coiled-Coil Domains in Glycoproteins B and H Are Involved in Human Cytomegalovirus Membrane Fusion. J. Virol. 2004, 78, 8333–8341.
- Cosman, D.; Müllberg, J.; Sutherland, C.L.; Chin, W.; Armitage, R.; Fanslow, W.; Kubin, M.; Chalupny, N.J. ULBPs, Novel MHC Class I–Related Molecules, Bind to CMV Glycoprotein UL16 and Stimulate NK Cytotoxicity through the NKG2D Receptor. Immunity 2001, 14, 123–133.
- Ross, S.A.; Pati, P.; Jensen, T.L.; Goll, J.B.; Gelber, C.E.; Singh, A.; McNeal, M.; Boppana, S.B.; Bernstein, D.I. Cytomegalovirus Genetic Diversity Following Primary Infection. J. Infect. Dis. 2020, 221, 715–720.
- Dunn, W.; Chou, C.; Li, H.; Hai, R.; Patterson, D.; Stolc, V.; Zhu, H.; Liu, F. Functional Profiling of a Human Cytomegalovirus Genome. Proc. Natl. Acad. Sci. USA 2003, 100, 14223–14228.
- Zuhair, M.; Smit, G.S.A.; Wallis, G.; Jabbar, F.; Smith, C.; Devleesschauwer, B.; Griffiths, P. Estimation of the Worldwide Seroprevalence of Cytomegalovirus: A Systematic Review and Meta-analysis. Rev. Med. Virol. 2019, 29, e2034.
- Anduze-Faris, B.M.; Fillet, A.-M.; Gozlan, J.; Lancar, R.; Boukli, N.; Gasnault, J.; Caumes, E.; Livartowsky, J.; Matheron, S.; Leport, C.; et al. Induction and Maintenance Therapy of Cytomegalovirus Central Nervous System Infection in HIV-Infected Patients. AIDS 2000, 14, 517–524.
- Krstanović, F.; Britt, W.J.; Jonjić, S.; Brizić, I. Cytomegalovirus Infection and Inflammation in Developing Brain. Viruses 2021, 13, 1078.
- Slavuljica, I.; Kveštak, D.; Csaba Huszthy, P.; Kosmac, K.; Britt, W.J.; Jonjić, S. Immunobiology of Congenital Cytomegalovirus Infection of the Central Nervous System—The Murine Cytomegalovirus Model. Cell Mol. Immunol. 2015, 12, 180–191.
- Boehme, K.W.; Singh, J.; Perry, S.T.; Compton, T. Human Cytomegalovirus Elicits a Coordinated Cellular Antiviral Response via Envelope Glycoprotein B. J. Virol. 2004, 78, 1202–1211.
- Yurochko, A.D.; Hwang, E.S.; Rasmussen, L.; Keay, S.; Pereira, L.; Huang, E.S. The Human Cytomegalovirus UL55 (GB) and UL75 (GH) Glycoprotein Ligands Initiate the Rapid Activation of Sp1 and NF-KappaB during Infection. J. Virol. 1997, 71, 5051–5059.
- Ashley, C.; Abendroth, A.; McSharry, B.; Slobedman, B. Interferon-Independent Upregulation of Interferon-Stimulated Genes during Human Cytomegalovirus Infection Is Dependent on IRF3 Expression. Viruses 2019, 11, 246.
- Billstrom Schroeder, M.; Worthen, G.S. Viral Regulation of RANTES Expression during Human Cytomegalovirus Infection of Endothelial Cells. J. Virol. 2001, 75, 3383–3390.
- Browne, E.P.; Wing, B.; Coleman, D.; Shenk, T. Altered Cellular MRNA Levels in Human Cytomegalovirus-Infected Fibroblasts: Viral Block to the Accumulation of Antiviral MRNAs. J. Virol. 2001, 75, 12319–12330.
- Simmen, K.A.; Singh, J.; Luukkonen, B.G.M.; Lopper, M.; Bittner, A.; Miller, N.E.; Jackson, M.R.; Compton, T.; Früh, K. Global Modulation of Cellular Transcription by Human Cytomegalovirus Is Initiated by Viral Glycoprotein B. Proc. Natl. Acad. Sci. USA 2001, 98, 7140–7145.
- Maxim C-J Cheeran, S.H.S. Cytomegalovirus Induces Cytokine and Chemokine Production Differentially in Microglia and Astrocytes: Antiviral Implications. J. Neurovirol 2001, 7, 135–147.
- Füzik, T.; Moravcová, J.; Kalynych, S.; Plevka, P. Structure of Human Enterovirus 70 and Its Inhibition by Capsid-Binding Compounds. J. Virol. 2022, 96, e00604-22.
- Lee, K.-M.; Wu, C.-C.; Wu, S.-E.; Lin, Y.-H.; Wang, L.-T.; Chang, C.-R.; Huang, P.-N.; Shih, S.-R.; Kuo, R.-L. The RNA-Dependent RNA Polymerase of Enterovirus A71 Associates with Ribosomal Proteins and Positively Regulates Protein Translation. RNA Biol. 2020, 17, 608–622.
- Xiao, X.; Lei, X.; Zhang, Z.; Ma, Y.; Qi, J.; Wu, C.; Xiao, Y.; Li, L.; He, B.; Wang, J. Enterovirus 3A Facilitates Viral Replication by Promoting Phosphatidylinositol 4-Kinase IIIβ–ACBD3 Interaction. J. Virol. 2017, 91, 10–1128.
- Visser, L.J.; Langereis, M.A.; Rabouw, H.H.; Wahedi, M.; Muntjewerff, E.M.; de Groot, R.J.; van Kuppeveld, F.J.M. Essential Role of Enterovirus 2A Protease in Counteracting Stress Granule Formation and the Induction of Type I Interferon. J. Virol. 2019, 93, e00222-19.
- Brouwer, L.; Moreni, G.; Wolthers, K.C.; Pajkrt, D. World-Wide Prevalence and Genotype Distribution of Enteroviruses. Viruses 2021, 13, 434.
- Chen, B.-S.; Lee, H.-C.; Lee, K.-M.; Gong, Y.-N.; Shih, S.-R. Enterovirus and Encephalitis. Front. Microbiol. 2020, 11, 261.
- Feng, M.; Guo, S.; Fan, S.; Zeng, X.; Zhang, Y.; Liao, Y.; Wang, J.; Zhao, T.; Wang, L.; Che, Y.; et al. The Preferential Infection of Astrocytes by Enterovirus 71 Plays a Key Role in the Viral Neurogenic Pathogenesis. Front. Cell Infect. Microbiol. 2016, 6, 192.
- Poelaert, K.C.K.; van Kleef, R.G.D.M.; Liu, M.; van Vliet, A.; Lyoo, H.; Gerber, L.-S.; Narimatsu, Y.; Büll, C.; Clausen, H.; de Vries, E.; et al. Enterovirus D-68 Infection of Primary Rat Cortical Neurons: Entry, Replication, and Functional Consequences. mBio 2023, 14, e00245-23.
- Ye, N.; Gong, X.; Pang, L.; Gao, W.; Zhang, Y.; Li, X.; Liu, N.; Li, D.; Jin, Y.; Duan, Z. Cytokine Responses and Correlations Thereof with Clinical Profiles in Children with Enterovirus 71 Infections. BMC Infect. Dis. 2015, 15, 225.
- Gong, X.; Zhou, J.; Zhu, W.; Liu, N.; Li, J.; Li, L.; Jin, Y.; Duan, Z. Excessive Proinflammatory Cytokine and Chemokine Responses of Human Monocyte-Derived Macrophages to Enterovirus 71 Infection. BMC Infect. Dis. 2012, 12, 224.
- Chang, C.; Li, J.; Ou, Y.; Chen, W.; Liao, S.; Raung, S.; Hsiao, A.; Chen, C. Enterovirus 71 Infection Caused Neuronal Cell Death and Cytokine Expression in Cultured Rat Neural Cells. IUBMB Life 2015, 67, 789–800.
- Sugimoto, A.; Yamashita, Y.; Kanda, T.; Murata, T.; Tsurumi, T. Epstein-Barr Virus Genome Packaging Factors Accumulate in BMRF1-Cores within Viral Replication Compartments. PLoS ONE 2019, 14, e0222519.
- Henson, B.W.; Perkins, E.M.; Cothran, J.E.; Desai, P. Self-Assembly of Epstein-Barr Virus Capsids. J. Virol. 2009, 83, 3877–3890.
- Zuo, J.; Currin, A.; Griffin, B.D.; Shannon-Lowe, C.; Thomas, W.A.; Ressing, M.E.; Wiertz, E.J.H.J.; Rowe, M. The Epstein-Barr Virus G-Protein-Coupled Receptor Contributes to Immune Evasion by Targeting MHC Class I Molecules for Degradation. PLoS Pathog. 2009, 5, e1000255.
- Walston, J.J.; Hayman, I.R.; Gore, M.; Ferguson, M.; Temple, R.M.; Liao, J.; Alam, S.; Meyers, C.; Tugizov, S.M.; Hutt-Fletcher, L.; et al. The Epstein-Barr Virus Glycoprotein BDLF2 Is Essential for Efficient Viral Spread in Stratified Epithelium. J. Virol. 2023, 97, e01528-22.
- Chen, J.; Zhang, X.; Jardetzky, T.S.; Longnecker, R. The Epstein-Barr Virus (EBV) Glycoprotein B Cytoplasmic C-Terminal Tail Domain Regulates the Energy Requirement for EBV-Induced Membrane Fusion. J. Virol. 2014, 88, 11686–11695.
- Sathiyamoorthy, K.; Hu, Y.X.; Möhl, B.S.; Chen, J.; Longnecker, R.; Jardetzky, T.S. Structural Basis for Epstein–Barr Virus Host Cell Tropism Mediated by Gp42 and GHgL Entry Glycoproteins. Nat. Commun. 2016, 7, 13557.
- Watanabe, T.; Sakaida, K.; Yoshida, M.; Al Masud, H.M.A.; Sato, Y.; Goshima, F.; Kimura, H.; Murata, T. The C-Terminus of Epstein-Barr Virus BRRF2 Is Required for Its Proper Localization and Efficient Virus Production. Front. Microbiol. 2017, 8, 125.
- Chen, J.; Lu, Z.; Gong, W.; Xiao, X.; Feng, X.; Li, W.; Shan, S.; Xu, D.; Zhou, Z. Epstein-Barr Virus Protein BKRF4 Restricts Nucleosome Assembly to Suppress Host Antiviral Responses. Proc. Natl. Acad. Sci. USA 2022, 119, e2203782119.
- Hung, C.-H.; Chiu, Y.-F.; Wang, W.-H.; Chen, L.-W.; Chang, P.-J.; Huang, T.-Y.; Lin, Y.-J.; Tsai, W.-J.; Yang, C.-C. Interaction Between BGLF2 and BBLF1 Is Required for the Efficient Production of Infectious Epstein–Barr Virus Particles. Front. Microbiol. 2020, 10, 3021.
- Al Masud, H.M.A.; Yanagi, Y.; Watanabe, T.; Sato, Y.; Kimura, H.; Murata, T. Epstein-Barr Virus BBRF2 Is Required for Maximum Infectivity. Microorganisms 2019, 7, 705.
- Uddin, M.K.; Watanabe, T.; Arata, M.; Sato, Y.; Kimura, H.; Murata, T. Epstein-Barr Virus BBLF1 Mediates Secretory Vesicle Transport to Facilitate Mature Virion Release. J. Virol. 2023, 97, e00437-23.
- Balfour, H.H.; Sifakis, F.; Sliman, J.A.; Knight, J.A.; Schmeling, D.O.; Thomas, W. Age-Specific Prevalence of Epstein–Barr Virus Infection Among Individuals Aged 6–19 Years in the United States and Factors Affecting Its Acquisition. J. Infect. Dis. 2013, 208, 1286–1293.
- Sharifipour, S.; Davoodi Rad, K. Seroprevalence of Epstein–Barr Virus among Children and Adults in Tehran, Iran. New Microbes New Infect. 2020, 34, 100641.
- Ye, Z.; Chen, L.; Zhong, H.; Cao, L.; Fu, P.; Xu, J. Epidemiology and Clinical Characteristics of Epstein-Barr Virus Infection among Children in Shanghai, China, 2017–2022. Front Cell Infect. Microbiol 2023, 13, 1139068.
- Jakhmola, S.; Jha, H.C. Glial Cell Response to Epstein-Barr Virus Infection: A Plausible Contribution to Virus-Associated Inflammatory Reactions in the Brain. Virology 2021, 559, 182–195.
- Jha, H.C.; Mehta, D.; Lu, J.; El-Naccache, D.; Shukla, S.K.; Kovacsics, C.; Kolson, D.; Robertson, E.S. Gammaherpesvirus Infection of Human Neuronal Cells. mBio 2015, 6, 10–1128.
- Cheng, H.; Chen, D.; Peng, X.; Wu, P.; Jiang, L.; Hu, Y. Clinical Characteristics of Epstein–Barr Virus Infection in the Pediatric Nervous System. BMC Infect. Dis. 2020, 20, 886.
- Johnson, D.B.; McDonnell, W.J.; Gonzalez-Ericsson, P.I.; Al-Rohil, R.N.; Mobley, B.C.; Salem, J.-E.; Wang, D.Y.; Sanchez, V.; Wang, Y.; Chastain, C.A.; et al. A Case Report of Clonal EBV-like Memory CD4+ T Cell Activation in Fatal Checkpoint Inhibitor-Induced Encephalitis. Nat. Med. 2019, 25, 1243–1250.
- D’Ambrosio, E.; Khalighinejad, F.; Ionete, C. Intravenous Immunoglobulins in an Adult Case of Post-EBV Cerebellitis. BMJ Case Rep. 2020, 13, e231661.
- Patra, P.; Rani, A.; Sharma, N.; Mukherjee, C.; Jha, H.C. Unraveling the Connection of Epstein–Barr Virus and Its Glycoprotein M 146–157 Peptide with Neurological Ailments. ACS Chem. Neurosci. 2023, 14, 2450–2460.
- Wada, T.; Muraoka, M.; Yokoyama, T.; Toma, T.; Kanegane, H.; Yachie, A. Cytokine Profiles in Children with Primary Epstein-Barr Virus Infection. Pediatr. Blood Cancer 2013, 60, E46–E48.
- Lui, W.-Y.; Bharti, A.; Wong, N.-H.M.; Jangra, S.; Botelho, M.G.; Yuen, K.-S.; Jin, D.-Y. Suppression of CGAS- and RIG-I-Mediated Innate Immune Signaling by Epstein-Barr Virus Deubiquitinase BPLF1. PLoS Pathog. 2023, 19, e1011186.
- Hooykaas, M.J.G.; van Gent, M.; Soppe, J.A.; Kruse, E.; Boer, I.G.J.; van Leenen, D.; Groot Koerkamp, M.J.A.; Holstege, F.C.P.; Ressing, M.E.; Wiertz, E.J.H.J.; et al. EBV MicroRNA BART16 Suppresses Type I IFN Signaling. J. Immunol. 2017, 198, 4062–4073.
- Mullen, M.M.; Haan, K.M.; Longnecker, R.; Jardetzky, T.S. Structure of the Epstein-Barr Virus Gp42 Protein Bound to the MHC Class II Receptor HLA-DR1. Mol. Cell 2002, 9, 375–385.
- Kamperschroer, C.; Quinn, D.G. The Role of Proinflammatory Cytokines in Wasting Disease During Lymphocytic Choriomeningitis Virus Infection. J. Immunol. 2002, 169, 340–349.
- James, C.; Harfouche, M.; Welton, N.J.; Turner, K.M.; Abu-Raddad, L.J.; Gottlieb, S.L.; Looker, K.J. Herpes Simplex Virus: Global Infection Prevalence and Incidence Estimates, 2016. Bull World Health Organ 2020, 98, 315–329.
- Feige, L.; Zaeck, L.M.; Sehl-Ewert, J.; Finke, S.; Bourhy, H. Innate Immune Signaling and Role of Glial Cells in Herpes Simplex Virus- and Rabies Virus-Induced Encephalitis. Viruses 2021, 13, 2364.
- Babaei, A.; Shatizadeh Malekshahi, S.; Pirbonyeh, N.; Moattari, A. Prevalence and Clinical Manifestations of Herpes Simplex Virus Infection among Suspected Patients of Herpes Simplex Encephalitis in Shiraz, Iran. Virusdisease 2021, 32, 266–271.
- Choi, R.; Kim, G.; Jo, I.J.; Sim, M.S.; Song, K.J.; Kim, B.J.; Na, D.L.; Huh, H.J.; Kim, J.; Ki, C.; et al. Incidence and Clinical Features of Herpes Simplex Viruses (1 and 2) and Varicella-zoster Virus Infections in an Adult Korean Population with Aseptic Meningitis or Encephalitis. J. Med. Virol. 2014, 86, 957–962.
- Xiao, T.S.; Fitzgerald, K.A. The CGAS-STING Pathway for DNA Sensing. Mol. Cell 2013, 51, 135–139.
- Unterholzner, L. The Interferon Response to Intracellular DNA: Why so Many Receptors? Immunobiology 2013, 218, 1312–1321.
- Thompson, M.R.; Kaminski, J.J.; Kurt-Jones, E.A.; Fitzgerald, K.A. Pattern Recognition Receptors and the Innate Immune Response to Viral Infection. Viruses 2011, 3, 920–940.
- Smith, J.B.; Herbert, J.J.; Truong, N.R.; Cunningham, A.L. Cytokines and Chemokines: The Vital Role They Play in Herpes Simplex Virus Mucosal Immunology. Front. Immunol. 2022, 13, 936235.
- Wang, J.P.; Bowen, G.N.; Zhou, S.; Cerny, A.; Zacharia, A.; Knipe, D.M.; Finberg, R.W.; Kurt-Jones, E.A. Role of Specific Innate Immune Responses in Herpes Simplex Virus Infection of the Central Nervous System. J. Virol. 2012, 86, 2273–2281.
- Kurt-Jones, E.A.; Chan, M.; Zhou, S.; Wang, J.; Reed, G.; Bronson, R.; Arnold, M.M.; Knipe, D.M.; Finberg, R.W. Herpes Simplex Virus 1 Interaction with Toll-like Receptor 2 Contributes to Lethal Encephalitis. Proc. Natl. Acad. Sci. USA 2004, 101, 1315–1320.
- Aravalli, R.N.; Hu, S.; Rowen, T.N.; Palmquist, J.M.; Lokensgard, J.R. Cutting Edge: TLR2-Mediated Proinflammatory Cytokine and Chemokine Production by Microglial Cells in Response to Herpes Simplex Virus. J. Immunol. 2005, 175, 4189–4193.
- Qiao, H.; Chiu, Y.; Liang, X.; Xia, S.; Ayrapetyan, M.; Liu, S.; He, C.; Song, R.; Zeng, J.; Deng, X.; et al. Microglia Innate Immune Response Contributes to the Antiviral Defense and Blood–CSF Barrier Function in Human Choroid Plexus Organoids during HSV-1 Infection. J. Med. Virol. 2023, 95, e28472.
- Michael, B.D.; Bricio-Moreno, L.; Sorensen, E.W.; Miyabe, Y.; Lian, J.; Solomon, T.; Kurt-Jones, E.A.; Luster, A.D. Astrocyte- and Neuron-Derived CXCL1 Drives Neutrophil Transmigration and Blood-Brain Barrier Permeability in Viral Encephalitis. Cell Rep. 2020, 32, 108150.
- Poppers, J.; Mulvey, M.; Khoo, D.; Mohr, I. Inhibition of PKR Activation by the Proline-Rich RNA Binding Domain of the Herpes Simplex Virus Type 1 Us11 Protein. J. Virol. 2000, 74, 11215–11221.
- Gobeil, P.A.M.; Leib, D.A. Herpes Simplex Virus Γ34.5 Interferes with Autophagosome Maturation and Antigen Presentation in Dendritic Cells. mBio 2012, 3, 10-1128.
- Trgovcich, J.; Johnson, D.; Roizman, B. Cell Surface Major Histocompatibility Complex Class II Proteins Are Regulated by the Products of the γ 1 34.5 and U L 41 Genes of Herpes Simplex Virus 1. J. Virol. 2002, 76, 6974–6986.
- Riviere, Y.; Ahmed, R.; Southern, P.J.; Buchmeier, M.J.; Dutko, F.J.; Oldstone, M.B. The S RNA Segment of Lymphocytic Choriomeningitis Virus Codes for the Nucleoprotein and Glycoproteins 1 and 2. J. Virol. 1985, 53, 966–968.
- Morin, B.; Coutard, B.; Lelke, M.; Ferron, F.; Kerber, R.; Jamal, S.; Frangeul, A.; Baronti, C.; Charrel, R.; de Lamballerie, X.; et al. The N-Terminal Domain of the Arenavirus L Protein Is an RNA Endonuclease Essential in MRNA Transcription. PLoS Pathog. 2010, 6, e1001038.
- Singh, M.K.; Fuller-Pace, F.V.; Buchmeier, M.J.; Southern, P.J. Analysis of the Genomic l Rna Segment from Lymphocytic Choriomeningitis Virus. Virology 1987, 161, 448–456.
- Charrel, R.N.; Coutard, B.; Baronti, C.; Canard, B.; Nougairede, A.; Frangeul, A.; Morin, B.; Jamal, S.; Schmidt, C.L.; Hilgenfeld, R.; et al. Arenaviruses and Hantaviruses: From Epidemiology and Genomics to Antivirals. Antivir. Res. 2011, 90, 102–114.
- Salvato, M.S.; Shimomaye, E.M. The Completed Sequence of Lymphocytic Choriomeningitis Virus Reveals a Unique RNA Structure and a Gene for a Zinc Finger Protein. Virology 1989, 173, 1–10.
- Ortiz-Riaño, E.; Cheng, B.Y.H.; de la Torre, J.C.; Martínez-Sobrido, L. The C-Terminal Region of Lymphocytic Choriomeningitis Virus Nucleoprotein Contains Distinct and Segregable Functional Domains Involved in NP-Z Interaction and Counteraction of the Type I Interferon Response. J. Virol. 2011, 85, 13038–13048.
- Martínez-Sobrido, L.; Zúñiga, E.I.; Rosario, D.; García-Sastre, A.; de la Torre, J.C. Inhibition of the Type I Interferon Response by the Nucleoprotein of the Prototypic Arenavirus Lymphocytic Choriomeningitis Virus. J. Virol. 2006, 80, 9192–9199.
- Pinschewer, D.D.; Perez, M.; de la Torre, J.C. Role of the Virus Nucleoprotein in the Regulation of Lymphocytic Choriomeningitis Virus Transcription and RNA Replication. J. Virol. 2003, 77, 3882–3887.
- Labudova, M.; Tomaskova, J.; Skultety, L.; Pastorek, J.; Pastorekova, S. The Nucleoprotein of Lymphocytic Choriomeningitis Virus Facilitates Spread of Persistent Infection through Stabilization of the Keratin Network. J. Virol. 2009, 83, 7842–7849.
- Hashizume, M.; Takashima, A.; Iwasaki, M. A Small Stem-Loop-Forming Region within the 3′-UTR of a Nonpolyadenylated LCMV MRNA Promotes Translation. J. Biol. Chem. 2022, 298, 101576.
- Igonet, S.; Vaney, M.-C.; Vonrhein, C.; Bricogne, G.; Stura, E.A.; Hengartner, H.; Eschli, B.; Rey, F.A. X-Ray Structure of the Arenavirus Glycoprotein GP2 in Its Postfusion Hairpin Conformation. Proc. Natl. Acad. Sci. USA 2011, 108, 19967–19972.
- Sullivan, B.M.; Emonet, S.F.; Welch, M.J.; Lee, A.M.; Campbell, K.P.; de la Torre, J.C.; Oldstone, M.B. Point Mutation in the Glycoprotein of Lymphocytic Choriomeningitis Virus Is Necessary for Receptor Binding, Dendritic Cell Infection, and Long-Term Persistence. Proc. Natl. Acad. Sci. USA 2011, 108, 2969–2974.
- Dykewicz, C.A. Lymphocytic Choriomeningitis Outbreak Associated With Nude Mice in a Research Institute. JAMA J. Am. Med. Assoc. 1992, 267, 1349.
- Vilibic-Cavlek, T.; Savic, V.; Ferenc, T.; Mrzljak, A.; Barbic, L.; Bogdanic, M.; Stevanovic, V.; Tabain, I.; Ferencak, I.; Zidovec-Lepej, S. Lymphocytic Choriomeningitis—Emerging Trends of a Neglected Virus: A Narrative Review. Trop. Med. Infect. Dis. 2021, 6, 88.
- Bonthius, D.J.; Mahoney, J.; Buchmeier, M.J.; Karacay, B.; Taggard, D. Critical Role for Glial Cells in the Propagation and Spread of Lymphocytic Choriomeningitis Virus in the Developing Rat Brain. J. Virol. 2002, 76, 6618–6635.
- Fox, J.G.; Otto, G.; Colby, L.A. Selected Zoonoses. In Laboratory Animal Medicine; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1313–1370.
- Puccini, J.M.; Ruller, C.M.; Robinson, S.M.; Knopp, K.A.; Buchmeier, M.J.; Doran, K.S.; Feuer, R. Distinct Neural Stem Cell Tropism, Early Immune Activation, and Choroid Plexus Pathology Following Coxsackievirus Infection in the Neonatal Central Nervous System. Lab. Investig. 2014, 94, 161–181.
- Bonthius, D.J. Lymphocytic Choriomeningitis Virus: An Underrecognized Cause of Neurologic Disease in the Fetus, Child, and Adult. Semin. Pediatr. Neurol. 2012, 19, 89–95.
- Pearce, B.D.; Po, C.L.; Pisell, T.L.; Miller, A.H. Lymphocytic Responses and the Gradual Hippocampal Neuron Loss Following Infection with Lymphocytic Choriomeningitis Virus (LCMV). J. Neuroimmunol. 1999, 101, 137–147.
- Zhou, S.; Kurt-Jones, E.A.; Mandell, L.; Cerny, A.; Chan, M.; Golenbock, D.T.; Finberg, R.W. MyD88 Is Critical for the Development of Innate and Adaptive Immunity during Acute Lymphocytic Choriomeningitis Virus Infection. Eur. J. Immunol. 2005, 35, 822–830.
- Pratumchai, I.; Zak, J.; Huang, Z.; Min, B.; Oldstone, M.B.A.; Teijaro, J.R. B Cell–Derived IL-27 Promotes Control of Persistent LCMV Infection. Proc. Natl. Acad. Sci. USA 2022, 119, e2116741119.
- Lin, A.A.; Wojciechowski, S.E.; Hildeman, D.A. Androgens Suppress Antigen-Specific T Cell Responses and IFN-γ Production during Intracranial LCMV Infection. J. Neuroimmunol. 2010, 226, 8–19.
- Che Mat, N.F.; Siddiqui, S.; Mehta, D.; Seaver, K.; Banete, A.; Alothaimeen, T.; Gee, K.; Basta, S. Lymphocytic Choriomeningitis Virus Infection of Dendritic Cells Interferes with TLR-Induced IL-12/IL-23 Cytokine Production in an IL-10 Independent Manner. Cytokine 2018, 108, 105–114.
- Zhou, S.; Halle, A.; Kurt-Jones, E.A.; Cerny, A.M.; Porpiglia, E.; Rogers, M.; Golenbock, D.T.; Finberg, R.W. Lymphocytic Choriomeningitis Virus (LCMV) Infection of CNS Glial Cells Results in TLR2-MyD88/Mal-Dependent Inflammatory Responses. J. Neuroimmunol. 2008, 194, 70–82.
- Khailany, R.A.; Safdar, M.; Ozaslan, M. Genomic Characterization of a Novel SARS-CoV-2. Gene Rep. 2020, 19, 100682.
- Hardenbrook, N.J.; Zhang, P. A Structural View of the SARS-CoV-2 Virus and Its Assembly. Curr. Opin. Virol. 2022, 52, 123–134.
- Boson, B.; Legros, V.; Zhou, B.; Siret, E.; Mathieu, C.; Cosset, F.-L.; Lavillette, D.; Denolly, S. The SARS-CoV-2 Envelope and Membrane Proteins Modulate Maturation and Retention of the Spike Protein, Allowing Assembly of Virus-like Particles. J. Biol. Chem. 2021, 296, 100111.
- Zhang, Z.; Nomura, N.; Muramoto, Y.; Ekimoto, T.; Uemura, T.; Liu, K.; Yui, M.; Kono, N.; Aoki, J.; Ikeguchi, M.; et al. Structure of SARS-CoV-2 Membrane Protein Essential for Virus Assembly. Nat. Commun. 2022, 13, 4399.
- Zaffagni, M.; Harris, J.M.; Patop, I.L.; Pamudurti, N.R.; Nguyen, S.; Kadener, S. SARS-CoV-2 Nsp14 Mediates the Effects of Viral Infection on the Host Cell Transcriptome. Elife 2022, 11, e71945.
- Angeletti, S.; Benvenuto, D.; Bianchi, M.; Giovanetti, M.; Pascarella, S.; Ciccozzi, M. COVID-2019: The Role of the Nsp2 and Nsp3 in Its Pathogenesis. J. Med. Virol. 2020, 92, 584–588.
- Konno, Y.; Kimura, I.; Uriu, K.; Fukushi, M.; Irie, T.; Koyanagi, Y.; Sauter, D.; Gifford, R.J.; Nakagawa, S.; Sato, K. SARS-CoV-2 ORF3b Is a Potent Interferon Antagonist Whose Activity Is Increased by a Naturally Occurring Elongation Variant. Cell Rep. 2020, 32, 108185.
- Miyamoto, Y.; Itoh, Y.; Suzuki, T.; Tanaka, T.; Sakai, Y.; Koido, M.; Hata, C.; Wang, C.-X.; Otani, M.; Moriishi, K.; et al. SARS-CoV-2 ORF6 Disrupts Nucleocytoplasmic Trafficking to Advance Viral Replication. Commun. Biol. 2022, 5, 483.
- Yang, R.; Zhao, Q.; Rao, J.; Zeng, F.; Yuan, S.; Ji, M.; Sun, X.; Li, J.; Yang, J.; Cui, J.; et al. SARS-CoV-2 Accessory Protein ORF7b Mediates Tumor Necrosis Factor-α-Induced Apoptosis in Cells. Front. Microbiol. 2021, 12, 654709.
- Han, L.; Zhuang, M.; Deng, J.; Zheng, Y.; Zhang, J.; Nan, M.; Zhang, X.; Gao, C.; Wang, P. SARS-CoV-2 ORF9b Antagonizes Type I and III Interferons by Targeting Multiple Components of the RIG-I/MDA-5–MAVS, TLR3–TRIF, and CGAS–STING Signaling Pathways. J. Med. Virol. 2021, 93, 5376–5389.
- Mena, G.E.; Martinez, P.P.; Mahmud, A.S.; Marquet, P.A.; Buckee, C.O.; Santillana, M. Socioeconomic Status Determines COVID-19 Incidence and Related Mortality in Santiago, Chile. Science 2021, 372, eabg5298.
- Balloff, C.; Bandlow, C.; Bernhard, M.; Brandenburger, T.; Bludau, P.; Elben, S.; Feldt, T.; Hartmann, C.J.; Heinen, E.; Ingwersen, J.; et al. Prevalence and Prognostic Value of Neurological Affections in Hospitalized Patients with Moderate to Severe COVID-19 Based on Objective Assessments. Sci. Rep. 2023, 13, 19619.
- Malik, J.R.; Acharya, A.; Avedissian, S.N.; Byrareddy, S.N.; Fletcher, C.V.; Podany, A.T.; Dyavar, S.R. ACE-2, TMPRSS2, and Neuropilin-1 Receptor Expression on Human Brain Astrocytes and Pericytes and SARS-CoV-2 Infection Kinetics. Int. J. Mol. Sci. 2023, 24, 8622.
- Crunfli, F.; Carregari, V.C.; Veras, F.P.; Silva, L.S.; Nogueira, M.H.; Antunes, A.S.L.M.; Vendramini, P.H.; Valença, A.G.F.; Brandão-Teles, C.; Zuccoli, G.d.S.; et al. Morphological, Cellular, and Molecular Basis of Brain Infection in COVID-19 Patients. Proc. Natl. Acad. Sci. USA 2022, 119, e2200960119.
- Martínez-Mármol, R.; Giordano-Santini, R.; Kaulich, E.; Cho, A.-N.; Przybyla, M.; Riyadh, M.A.; Robinson, E.; Chew, K.Y.; Amor, R.; Meunier, F.A.; et al. SARS-CoV-2 Infection and Viral Fusogens Cause Neuronal and Glial Fusion That Compromises Neuronal Activity. Sci. Adv. 2023, 9, eadg2248.
- Zhu, X.; Gebo, K.A.; Abraham, A.G.; Habtehyimer, F.; Patel, E.U.; Laeyendecker, O.; Gniadek, T.J.; Fernandez, R.E.; Baker, O.R.; Ram, M.; et al. Dynamics of Inflammatory Responses after SARS-CoV-2 Infection by Vaccination Status in the USA: A Prospective Cohort Study. Lancet Microbe 2023, 4, e692–e703.
- Pereson, M.J.; Badano, M.N.; Aloisi, N.; Chuit, R.E.; de Bracco, M.M.; Bare, P. TNF-α Levels in Respiratory Samples Are Associated with SARS-CoV-2 Infection. Microbiol. Spectr. 2022, 10, e01411-21.
- Neufeldt, C.J.; Cerikan, B.; Cortese, M.; Frankish, J.; Lee, J.-Y.; Plociennikowska, A.; Heigwer, F.; Prasad, V.; Joecks, S.; Burkart, S.S.; et al. SARS-CoV-2 Infection Induces a pro-Inflammatory Cytokine Response through CGAS-STING and NF-ΚB. Commun. Biol. 2022, 5, 45.
- Planès, R.; Bert, J.-B.; Tairi, S.; BenMohamed, L.; Bahraoui, E. SARS-CoV-2 Envelope (E) Protein Binds and Activates TLR2 Pathway: A Novel Molecular Target for COVID-19 Interventions. Viruses 2022, 14, 999.

