Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Ilya Kirov | -- | 1724 | 2023-12-05 15:49:34 | | | |
| 2 | Jessie Wu | + 3 word(s) | 1727 | 2023-12-06 06:21:47 | | | | |
| 3 | Jessie Wu | Meta information modification | 1727 | 2023-12-06 06:22:16 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Kirov, I. Genotype-Mediated Transposable Element Transposition Reactivation. Encyclopedia. Available online: https://encyclopedia.pub/entry/52394 (accessed on 24 June 2026).
Kirov I. Genotype-Mediated Transposable Element Transposition Reactivation. Encyclopedia. Available at: https://encyclopedia.pub/entry/52394. Accessed June 24, 2026.
Kirov, Ilya. "Genotype-Mediated Transposable Element Transposition Reactivation" Encyclopedia, https://encyclopedia.pub/entry/52394 (accessed June 24, 2026).
Kirov, I. (2023, December 05). Genotype-Mediated Transposable Element Transposition Reactivation. In Encyclopedia. https://encyclopedia.pub/entry/52394
Kirov, Ilya. "Genotype-Mediated Transposable Element Transposition Reactivation." Encyclopedia. Web. 05 December, 2023.
Copy Citation
Genetic diversity is a key factor for plant breeding. The birth of novel genic and genomic variants is also crucial for plant adaptation in nature. Therefore, the genomes of almost all living organisms possess natural mutagenic mechanisms. Transposable elements (TEs) are a major mutagenic force driving genetic diversity in wild plants and modern crops. The relatively rare TE transposition activity during the thousand-year crop domestication process has led to the phenotypic diversity of many cultivated species. The utilization of TE mutagenesis by artificial and transient acceleration of their activity in a controlled mode is an attractive foundation for a novel type of mutagenesis called TE-mediated biological mutagenesis.
transposons
plants
mutagenesis
RNA-dependent DNA methylation
gene silencing
1. Introduction
Plant breeding relies on the genetic diversity available to breeders, which is used to generate new combinations of genes and traits. Therefore, methods and approaches for generating genetic diversity in crops are the cornerstones of the plant breeding process. Implication of wild relatives and different genotypes and varieties in crosses followed by the selection of the most desirable genotypes from the progeny is a common way to create genetic diversity. Another approach involves chemical (e.g., EMS treatment) and radiation mutagenesis [1]. Mutagenesis has led to the development of new crop varieties and genetic resources [1]. Mutated genetic populations have been created for many plant species, leading to tremendous progress in functional genomics. However, radiation mutagenesis has several disadvantages, including a low frequency of valuable mutations and challenging identification of the causative mutations. However, novel techniques for precise gene targeting (e.g., genome editing) have been developed and adapted for many crop species [2][3]. Therefore, the application of classical mutagenesis in plant breeding has significantly decreased in recent decades [1].
Mutagenesis also occurs in nature but at a significantly lower rate than that achieved by artificial mutagenesis. It is obvious that in contrast to animals, plants cannot escape stress factors and cannot conquer new ecological niches by long-distance physical movement. Therefore, the main strategy for plants to adapt to new environmental conditions and stress is to explore their natural genetic diversity. It is now clear that environmental stress may increase the rate of single-nucleotide mutations [4] as well as structural rearrangements, including transposable element (TE) movements [5]. In the last few decades, a list of studies have concluded that stress may stimulate transposable elements (TEs) to transpose in the genome and to broaden genetic diversity [6][7][8]. Importantly, novel TE insertions (TEIs) are not randomly generated over the genome and may occur frequently in stress-responsive genes, providing connections between stress action and the occurrence of novel alleles of stress-responsive genes [9][10][11]. This indicates that TE transposition is an evolutionary mechanism that serves as a natural biological mutagen and an important source of genetic diversity for plant adaptation.
Interestingly, TE transposition is also a major source of crop diversity and has been widely used by plant breeders for hundreds of years [12][13]. The gene candidate approach revealed a number of cases in which TE transposition in or near the genes caused the origin of novel phenotypic variants in different species [6][10][13]. Modern genomics has provided tools to perform genome-wide analyses of the association between trait variation and TEIs. Indeed, recent genome-wide association studies (GWASs) exploiting small variants (for example, SNPs and InDels) and TEI polymorphisms (TIPs) have revealed the great impact of TIPs on crop domestication and breeding [13][14]. For example, Hopscotch TE insertion into the teosinte branched1 (tb1) gene is associated with increased apical dominance in modern maize [15]. Also, TE insertions were shown to play a key role in tomato domestication, causing changes in tomato leaf shape, scene, and fruit color [13]. Recently, the implication of TE insertions in the diversity of modern Brassica rapa morphotypes has been demonstrated [14]. Therefore, TEs can be considered one of the most effective biological mutagens that can be exploited by plant breeders. The most recent understanding of TE biology and TE suppression mechanisms (e.g., RNA-directed DNA methylation pathways and various small RNAs), combined with the most recent molecular tools (e.g., CRISPR-based technology and virus-induced gene silencing), could allow for the use of TE mutational potential in the control mode.
TE insertions may result in a broad spectrum of consequences at the genome, epigenetic, transcriptome, and phenotype levels [12][13][14][15][16][17][18]. Based on this, the idea that the controlled activation of native TEs in a plant genome can bring new TE-mediated mutagenesis technology to accelerate plant breeding has been proposed and discussed recently [11][12][19]. In recent years, the validity of this concept has been established through research on the model organism Arabidopsis thaliana [18][19][20][21][22], but studies on crops are still lagging behind.
2. Heterologous Expression of Transposable Elements via Plant Transformation
In 1983, the first active plant transposon, Ac, was cloned and sequenced from maize [23]. This began the era of TE application for the mutagenesis of plants via genetic transformation. In initial reports, the active TEs of one species were transformed into the genomes of other species to overcome host silencing mechanisms [24]. Using this approach, the transformed TEs were kept active in the foreign genome until they were recognized by the host silencing system. For example, Tos17 (rice), Tnt1, and TtoI (tobacco) retrotransposons were involved in gene tagging experiments in rice, Arabidopsis thaliana, Medicago truncatula, and other plant species [24][25][26][27][28]. It is important to note that these TEs are cell-culture-inducible, and therefore their activity can be tightly controlled or reactivated by a new round of in vitro regeneration [28]. Although heterologous expression of TEs plays an important role in understanding the TE transposition process and its silencing, the application of this approach for plant breeding is challenging in many countries because of the GMO legislation. Therefore, alternative approaches are required to establish GMO-free, breeder-friendly methods for TE-mediated mutagenesis.
3. Implication of DNA-Methylation-Deficient Genotypes
In the plant genome, TE activity is strictly controlled at different levels, including transcription (DNA methylation and histone modifications), post-transcription (post-transcriptional gene silencing), translation (ribosome stalling [29]), and post-translation barriers [29][30][31][32]. Among them, RNA-dependent DNA methylation pathways (PolIV-RDR2 and PolII-RDR6) tend to control most long and autonomous plant transposons [33]. Because DNA methylation plays a central role in TE silencing, mutations in genes involved in DNA methylation initiation and maintenance were used to trigger TE activity. Such mutants were obtained by mutagenesis, although a naturally occurring mutation in wild-type plants has also been reported [6]. The plants produced by crossing mutant lines with wild-types plants were used to produced so-called epiRILs (epigenetic recombinant inbred lines) [34][35]. Two types of mutants are mostly used for epiRIL production: met1 and ddm1 [36][37]. EpiRILs have extensively been used to study TEs and the epigenetic inheritance and trait control in model plants [6][10][34][35][37]. It was shown that, for example, Arabidopsis epiRILs possess multiple insertions of some TE families including AtCOPIA93, which possess EVD, one of the most active Arabidopsis LTR retrotransposons [38][39]. However, the application of epiRILs to boost TE transposition and phenotypic diversity in crops has not been demonstrated (Figure 1).
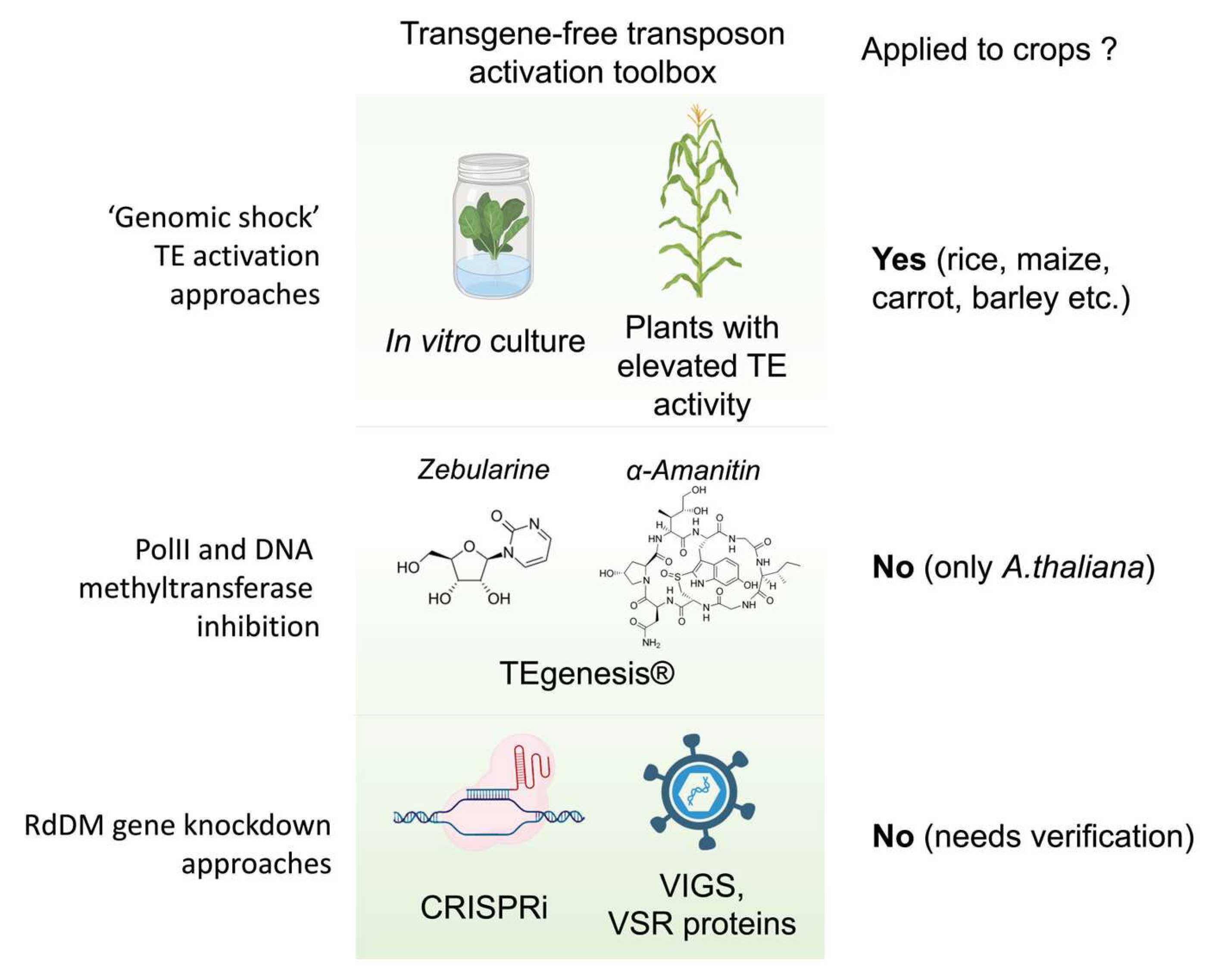
Figure 1. Approaches for TE-mediated biological mutagenesis: application of various stress factors (‘genomic shock’) such as crossing with plant lines possessing hyperactive TEs (e.g., Mutator lines of maize) or plant tissue culture (e.g., activation of Tos17 TE of rice); TEgenesis® (application of stress (e.g., heat stress) to the plants grown on medium containing zebularine (cytidine analog) and alpha-amanitin (a PolII inhibitor)); and new perspective tools (e.g., CRISPRi and VIGS) for transient knock-down of genes involved in RdDM pathway.
4. Utilization of Mutagenic Plant Lines with Reactivated Transposable Elements
Since the work of Barbara McClintock, it was discovered that some maize lines may possess increased mutagenic TE activity [40][41]. In her early works, McClintock observed ‘bizarre’ phenotypes after crossing maize lines carrying a chromosome with a ruptured end. Such phenotypes include leaves possessing twin sectors and cells with abnormal chlorophyll development [41]. Subsequently, the sequence of the corresponding transposons (Ac/Ds) was determined using the waxy allele carrying the insertion [23]. These studies showed, for the first time, that TEs can be naturally activated and that the insertions can be inherited, originating new alleles of genes. Sequencing of the Ac/Ds elements paved the way for the faster identification of the mutants and genes affected by insertions [42]. Another early example is Robertson’s Mutator maize lines, discovered by Robertson in 1978 [40] (Figure 1). Mutator-line mutagenic systems are based on the mutator (Mu) element, which is one of the most actively proliferating TEs in the maize genome [43]. Mu-like transposons have been identified in many other plant species, including the recently characterized VANDAL transposon family in Arabidopsis [11][44]. Mutator lines exhibit a 50–100 times higher frequency of mutations, as deduced by phenotypic analysis of kernels and seedlings. The “Mutator” locus generated these phenotypic abnormalities was dominant when the lines were crossed with maize lines lacking this locus. A few years later, the causative element for the “Mutator” locus was identified as a transposon and was called Mu1 [45]. Both Mutator and non-mutator maize lines have Mu-like elements, but only the former has Mu elements capable of transposition [46]. Interestingly, it was found that some maize lines carry a Muk locus (‘Mu killer’) that encodes a rearranged version of the autonomus MuDR element that is capable of producing small RNAs that can trigger the DNA methylation of active Mutator elements [47][48]. This natural system consisting of an active TE and its silencer can be useful for the activation of TEs and their heritable silencing when the required number of TEIs and mutagenesis efficiencies are achieved.
Mutator lines have been used to create collections of crop lines that carry TEIs [49][50]. These non-transgenic collections comprising tens of thousands of individuals have been created for maize and rice (TUSC (trait utility system for corn), UniformMu, and MTM) [51][52]. The first maize collection (TUSC) of plants with Mu insertions, developed by Pioneer Hi-Bred, provides useful information for accelerating genotype-based plant breeding and target gene discovery [53]. Transposon tagging has also been applied in rice via the utilization of plants of certain landraces possessing a hyperactive miniature Ping (mPing) transposon [52][54]. For example, the Aikoku and Gimbozu rice cultivars have >1000 mPing insertions compared to 51 copies in the Nipponbare cultivar [52]. Importantly, mPing TEs remained active and produced novel insertions after crossing Gimbozu plants with cultivars without active mPing, paving the way for the generation of mPing insertions in different rice cultivars.
Thus, several crop plant collections have been obtained via the utilization of lines with a natural hyperactivity of distinct TEs. These collections have been used in the plant breeding process and in functional genomic studies via the rapid isolation of genic mutations for reverse genetics.
References
- Ma, L.; Kong, F.; Sun, K.; Wang, T.; Guo, T. From Classical Radiation to Modern Radiation: Past, Present, and Future of Radiation Mutation Breeding. Front. Public Health 2021, 9, 768071.
- Greenwood, J.R.; Zhang, X.; Rathjen, J.P. Precision Genome Editing of Crops for Improved Disease Resistance. Curr. Biol. 2023, 33, R650–R657.
- Zhang, Y.; Massel, K.; Godwin, I.D.; Gao, C. Applications and Potential of Genome Editing in Crop Improvement. Genome Biol. 2018, 19, 210.
- Belfield, E.J.; Brown, C.; Ding, Z.J.; Chapman, L.; Luo, M.; Hinde, E.; Es, S.W.; van Johnson, S.; Ning, Y.; Zheng, S.J.; et al. Thermal Stress Accelerates Arabidopsis Thaliana Mutation Rate. Genome Res. 2021, 31, 40–50.
- Lu, Z.; Cui, J.; Wang, L.; Teng, N.; Zhang, S.; Lam, H.-M.; Zhu, Y.; Xiao, S.; Ke, W.; Lin, J.; et al. Genome-Wide DNA Mutations in Arabidopsis Plants after Multigenerational Exposure to High Temperatures. Genome Biol. 2021, 22, 160.
- Baduel, P.; Leduque, B.; Ignace, A.; Gy, I.; Gil, J.; Loudet, O.; Colot, V.; Quadrana, L. Genetic and Environmental Modulation of Transposition Shapes the Evolutionary Potential of Arabidopsis Thaliana. Genome Biol 2021, 22, 138.
- Hämälä, T.; Ning, W.; Kuittinen, H.; Aryamanesh, N.; Savolainen, O. Environmental Response in Gene Expression and DNA Methylation Reveals Factors Influencing the Adaptive Potential of Arabidopsis Lyrata. eLife 2022, 11, e83115.
- Quadrana, L.; Silveira, A.B.; Mayhew, G.F.; LeBlanc, C.; Martienssen, R.A.; Jeddeloh, J.A.; Colot, V. The Arabidopsis Thaliana Mobilome and Its Impact at the Species Level. eLife 2016, 5, e15716.
- Merkulov, P.; Gvaramiya, S.; Dudnikov, M.; Komakhin, R.; Omarov, M.; Kocheshkova, A.; Konstantinov, Z.; Soloviev, A.; Karlov, G.; Divashuk, M.; et al. Cas9-targeted Nanopore Sequencing Rapidly Elucidates the Transposition Preferences and DNA Methylation Profiles of Mobile Elements in Plants. J. Integr. Plant Biol. 2023, 65, 2242–2261.
- Quadrana, L.; Etcheverry, M.; Gilly, A.; Caillieux, E.; Madoui, M.-A.; Guy, J.; Silveira, A.B.; Engelen, S.; Baillet, V.; Wincker, P.; et al. Transposition Favors the Generation of Large Effect Mutations That May Facilitate Rapid Adaption. Nat. Commun. 2019, 10, 3421.
- Paszkowski, J. Controlled Activation of Retrotransposition for Plant Breeding. Curr. Opin. Biotechnol. 2015, 32, 200–206.
- Dubin, M.J.; Scheid, O.M.; Becker, C. Transposons: A Blessing Curse. Curr. Opin. Plant Biol. 2018, 42, 23–29.
- Domínguez, M.; Dugas, E.; Benchouaia, M.; Leduque, B.; Jiménez-Gómez, J.M.; Colot, V.; Quadrana, L. The Impact of Transposable Elements on Tomato Diversity. Nat. Commun. 2020, 11, 4058.
- Cai, X.; Lin, R.; Liang, J.; King, G.J.; Wu, J.; Wang, X. Transposable Element Insertion: A Hidden Major Source of Domesticated Phenotypic Variation in Brassica Rapa. Plant Biotechnol. J. 2022, 20, 1298–1310.
- Studer, A.; Zhao, Q.; Ross-Ibarra, J.; Doebley, J. Identification of a Functional Transposon Insertion in the Maize Domestication Gene Tb1. Nat. Genet. 2011, 43, 1160–1163.
- Wei, L.; Cao, X. The Effect of Transposable Elements on Phenotypic Variation: Insights from Plants to Humans. Sci. China Life Sci. 2016, 59, 24–37.
- Lisch, D. How Important Are Transposons for Plant Evolution? Nat. Rev. Genet. 2013, 14, 49–61.
- Latzel, V.; Puy, J.; Thieme, M.; Bucher, E.; Götzenberger, L.; Bello, F. Phenotypic Diversity Influenced by a Transposable Element Increases Productivity and Resistance to Competitors in Plant Populations. J. Ecol. 2023, 111, 2376–2387.
- Lieberman-Lazarovich, M.; Kaiserli, E.; Bucher, E.; Mladenov, V. Natural and Induced Epigenetic Variation for Crop Improvement. Curr. Opin. Plant Biol. 2022, 70, 102297.
- Thieme, M.; Lanciano, S.; Balzergue, S.; Daccord, N.; Mirouze, M.; Bucher, E. Inhibition of RNA Polymerase II Allows Controlled Mobilisation of Retrotransposons for Plant Breeding. Genome Biol. 2017, 18, 134.
- Thieme, M.; Brêchet, A.; Bourgeois, Y.; Keller, B.; Bucher, E.; Roulin, A.C. Experimentally Heat-induced Transposition Increases Drought Tolerance in Arabidopsis Thaliana. New Phytol. 2022, 236, 182–194.
- Roquis, D.; Robertson, M.; Yu, L.; Thieme, M.; Julkowska, M.; Bucher, E. Genomic Impact of Stress-Induced Transposable Element Mobility in Arabidopsis. Nucleic Acids Res. 2021, 49, gkab828.
- Fedoroff, N.; Wessler, S.; Shure, M. Isolation of the Transposable Maize Controlling Elements Ac and Ds. Cell 1983, 35, 235–242.
- Baker, B.; Schell, J.; Lörz, H.; Fedoroff, N. Transposition of the Maize Controlling Element “Activator” in Tobacco. Proc. Natl. Acad. Sci. USA 1986, 83, 4844–4848.
- Yamazaki, M.; Tsugawa, H.; Miyao, A.; Yano, M.; Wu, J.; Yamamoto, S.; Matsumoto, T.; Sasaki, T.; Hirochika, H. The Rice Retrotransposon Tos17 Prefers Low-Copy-Number Sequences as Integration Targets. Mol. Genet. Genom. 2001, 265, 336–344.
- Okamoto, H.; Hirochika, H. Efficient Insertion Mutagenesis of Arabidopsis by Tissue Culture-induced Activation of the Tobacco Retrotransposon Tto1. Plant J. 2000, 23, 291–304.
- Courtial, B.; Feuerbach, F.; Eberhard, S.; Rohmer, L.; Chiapello, H.; Camilleri, C.; Lucas, H. Tnt1 Transposition Events Are Induced by in Vitro Transformation of Arabidopsis Thaliana, and Transposed Copies Integrate into Genes. Mol. Genet. Genom. 2001, 265, 32–42.
- D’Erfurth, I.; Cosson, V.; Eschstruth, A.; Lucas, H.; Kondorosi, A.; Ratet, P. Efficient Transposition of the Tnt1 Tobacco Retrotransposon in the Model Legume Medicago Truncatula. Plant J. 2003, 34, 95–106.
- Kim, E.Y.; Wang, L.; Lei, Z.; Li, H.; Fan, W.; Cho, J. Ribosome Stalling and SGS3 Phase Separation Prime the Epigenetic Silencing of Transposons. Nat. Plants 2021, 7, 303–309.
- Nicolau, M.; Picault, N.; Moissiard, G. The Evolutionary Volte-Face of Transposable Elements: From Harmful Jumping Genes to Major Drivers of Genetic Innovation. Cells 2021, 10, 2952.
- Liu, B.; Zhao, M. How Transposable Elements Are Recognized and Epigenetically Silenced in Plants? Curr. Opin. Plant Biol. 2023, 75, 102428.
- Liu, P.; Cuerda-Gil, D.; Shahid, S.; Slotkin, R.K. The Epigenetic Control of the Transposable Element Life Cycle in Plant Genomes and Beyond. Annu. Rev. Genet. 2022, 56, 63–87.
- Panda, K.; Ji, L.; Neumann, D.A.; Daron, J.; Schmitz, R.J.; Slotkin, R.K. Full-Length Autonomous Transposable Elements Are Preferentially Targeted by Expression-Dependent Forms of RNA-Directed DNA Methylation. Genome Biol. 2016, 17, 170.
- Catoni, M.; Cortijo, S. Chapter Four EpiRILs Lessons From Arabidopsis. Adv. Bot. Res. 2018, 88, 87–116.
- Kooke, R.; Johannes, F.; Wardenaar, R.; Becker, F.; Etcheverry, M.; Colot, V.; Vreugdenhil, D.; Keurentjes, J.J.B. Epigenetic Basis of Morphological Variation and Phenotypic Plasticity in Arabidopsis Thaliana. Plant Cell 2015, 27, 337–348.
- Reinders, J.; Wulff, B.B.H.; Mirouze, M.; Marí-Ordóñez, A.; Dapp, M.; Rozhon, W.; Bucher, E.; Theiler, G.; Paszkowski, J. Compromised Stability of DNA Methylation and Transposon Immobilization in Mosaic Arabidopsis Epigenomes. Genes Dev. 2009, 23, 939–950.
- Johannes, F.; Porcher, E.; Teixeira, F.K.; Saliba-Colombani, V.; Simon, M.; Agier, N.; Bulski, A.; Albuisson, J.; Heredia, F.; Audigier, P.; et al. Assessing the Impact of Transgenerational Epigenetic Variation on Complex Traits. PLoS Genet. 2009, 5, e1000530.
- Tsukahara, S.; Kobayashi, A.; Kawabe, A.; Mathieu, O.; Miura, A.; Kakutani, T. Bursts of Retrotransposition Reproduced in Arabidopsis. Nature 2009, 461, 423–426.
- Mirouze, M.; Reinders, J.; Bucher, E.; Nishimura, T.; Schneeberger, K.; Ossowski, S.; Cao, J.; Weigel, D.; Paszkowski, J.; Mathieu, O. Selective Epigenetic Control of Retrotransposition in Arabidopsis. Nature 2009, 461, 427–430.
- Robertson, D.S. Characterization of a Mutator System in Maize. Mutat. Res. Fundam. Mol. Mech. Mutagen. 1978, 51, 21–28.
- McClintock, B. The Significance of Responses of the Genome to Challenge. Science 1984, 226, 792–801.
- Richardson, A.E.; Hake, S. The Power of Classic Maize Mutants: Driving Forward Our Fundamental Understanding of Plants. Plant Cell 2021, 34, 2505–2517.
- Lisch, D. Mutator Transposons. Trends Plant Sci. 2002, 7, 498–504.
- Sasaki, T.; Kato, K.; Hosaka, A.; Fu, Y.; Toyoda, A.; Fujiyama, A.; Tarutani, Y.; Kakutani, T. Arms Race between Anti-silencing and RdDM in Noncoding Regions of Transposable Elements. EMBO Rep. 2023, 24, e56678.
- Barker, R.F.; Thompson, D.V.; Talbot, D.R.; Swanson, J.; Bennetzen, J.L. Nucleotide Sequence of the Maize Transposable Element Mul. Nucleic Acids Res. 1984, 12, 6924.
- Alleman, M.; Freeling, M. The Mu Transposable Elements Of Maize: Evidence For Transposition And Copy Number Regulation During Development. Genetics 1986, 112, 107–119.
- Slotkin, R.K.; Freeling, M.; Lisch, D. Mu Killer Causes the Heritable Inactivation of the Mutator Family of Transposable Elements in Zea Mays. Genetics 2003, 165, 781–797.
- Burgess, D.; Li, H.; Zhao, M.; Kim, S.Y.; Lisch, D. Silencing of Mutator Elements in Maize Involves Distinct Populations of Small RNAs and Distinct Patterns of DNA Methylation. Genetics 2020, 215, 379–391.
- Kolkman, J.M.; Conrad, L.J.; Farmer, P.R.; Hardeman, K.; Ahern, K.R.; Lewis, P.E.; Sawers, R.J.H.; Lebejko, S.; Chomet, P.; Brutnell, T.P. Distribution of Activator (Ac) Throughout the Maize Genome for Use in Regional Mutagenesis. Genetics 2005, 169, 981–995.
- Bai, L.; Singh, M.; Pitt, L.; Sweeney, M.; Brutnell, T.P. Generating Novel Allelic Variation Through Activator Insertional Mutagenesis in Maize. Genetics 2007, 175, 981–992.
- Bensen, R.J.; Johal, G.S.; Crane, V.C.; Tossberg, J.T.; Schnable, P.S.; Meeley, R.B.; Briggs, S.P. Cloning and Characterization of the Maize An1 Gene. Plant Cell 1995, 7, 75–84.
- Naito, K.; Cho, E.; Yang, G.; Campbell, M.A.; Yano, K.; Okumoto, Y.; Tanisaka, T.; Wessler, S.R. Dramatic Amplification of a Rice Transposable Element during Recent Domestication. Proc. Natl. Acad. Sci. USA 2006, 103, 17620–17625.
- Simmons, C.R.; Lafitte, H.R.; Reimann, K.S.; Brugière, N.; Roesler, K.; Albertsen, M.C.; Greene, T.W.; Habben, J.E. Successes and Insights of an Industry Biotech Program to Enhance Maize Agronomic Traits. Plant Sci. 2021, 307, 110899.
- Yasuda, K.; Ito, M.; Sugita, T.; Tsukiyama, T.; Saito, H.; Naito, K.; Teraishi, M.; Tanisaka, T.; Okumoto, Y. Utilization of Transposable Element MPing as a Novel Genetic Tool for Modification of the Stress Response in Rice. Mol. Breed. 2013, 32, 505–516.
More
Information
Subjects:
Plant Sciences
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
618
Revisions:
3 times
(View History)
Update Date:
06 Dec 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No