Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Ilya Kirov and Version 3 by Jessie Wu.
Genetic diversity is a key factor for plant breeding. The birth of novel genic and genomic variants is also crucial for plant adaptation in nature. Therefore, the genomes of almost all living organisms possess natural mutagenic mechanisms. Transposable elements (TEs) are a major mutagenic force driving genetic diversity in wild plants and modern crops. The relatively rare TE transposition activity during the thousand-year crop domestication process has led to the phenotypic diversity of many cultivated species. The utilization of TE mutagenesis by artificial and transient acceleration of their activity in a controlled mode is an attractive foundation for a novel type of mutagenesis called TE-mediated biological mutagenesis.
- transposons
- plants
- mutagenesis
- RNA-dependent DNA methylation
- gene silencing
1. Introduction
Plant breeding relies on the genetic diversity available to breeders, which is used to generate new combinations of genes and traits. Therefore, methods and approaches for generating genetic diversity in crops are the cornerstones of the plant breeding process. Implication of wild relatives and different genotypes and varieties in crosses followed by the selection of the most desirable genotypes from the progeny is a common way to create genetic diversity. Another approach involves chemical (e.g., EMS treatment) and radiation mutagenesis [1]. Mutagenesis has led to the development of new crop varieties and genetic resources [1]. Mutated genetic populations have been created for many plant species, leading to tremendous progress in functional genomics. However, radiation mutagenesis has several disadvantages, including a low frequency of valuable mutations and challenging identification of the causative mutations. However, novel techniques for precise gene targeting (e.g., genome editing) have been developed and adapted for many crop species [2][3][2,3]. Therefore, the application of classical mutagenesis in plant breeding has significantly decreased in recent decades [1].
Mutagenesis also occurs in nature but at a significantly lower rate than that achieved by artificial mutagenesis. It is obvious that in contrast to animals, plants cannot escape stress factors and cannot conquer new ecological niches by long-distance physical movement. Therefore, the main strategy for plants to adapt to new environmental conditions and stress is to explore their natural genetic diversity. It is now clear that environmental stress may increase the rate of single-nucleotide mutations [4] as well as structural rearrangements, including transposable element (TE) movements [5]. In the last few decades, a list of studies have concluded that stress may stimulate transposable elements (TEs) to transpose in the genome and to broaden genetic diversity [6][7][8][6,7,8]. Importantly, novel TE insertions (TEIs) are not randomly generated over the genome and may occur frequently in stress-responsive genes, providing connections between stress action and the occurrence of novel alleles of stress-responsive genes [9][10][11][9,10,11]. This indicates that TE transposition is an evolutionary mechanism that serves as a natural biological mutagen and an important source of genetic diversity for plant adaptation.
Interestingly, TE transposition is also a major source of crop diversity and has been widely used by plant breeders for hundreds of years [12][13][12,13]. The gene candidate approach revealed a number of cases in which TE transposition in or near the genes caused the origin of novel phenotypic variants in different species [6][10][13][6,10,13]. Modern genomics has provided tools to perform genome-wide analyses of the association between trait variation and TEIs. Indeed, recent genome-wide association studies (GWASs) exploiting small variants (for example, SNPs and InDels) and TEI polymorphisms (TIPs) have revealed the great impact of TIPs on crop domestication and breeding [13][14][13,14]. For example, Hopscotch TE insertion into the teosinte branched1 (tb1) gene is associated with increased apical dominance in modern maize [15]. Also, TE insertions were shown to play a key role in tomato domestication, causing changes in tomato leaf shape, scene, and fruit color [13]. Recently, the implication of TE insertions in the diversity of modern Brassica rapa morphotypes has been demonstrated [14]. Therefore, TEs can be considered one of the most effective biological mutagens that can be exploited by plant breeders. The most recent understanding of TE biology and TE suppression mechanisms (e.g., RNA-directed DNA methylation pathways and various small RNAs), combined with the most recent molecular tools (e.g., CRISPR-based technology and virus-induced gene silencing), could allow for the use of TE mutational potential in the control mode.
TE insertions may result in a broad spectrum of consequences at the genome, epigenetic, transcriptome, and phenotype levels [12][13][14][15][16][17][18][12,13,14,15,16,17,18]. Based on this, the idea that the controlled activation of native TEs in a plant genome can bring new TE-mediated mutagenesis technology to accelerate plant breeding has been proposed and discussed recently [11][12][19][11,12,19]. In recent years, the validity of this concept has been established through research on the model organism Arabidopsis thaliana [18][19][20][21][22][18,19,20,21,22], but studies on crops are still lagging behind.
2. Heterologous Expression of Transposable Elements via Plant Transformation
In 1983, the first active plant transposon, Ac, was cloned and sequenced from maize [23]. This began the era of TE application for the mutagenesis of plants via genetic transformation. In initial reports, the active TEs of one species were transformed into the genomes of other species to overcome host silencing mechanisms [24]. Using this approach, the transformed TEs were kept active in the foreign genome until they were recognized by the host silencing system. For example, Tos17 (rice), Tnt1, and TtoI (tobacco) retrotransposons were involved in gene tagging experiments in rice, Arabidopsis thaliana, Medicago truncatula, and other plant species [24][25][26][27][28][24,25,26,27,28]. It is important to note that these TEs are cell-culture-inducible, and therefore their activity can be tightly controlled or reactivated by a new round of in vitro regeneration [28]. Although heterologous expression of TEs plays an important role in understanding the TE transposition process and its silencing, the application of this approach for plant breeding is challenging in many countries because of the GMO legislation. Therefore, alternative approaches are required to establish GMO-free, breeder-friendly methods for TE-mediated mutagenesis.3. Implication of DNA-Methylation-Deficient Genotypes
In the plant genome, TE activity is strictly controlled at different levels, including transcription (DNA methylation and histone modifications), post-transcription (post-transcriptional gene silencing), translation (ribosome stalling [29]), and post-translation barriers [29][30][31][32][29,30,31,32]. Among them, RNA-dependent DNA methylation pathways (PolIV-RDR2 and PolII-RDR6) tend to control most long and autonomous plant transposons [33]. Because DNA methylation plays a central role in TE silencing, mutations in genes involved in DNA methylation initiation and maintenance were used to trigger TE activity. Such mutants were obtained by mutagenesis, although a naturally occurring mutation in wild-type plants has also been reported [6]. The plants produced by crossing mutant lines with wild-types plants were used to produced so-called epiRILs (epigenetic recombinant inbred lines) [34][35][34,35]. Two types of mutants are mostly used for epiRIL production: met1 and ddm1 [36][37][36,37]. EpiRILs have extensively been used to study TEs and the epigenetic inheritance and trait control in model plants [6][10][34][35][37][6,10,34,35,37]. It was shown that, for example, Arabidopsis epiRILs possess multiple insertions of some TE families including AtCOPIA93, which possess EVD, one of the most active Arabidopsis LTR retrotransposons [38][39][38,39]. However, the application of epiRILs to boost TE transposition and phenotypic diversity in crops has not been demonstrated (Figure 1).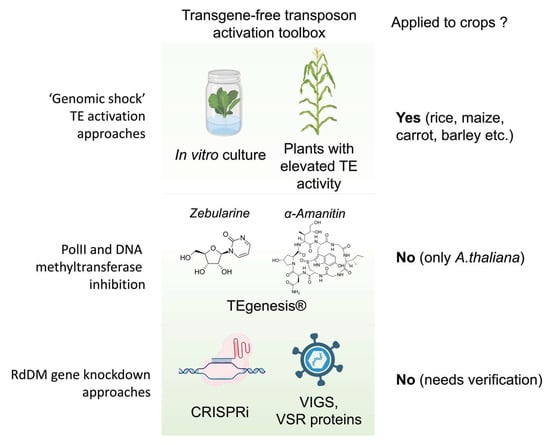
Figure 1. Approaches for TE-mediated biological mutagenesis: application of various stress factors (‘genomic shock’) such as crossing with plant lines possessing hyperactive TEs (e.g., Mutator lines of maize) or plant tissue culture (e.g., activation of Tos17 TE of rice); TEgenesis® (application of stress (e.g., heat stress) to the plants grown on medium containing zebularine (cytidine analog) and alpha-amanitin (a PolII inhibitor)); and new perspective tools (e.g., CRISPRi and VIGS) for transient knock-down of genes involved in RdDM pathway.