 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Bojana Kožik | -- | 2683 | 2023-11-23 14:25:17 | | | |
| 2 | Lindsay Dong | + 2 word(s) | 2685 | 2023-11-24 07:36:42 | | |
Video Upload Options
Hepatocellular carcinoma (HCC) is one of the most frequent cancers in humans, characterised by a high resistance to conventional chemotherapy, late diagnosis, and a high mortality rate. It is necessary to elucidate the molecular mechanisms involved in hepatocarcinogenesis to improve diagnosis and treatment outcomes. The Runt-related (RUNX) family of transcription factors (RUNX1, RUNX2, and RUNX3) participates in cardinal biological processes and plays paramount roles in the pathogenesis of numerous human malignancies. Their role is often controversial as they can act as oncogenes or tumour suppressors and depends on cellular context. Evidence shows that deregulated RUNX genes may be involved in hepatocarcinogenesis from the earliest to the latest stages.
1. Introduction
1.1. RUNX Genes’ and Proteins’ Structure
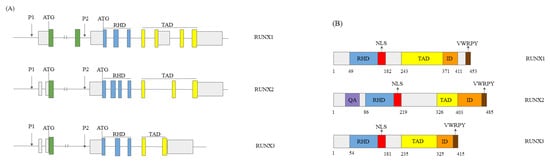
1.2. The Role of the RUNX Genes in Normal Development
1.3. The Role of RUNX Genes in Cancer
1.4. The Mechanisms of Action of RUNX Proteins
2. RUNX1 in HCC
2.1. RUNX1 Role in HCC
2.2. RUNX1 and miRNAs
3. RUNX2 in HCC
3.1. General Role of RUNX2 in HCC
3.2. RUNX2 Tumour Invasion Activity in HCC
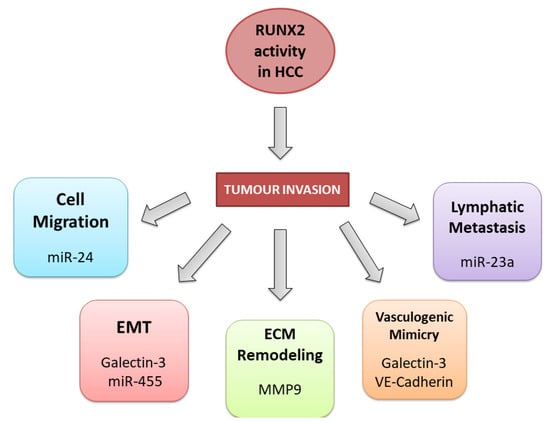
3.3. RUNX2 and Non-Coding RNAs in HCC
4. RUNX3 in HCC
4.1. General Role of RUNX3 in HCC
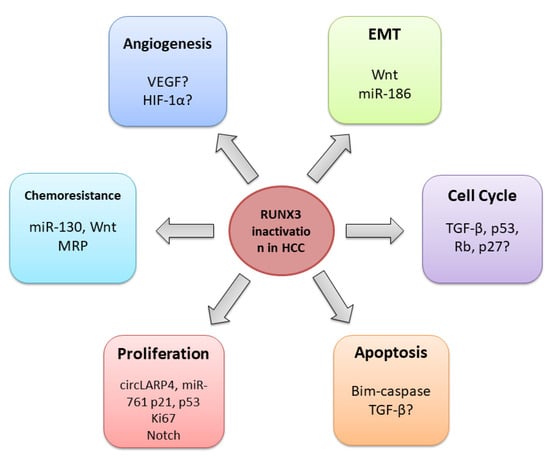
4.2. RUNX3 Regulates Cell Cycle, Proliferation, and Apoptosis
4.3. RUNX3 in the Angiogenesis Regulation
4.4. RUNX3 and Epithelial-Mesenchymal Transition
4.5. RUNX3 and Chemoresistance
5. Conclusions
Despite considerable advances in cancer diagnosis and treatment, HCC remains one of the most common and hard-to-treat human cancers. Revealing the essential molecular processes underlying hepatocarcinogenesis is crucial for establishing reliable diagnostic, prognostic, and therapeutic markers. RUNX genes are often deregulated in HCC, exerting complex and conflicting functions. The role of RUNX1 is still contradictory, as there are reports of its tumour-suppressive but also oncogenic role in HCC. According to current knowledge, RUNX2 acts as an oncogene and is related to the more aggressive forms of the disease, whereas RUNX3 exerts a tumour-suppressive role and could be used as a biomarker for early HCC detection. All three genes could serve as therapeutic targets. However, a deeper understanding of the relationship between different RUNX family members and the signalling pathways they are involved in, considering the cell-specific microenvironment, is necessary for effective HCC therapeutic strategy development.
Increasing evidence suggests that RUNX genes act as epigenetic modulators that interact with other chromatin landscape regulators to activate or repress the transcription of target genes [89]. Since normal epigenetic patterns are altered in all types of human cancers, it would be of great interest to investigate interactions between RUNX proteins and other epigenetic regulators, especially in HCC. This could potentially provide an avenue for epigenetic therapy.
References
- Asafo-Agyei, K.O.; Samant, H. Hepatocellular Carcinoma. In StatPearls; StatPearls Publishing: Treasure Islan, FL, USA, 2023.
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular Carcinoma. Nat. Rev. Dis. Prim. 2021, 7, 6.
- Balogh, J.; Victor, D.; Asham, E.H.; Burroughs, S.G.; Boktour, M.; Saharia, A.; Li, X.; Ghobrial, M.; Monsour, H. Hepatocellular Carcinoma: A Review. J. Hepatocell. Carcinoma 2016, 3, 41–53.
- Sim, H.-W.; Knox, J. Hepatocellular Carcinoma in the Era of Immunotherapy. Curr. Probl. Cancer 2018, 42, 40–48.
- Suresh, D.; Srinivas, A.N.; Prashant, A.; Harikumar, K.B.; Kumar, D.P. Therapeutic Options in Hepatocellular Carcinoma: A Comprehensive Review. Clin. Exp. Med. 2023.
- Chuang, L.S.H.; Ito, K.; Ito, Y. RUNX Family: Regulation and Diversification of Roles through Interacting Proteins. Int. J. Cancer 2013, 132, 1260–1271.
- Otálora-Otálora, B.A.; Henríquez, B.; López-Kleine, L.; Rojas, A. RUNX Family: Oncogenes or Tumor Suppressors (Review). Oncol. Rep. 2019, 42, 3–19.
- Ito, Y.; Bae, S.-C.; Chuang, L.S.H. The RUNX Family: Developmental Regulators in Cancer. Nat. Rev. Cancer 2015, 15, 81–95.
- Levanon, D.; Groner, Y. Structure and Regulated Expression of Mammalian RUNX Genes. Oncogene 2004, 23, 4211–4219.
- Yi, H.; He, Y.; Zhu, Q.; Fang, L. RUNX Proteins as Epigenetic Modulators in Cancer. Cells 2022, 11, 3687.
- Mevel, R.; Draper, J.E.; Lie-a-Ling, M.; Kouskoff, V.; Lacaud, G. RUNX Transcription Factors: Orchestrators of Development. Development 2019, 146, dev148296.
- Darnell, J.E. Transcription Factors as Targets for Cancer Therapy. Nat. Rev. Cancer 2002, 2, 740–749.
- Ito, Y. Oncogenic Potential of the RUNX Gene Family: ‘Overview’. Oncogene 2004, 23, 4198–4208.
- Lee, Y.M. RUNX Family in Hypoxic Microenvironment and Angiogenesis in Cancers. Cells 2022, 11, 3098.
- Sood, R.; Kamikubo, Y.; Liu, P. Role of RUNX1 in Hematological Malignancies. Blood 2017, 129, 2070–2082.
- Yamagata, T.; Maki, K.; Mitani, K. Runx1/AML1 in Normal and Abnormal Hematopoiesis. Int. J. Hematol. 2005, 82, 1–8.
- Dalle Carbonare, L.; Innamorati, G.; Valenti, M.T. Transcription Factor Runx2 and Its Application to Bone Tissue Engineering. Stem Cell Rev. Rep. 2012, 8, 891–897.
- Lee, J.-M.; Lee, D.-J.; Bae, S.-C.; Jung, H.-S. Abnormal Liver Differentiation and Excessive Angiogenesis in Mice Lacking Runx3. Histochem. Cell Biol. 2013, 139, 751–758.
- Pan, S.; Sun, S.; Liu, B.; Hou, Y. Pan-Cancer Landscape of the RUNX Protein Family Reveals Their Potential as Carcinogenic Biomarkers and the Mechanisms Underlying Their Action. J. Transl. Intern. Med. 2022, 10, 156–174.
- Papaemmanuil, E.; Gerstung, M.; Bullinger, L.; Gaidzik, V.I.; Paschka, P.; Roberts, N.D.; Potter, N.E.; Heuser, M.; Thol, F.; Bolli, N.; et al. Genomic Classification and Prognosis in Acute Myeloid Leukemia. N. Engl. J. Med. 2016, 374, 2209–2221.
- Keita, M.; Bachvarova, M.; Morin, C.; Plante, M.; Gregoire, J.; Renaud, M.-C.; Sebastianelli, A.; Trinh, X.B.; Bachvarov, D. The RUNX1 Transcription Factor Is Expressed in Serous Epithelial Ovarian Carcinoma and Contributes to Cell Proliferation, Migration and Invasion. Cell Cycle 2013, 12, 972–986.
- Liu, C.; Xu, D.; Xue, B.; Liu, B.; Li, J.; Huang, J. Upregulation of RUNX1 Suppresses Proliferation and Migration through Repressing VEGFA Expression in Hepatocellular Carcinoma. Pathol. Oncol. Res. 2020, 26, 1301–1311.
- Li, Q.; Lai, Q.; He, C.; Fang, Y.; Yan, Q.; Zhang, Y.; Wang, X.; Gu, C.; Wang, Y.; Ye, L.; et al. RUNX1 Promotes Tumour Metastasis by Activating the Wnt/β-Catenin Signalling Pathway and EMT in Colorectal Cancer. J. Exp. Clin. Cancer Res. 2019, 38, 334.
- Lu, C.; Yang, Z.; Yu, D.; Lin, J.; Cai, W. RUNX1 Regulates TGF-β Induced Migration and EMT in Colorectal Cancer. Pathol. Res. Pract. 2020, 216, 153142.
- Sangpairoj, K.; Vivithanaporn, P.; Apisawetakan, S.; Chongthammakun, S.; Sobhon, P.; Chaithirayanon, K. RUNX1 Regulates Migration, Invasion, and Angiogenesis via P38 MAPK Pathway in Human Glioblastoma. Cell. Mol. Neurobiol. 2017, 37, 1243–1255.
- Fritz, A.J.; Hong, D.; Boyd, J.; Kost, J.; Finstaad, K.H.; Fitzgerald, M.P.; Hanna, S.; Abuarqoub, A.H.; Malik, M.; Bushweller, J.; et al. RUNX1 and RUNX2 Transcription Factors Function in Opposing Roles to Regulate Breast Cancer Stem Cells. J. Cell. Physiol. 2020, 235, 7261–7272.
- Lin, T.-C. RUNX2 and Cancer. Int. J. Mol. Sci. 2023, 24, 7001.
- Lee, Y.-S.; Lee, J.-W.; Jang, J.-W.; Chi, X.-Z.; Kim, J.-H.; Li, Y.-H.; Kim, M.-K.; Kim, D.-M.; Choi, B.-S.; Kim, E.-G.; et al. Runx3 Inactivation Is a Crucial Early Event in the Development of Lung Adenocarcinoma. Cancer Cell 2013, 24, 603–616.
- Li, Q.-L.; Ito, K.; Sakakura, C.; Fukamachi, H.; Inoue, K.; Chi, X.-Z.; Lee, K.-Y.; Nomura, S.; Lee, C.-W.; Han, S.-B.; et al. Causal Relationship between the Loss of RUNX3 Expression and Gastric Cancer. Cell 2002, 109, 113–124.
- Goel, A.; Arnold, C.N.; Tassone, P.; Chang, D.K.; Niedzwiecki, D.; Dowell, J.M.; Wasserman, L.; Compton, C.; Mayer, R.J.; Bertagnolli, M.M.; et al. Epigenetic Inactivation OfRUNX3 in Microsatellite Unstable Sporadic Colon Cancers. Int. J. Cancer 2004, 112, 754–759.
- Kang, G.H.; Lee, S.; Lee, H.J.; Hwang, K.S. Aberrant CpG Island Hypermethylation of Multiple Genes in Prostate Cancer and Prostatic Intraepithelial Neoplasia: CpG Island Methylation in Prostate Cancer and PIN. J. Pathol. 2004, 202, 233–240.
- Lau, Q.C.; Raja, E.; Salto-Tellez, M.; Liu, Q.; Ito, K.; Inoue, M.; Putti, T.C.; Loh, M.; Ko, T.K.; Huang, C.; et al. RUNX3 Is Frequently Inactivated by Dual Mechanisms of Protein Mislocalization and Promoter Hypermethylation in Breast Cancer. Cancer Res. 2006, 66, 6512–6520.
- Tsunematsu, T.; Kudo, Y.; Iizuka, S.; Ogawa, I.; Fujita, T.; Kurihara, H.; Abiko, Y.; Takata, T. RUNX3 Has an Oncogenic Role in Head and Neck Cancer. PLoS ONE 2009, 4, e5892.
- Zhang, Z.; Chen, G.; Cheng, Y.; Martinka, M.; Li, G. Prognostic Significance of RUNX3 Expression in Human Melanoma: RUNX3 in Melanoma Prognosis. Cancer 2011, 117, 2719–2727.
- Nevadunsky, N.S.; Barbieri, J.S.; Kwong, J.; Merritt, M.A.; Welch, W.R.; Berkowitz, R.S.; Mok, S.C. RUNX3 Protein Is Overexpressed in Human Epithelial Ovarian Cancer. Gynecol. Oncol. 2009, 112, 325–330.
- Kim, W.-J.; Kim, E.-J.; Jeong, P.; Quan, C.; Kim, J.; Li, Q.-L.; Yang, J.-O.; Ito, Y.; Bae, S.-C. RUNX3 Inactivation by Point Mutations and Aberrant DNA Methylation in Bladder Tumors. Cancer Res. 2005, 65, 9347–9354.
- Lund, A.H.; Van Lohuizen, M. RUNX: A Trilogy of Cancer Genes. Cancer Cell 2002, 1, 213–215.
- Nakanishi, Y.; Shiraha, H.; Nishina, S.; Tanaka, S.; Matsubara, M.; Horiguchi, S.; Iwamuro, M.; Takaoka, N.; Uemura, M.; Kuwaki, K.; et al. Loss of Runt-Related Transcription Factor 3 Expression Leads Hepatocellular Carcinoma Cells to Escape Apoptosis. BMC Cancer 2011, 11, 3.
- Mori, T.; Nomoto, S.; Koshikawa, K.; Fujii, T.; Sakai, M.; Nishikawa, Y.; Inoue, S.; Takeda, S.; Kaneko, T.; Nakao, A. Decreased Expression and Frequent Allelic Inactivation of the RUNX3 Gene at 1p36 in Human Hepatocellular Carcinoma. Liver Int. 2005, 25, 380–388.
- Steinhart, Z.; Angers, S. Wnt Signaling in Development and Tissue Homeostasis. Development 2018, 145, dev146589.
- Ito, K.; Lim, A.C.-B.; Salto-Tellez, M.; Motoda, L.; Osato, M.; Chuang, L.S.H.; Lee, C.W.L.; Voon, D.C.-C.; Koo, J.K.W.; Wang, H.; et al. RUNX3 Attenuates β-Catenin/T Cell Factors in Intestinal Tumorigenesis. Cancer Cell 2008, 14, 226–237.
- Lee, Y.M. Control of RUNX3 by Histone Methyltransferases. J. Cell. Biochem. 2011, 112, 394–400.
- Lee, S.H.; Kim, J.; Kim, W.-H.; Lee, Y.M. Hypoxic Silencing of Tumor Suppressor RUNX3 by Histone Modification in Gastric Cancer Cells. Oncogene 2009, 28, 184–194.
- Zaret, K.S.; Carroll, J.S. Pioneer Transcription Factors: Establishing Competence for Gene Expression. Genes Dev. 2011, 25, 2227–2241.
- Lin, T.-C. RUNX1 and Cancer. Biochim. Biophys. Acta (BBA) Rev. Cancer 2022, 1877, 188715.
- TIMER2.0. Available online: http://timer.cistrome.org/ (accessed on 3 April 2023).
- GENT2. Available online: http://gent2.appex.kr/gent2/ (accessed on 3 April 2023).
- Zhu, J.; Sanborn, J.Z.; Benz, S.; Szeto, C.; Hsu, F.; Kuhn, R.M.; Karolchik, D.; Archie, J.; Lenburg, M.E.; Esserman, L.J.; et al. The UCSC Cancer Genomics Browser. Nat. Methods 2009, 6, 239–240.
- UCSC Xena. Available online: http://xena.ucsc.edu/welcome-to-ucsc-xena/ (accessed on 3 April 2023).
- Miyagawa, K.; Sakakura, C.; Nakashima, S.; Yoshikawa, T.; Kin, S.; Nakase, Y.; Ito, K.; Yamagishi, H.; Ida, H.; Yazumi, S.; et al. Down-Regulation of RUNX1, RUNX3 and CBFbeta in Hepatocellular Carcinomas in an Early Stage of Hepatocarcinogenesis. Anticancer Res. 2006, 26, 3633–3643.
- Takakura, N.; Watanabe, T.; Suenobu, S.; Yamada, Y.; Noda, T.; Ito, Y.; Satake, M.; Suda, T. A Role for Hematopoietic Stem Cells in Promoting Angiogenesis. Cell 2000, 102, 199–209.
- Iwatsuki, K.; Tanaka, K.; Kaneko, T.; Kazama, R.; Okamoto, S.; Nakayama, Y.; Ito, Y.; Satake, M.; Takahashi, S.-I.; Miyajima, A.; et al. Runx1 Promotes Angiogenesis by Downregulation of Insulin-like Growth Factor-Binding Protein-3. Oncogene 2005, 24, 1129–1137.
- Ter Elst, A.; Ma, B.; Scherpen, F.J.G.; De Jonge, H.J.M.; Douwes, J.; Wierenga, A.T.J.; Schuringa, J.J.; Kamps, W.A.; De Bont, E.S.J.M. Repression of Vascular Endothelial Growth Factor Expression by the Runt-Related Transcription Factor 1 in Acute Myeloid Leukemia. Cancer Res. 2011, 71, 2761–2771.
- Tuo, Z.; Zhang, Y.; Wang, X.; Dai, S.; Liu, K.; Xia, D.; Wang, J.; Bi, L. RUNX1 Is a Promising Prognostic Biomarker and Related to Immune Infiltrates of Cancer-Associated Fibroblasts in Human Cancers. BMC Cancer 2022, 22, 523.
- National Library of Medicine. RUNX1-IT1 RUNX1 Intronic Transcript 1 . Available online: https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=80215 (accessed on 18 March 2023).
- Yan, P.-H.; Wang, L.; Chen, H.; Yu, F.-Q.; Guo, L.; Liu, Y.; Zhang, W.-J.; Bai, Y.-L. LncRNA RUNX1-IT1 Inhibits Proliferation and Promotes Apoptosis of Hepatocellular Carcinoma by Regulating MAPK Pathways. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8287–8294.
- Sun, L.; Wang, L.; Chen, T.; Shi, Y.; Yao, B.; Liu, Z.; Wang, Y.; Li, Q.; Liu, R.; Niu, Y.; et al. LncRNA RUNX1-IT1 Which Is Downregulated by Hypoxia-Driven Histone Deacetylase 3 Represses Proliferation and Cancer Stem-like Properties in Hepatocellular Carcinoma Cells. Cell Death Dis. 2020, 11, 95.
- Vivacqua, A.; De Marco, P.; Santolla, M.F.; Cirillo, F.; Pellegrino, M.; Panno, M.L.; Abonante, S.; Maggiolini, M. Estrogenic Gper Signaling Regulates Mir144 Expression in Cancer Cells and Cancer-Associated Fibroblasts (Cafs). Oncotarget 2015, 6, 16573–16587.
- Jing, G.; Zheng, X.; Ji, X. LncRNA HAND2-AS1 Overexpression Inhibits Cancer Cell Proliferation in Hepatocellular Carcinoma by Downregulating RUNX2 Expression. J. Clin. Lab. Anal. 2021, 35, e23717.
- Wang, Q.; Yu, W.; Huang, T.; Zhu, Y.; Huang, C. RUNX2 Promotes Hepatocellular Carcinoma Cell Migration and Invasion by Upregulating MMP9 Expression. Oncol. Rep. 2016, 36, 2777–2784.
- Cao, Z.; Sun, B.; Zhao, X.; Zhang, Y.; Gu, Q.; Liang, X.; Dong, X.; Zhao, N. Correction: Cao, Z.; et al. The Expression and Functional Significance of Runx2 in Hepatocellular Carcinoma: Its Role in Vasculogenic Mimicry and Epithelial—Mesenchymal Transition. Int. J. Mol. Sci. 2020, 18, 500.
- Emma, M.R.; Iovanna, J.L.; Bachvarov, D.; Puleio, R.; Loria, G.R.; Augello, G.; Candido, S.; Libra, M.; Gulino, A.; Cancila, V.; et al. NUPR1, a New Target in Liver Cancer: Implication in Controlling Cell Growth, Migration, Invasion and Sorafenib Resistance. Cell Death Dis. 2016, 7, e2269.
- Wang, X.; Li, L.; Wu, Y.; Zhang, R.; Zhang, M.; Liao, D.; Wang, G.; Qin, G.; Xu, R.; Kang, T. CBX4 Suppresses Metastasis via Recruitment of HDAC3 to the Runx2 Promoter in Colorectal Carcinoma. Cancer Res. 2016, 76, 7277–7289.
- Sase, T.; Suzuki, T.; Miura, K.; Shiiba, K.; Sato, I.; Nakamura, Y.; Takagi, K.; Onodera, Y.; Miki, Y.; Watanabe, M.; et al. Runt-Related Transcription Factor 2 in Human Colon Carcinoma: A Potent Prognostic Factor Associated with Estrogen Receptor. Int. J. Cancer 2012, 131, 2284–2293.
- Komori, T. Runx2, A Multifunctional Transcription Factor in Skeletal Development. J. Cell. Biochem. 2002, 87, 1–8.
- Pratap, J.; Javed, A.; Languino, L.R.; Van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. The Runx2 Osteogenic Transcription Factor Regulates Matrix Metalloproteinase 9 in Bone Metastatic Cancer Cells and Controls Cell Invasion. Mol. Cell. Biol. 2005, 25, 8581–8591.
- Boregowda, R.K.; Olabisi, O.O.; Abushahba, W.; Jeong, B.-S.; Haenssen, K.K.; Chen, W.; Chekmareva, M.; Lasfar, A.; Foran, D.J.; Goydos, J.S.; et al. RUNX2 Is Overexpressed in Melanoma Cells and Mediates Their Migration and Invasion. Cancer Lett. 2014, 348, 61–70.
- Li, X.-Q.; Du, X.; Li, D.-M.; Kong, P.-Z.; Sun, Y.; Liu, P.-F.; Wang, Q.-S.; Feng, Y.-M. ITGBL1 Is a Runx2 Transcriptional Target and Promotes Breast Cancer Bone Metastasis by Activating the TGFβ Signaling Pathway. Cancer Res. 2015, 75, 3302–3313.
- El-Gendi, S.M.; Mostafa, M.F. Runx2 Expression as a Potential Prognostic Marker in Invasive Ductal Breast Carcinoma. Pathol. Oncol. Res. 2016, 22, 461–470.
- Baniwal, S.K.; Khalid, O.; Gabet, Y.; Shah, R.R.; Purcell, D.J.; Mav, D.; Kohn-Gabet, A.E.; Shi, Y.; Coetzee, G.A.; Frenkel, B. Runx2 Transcriptome of Prostate Cancer Cells: Insights into Invasiveness and Bone Metastasis. Mol. Cancer 2010, 9, 258.
- Qin, L.; Zhang, Y.; Lin, J.; Shentu, Y.; Xie, X. MicroRNA-455 Regulates Migration and Invasion of Human Hepatocellular Carcinoma by Targeting Runx2. Oncol. Rep. 2016, 36, 3325–3332.
- Li, Y.-J.; Ping, C.; Tang, J.; Zhang, W. MicroRNA-455 Suppresses Non-Small Cell Lung Cancer through Targeting ZEB1: The Role of MiRNA-455 in NSCLC. Cell Biol. Int. 2016, 40, 621–628.
- Chai, J.; Wang, S.; Han, D.; Dong, W.; Xie, C.; Guo, H. MicroRNA-455 Inhibits Proliferation and Invasion of Colorectal Cancer by Targeting RAF Proto-Oncogene Serine/Threonine-Protein Kinase. Tumor Biol. 2015, 36, 1313–1321.
- Chou, C.-H.; Shrestha, S.; Yang, C.-D.; Chang, N.-W.; Lin, Y.-L.; Liao, K.-W.; Huang, W.-C.; Sun, T.-H.; Tu, S.-J.; Lee, W.-H.; et al. MiRTarBase Update 2018: A Resource for Experimentally Validated MicroRNA-Target Interactions. Nucleic Acids Res. 2018, 46, D296–D302.
- Zhao, W.; Zhang, S.; Wang, B.; Huang, J.; Lu, W.W.; Chen, D. Runx2 and MicroRNA Regulation in Bone and Cartilage Diseases: Runx2 and MiRNAs in Bone and Cartilage. Ann. N. Y. Acad. Sci. 2016, 1383, 80–87.
- Wang, S.-Y.; Chen, C.-L.; Hu, Y.-C.; Chi, Y.; Huang, Y.-H.; Su, C.-W.; Jeng, W.-J.; Liang, Y.-J.; Wu, J.-C. High Expression of MicroRNA-196a Is Associated with Progression of Hepatocellular Carcinoma in Younger Patients. Cancers 2019, 11, 1549.
- Wai, P.Y.; Kuo, P.C. The Role of Osteopontin in Tumor Metastasis. J. Surg. Res. 2004, 121, 228–241.
- Gou, Y.; Zhai, F.; Zhang, L.; Cui, L. RUNX3 Regulates Hepatocellular Carcinoma Cell Metastasis via Targeting MiR-186/E-Cadherin/EMT Pathway. Oncotarget 2017, 8, 61475–61486.
- Zhang, X.; He, H.; Zhang, X.; Guo, W.; Wang, Y. RUNX3 Promoter Methylation Is Associated with Hepatocellular Carcinoma Risk: A Meta-Analysis. Cancer Investig. 2015, 33, 121–125.
- Yang, Y.; Ye, Z.; Zou, Z.; Xiao, G.; Luo, G.; Yang, H. Clinicopathological Significance of RUNX3 Gene Hypermethylation in Hepatocellular Carcinoma. Tumor Biol. 2014, 35, 10333–10340.
- El-shaarawy, F.; Abo ElAzm, M.M.; Mohamed, R.H.; Radwan, M.I.; Abo-Elmatty, D.M.; Mehanna, E.T. Relation of the Methylation State of RUNX3 and P16 Gene Promoters with Hepatocellular Carcinoma in Egyptian Patients. Egypt. J. Med. Hum. Genet. 2022, 23, 48.
- Sun, G.; Zhang, C.; Feng, M.; Liu, W.; Xie, H.; Qin, Q.; Zhao, E.; Wan, L. Methylation Analysis of P16, SLIT2, SCARA5, and Runx3 Genes in Hepatocellular Carcinoma. Medicine 2017, 96, e8279.
- El-Bendary, M.; Nour, D.; Arafa, M.; Neamatallah, M. Methylation of Tumour Suppressor Genes RUNX3, RASSF1A and E-Cadherin in HCV-Related Liver Cirrhosis and Hepatocellular Carcinoma. Br. J. Biomed. Sci. 2020, 77, 35–40.
- Chen, F.; Liu, X.; Bai, J.; Pei, D.; Zheng, J. The Emerging Role of RUNX3 in Cancer Metastasis (Review). Oncol. Rep. 2016, 35, 1227–1236.
- Gao, J.; Chen, Y.; Wu, K.-C.; Liu, J.; Zhao, Y.-Q.; Pan, Y.-L.; Du, R.; Zheng, G.-R.; Xiong, Y.-M.; Xu, H.-L.; et al. RUNX3 Directly Interacts with Intracellular Domain of Notch1 and Suppresses Notch Signaling in Hepatocellular Carcinoma Cells. Exp. Cell Res. 2010, 316, 149–157.
- Chi, X.-Z.; Lee, J.-W.; Lee, Y.-S.; Park, I.Y.; Ito, Y.; Bae, S.-C. Runx3 Plays a Critical Role in Restriction-Point and Defense against Cellular Transformation. Oncogene 2017, 36, 6884–6894.
- Shiraha, H.; Nishina, S.; Yamamoto, K. Loss of Runt-Related Transcription Factor 3 Causes Development and Progression of Hepatocellular Carcinoma. J. Cell. Biochem. 2011, 112, 745–749.
- Tanaka, S.; Shiraha, H.; Nakanishi, Y.; Nishina, S.-I.; Matsubara, M.; Horiguchi, S.; Takaoka, N.; Iwamuro, M.; Kataoka, J.; Kuwaki, K.; et al. Runt-Related Transcription Factor 3 Reverses Epithelial-Mesenchymal Transition in Hepatocellular Carcinoma. Int. J. Cancer 2012, 131, 2537–2546.
- Kataoka, J.; Shiraha, H.; Horiguchi, S.; Sawahara, H.; Uchida, D.; Nagahara, T.; Iwamuro, M.; Morimoto, H.; Takeuchi, Y.; Kuwaki, K.; et al. Loss of Runt-Related Transcription Factor 3 Induces Resistance to 5-Fluorouracil and Cisplatin in Hepatocellular Carcinoma. Oncol. Rep. 2016, 35, 2576–2582.
- Xu, N.; Shen, C.; Luo, Y.; Xia, L.; Xue, F.; Xia, Q.; Zhang, J. Upregulated MiR-130a Increases Drug Resistance by Regulating RUNX3 and Wnt Signaling in Cisplatin-Treated HCC Cell. Biochem. Biophys. Res. Commun. 2012, 425, 468–472.
- Chen, W.; Gao, N.; Shen, Y.; Cen, J. Hypermethylation Downregulates Runx3 Gene Expression and Its Restoration Suppresses Gastric Epithelial Cell Growth by Inducing P27 and Caspase3 in Human Gastric Cancer. J. Gastroenterol. Hepatol. 2010, 25, 823–831.
- Wei, D.; Gong, W.; Oh, S.C.; Li, Q.; Kim, W.D.; Wang, L.; Le, X.; Yao, J.; Wu, T.T.; Huang, S.; et al. Loss of RUNX3 Expression Significantly Affects the Clinical Outcome of Gastric Cancer Patients and Its Restoration Causes Drastic Suppression of Tumor Growth and Metastasis. Cancer Res. 2005, 65, 4809–4816.
- Chi, X.-Z.; Yang, J.-O.; Lee, K.-Y.; Ito, K.; Sakakura, C.; Li, Q.-L.; Kim, H.-R.; Cha, E.-J.; Lee, Y.-H.; Kaneda, A.; et al. RUNX3 Suppresses Gastric Epithelial Cell Growth by Inducing P21WAF1/Cip1 Expression in Cooperation with Transforming Growth Factor β-Activated SMAD. Mol. Cell. Biol. 2005, 25, 8097–8107.
- Ito, Y.; Miyazono, K. RUNX Transcription Factors as Key Targets of TGF-β Superfamily Signaling. Curr. Opin. Genet. Dev. 2003, 13, 43–47.
- Yano, T.; Ito, K.; Fukamachi, H.; Chi, X.-Z.; Wee, H.-J.; Inoue, K.; Ida, H.; Bouillet, P.; Strasser, A.; Bae, S.-C.; et al. The RUNX3 Tumor Suppressor Upregulates Bim in Gastric Epithelial Cells Undergoing Transforming Growth Factorβ-Induced Apoptosis. Mol. Cell. Biol. 2006, 26, 4474–4488.
- Zeng, S.; Shen, W.; Liu, L. Senescence and Cancer. Cancer Transl. Med. 2018, 4, 70.
- Giovannini, C.; Fornari, F.; Piscaglia, F.; Gramantieri, L. Notch Signaling Regulation in HCC: From Hepatitis Virus to Non-Coding RNAs. Cells 2021, 10, 521.
- Ahn, S.; Hyeon, J.; Park, C.-K. Notchl and Notch4 Are Markers for Poor Prognosis of Hepatocellular Carcinoma. Hepatobiliary Pancreat. Dis. Int. 2013, 12, 286–294.
- Chai, M.Y.; Kou, B.X.; Fu, Z.; Wei, F.L.; Dou, S.S.; Chen, D.X.; Liu, X.N. Sorafenib regulates vascular endothelial growth factor by runt-related transcription factor-3 to inhibit angiogenesis in hepatocellular carcinoma. Zhonghua Gan Zang Bing Za Zhi 2022, 30, 770–776.
- Lee, S.H.; Bae, S.C.; Kim, K.W.; Lee, Y.M. RUNX3 Inhibits Hypoxia-Inducible Factor-1α Protein Stability by Interacting with Prolyl Hydroxylases in Gastric Cancer Cells. Oncogene 2014, 33, 1458–1467.
- Lamouille, S.; Xu, J.; Derynck, R. Molecular Mechanisms of Epithelial–Mesenchymal Transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196.
- Voon, D.C.-C.; Wang, H.; Koo, J.K.W.; Nguyen, T.A.P.; Hor, Y.T.; Chu, Y.-S.; Ito, K.; Fukamachi, H.; Chan, S.L.; Thiery, J.P.; et al. Runx3 Protects Gastric Epithelial Cells Against Epithelial-Mesenchymal Transition-Induced Cellular Plasticity and Tumorigenicity. Stem Cells 2012, 30, 2088–2099.
- Loh, C.-Y.; Chai, J.; Tang, T.; Wong, W.; Sethi, G.; Shanmugam, M.; Chong, P.; Looi, C. The E-Cadherin and N-Cadherin Switch in Epithelial-to-Mesenchymal Transition: Signaling, Therapeutic Implications, and Challenges. Cells 2019, 8, 1118.
- Lee, S.H.; Jung, Y.D.; Choi, Y.S.; Lee, Y.M. Targeting of RUNX3 by MiR-130a and MiR-495 Cooperatively Increases Cell Proliferation and Tumor Angiogenesis in Gastric Cancer Cells. Oncotarget 2015, 6, 33269–33278.
- Wang, M.; Wang, X.; Liu, W. MicroRNA-130a-3p Promotes the Proliferation and Inhibits the Apoptosis of Cervical Cancer Cells via Negative Regulation of RUNX3. Mol. Med. Rep. 2020, 22, 2990–3000.


