 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Masaru Tanaka | -- | 2894 | 2023-11-20 10:10:58 | | | |
| 2 | Mona Zou | Meta information modification | 2894 | 2023-11-21 03:10:12 | | |
Video Upload Options
Migraine is a neurovascular disorder that can be debilitating for individuals and society. Current research focuses on finding effective analgesics and management strategies for migraines by targeting specific receptors and neuropeptides. Nonetheless, newly approved calcitonin gene-related peptide (CGRP) monoclonal antibodies (mAbs) have a 50% responder rate ranging from 27 to 71.0%, whereas CGRP receptor inhibitors have a 50% responder rate ranging from 56 to 71%. To address the need for novel therapeutic targets, researchers are exploring the potential of another secretin family peptide, pituitary adenylate cyclase-activating polypeptide (PACAP), as a ground-breaking treatment avenue for migraine. Preclinical models have revealed how PACAP affects the trigeminal system, which is implicated in headache disorders. Clinical studies have demonstrated the significance of PACAP in migraine pathophysiology; however, a few clinical trials remain inconclusive: the pituitary adenylate cyclase-activating peptide 1 receptor mAb, AMG 301 showed no benefit for migraine prevention, while the PACAP ligand mAb, Lu AG09222 significantly reduced the number of monthly migraine days over placebo in a phase 2 clinical trial. Meanwhile, another secretin family peptide vasoactive intestinal peptide (VIP) is gaining interest as a potential new target.
1. Pituitary Adenylate Cyclase-Activating Peptide and Vasoactive Intestinal Peptide
2. Background
3. Receptor and Signaling Mechanisms of PACAP and VIP
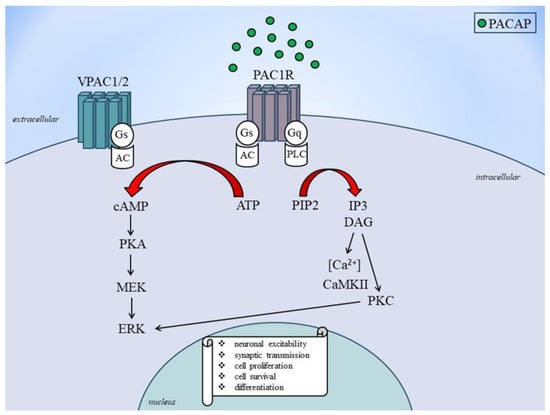
4. Role of PACAP and VIP in Migraine
5. Preclinical Studies
6. Clinical Studies
References
- Rassler, B.; Blinowska, K.; Kaminski, M.; Pfurtscheller, G. Analysis of Respiratory Sinus Arrhythmia and Directed Information Flow between Brain and Body Indicate Different Management Strategies of fMRI-Related Anxiety. Biomedicines 2023, 11, 1028.
- Arimura, A. Perspectives on pituitary adenylate cyclase activating polypeptide (PACAP) in the neuroendocrine, endocrine, and nervous systems. Jpn. J. Physiol. 1998, 48, 301–331.
- Holland, P.R.; Barloese, M.; Fahrenkrug, J. PACAP in hypothalamic regulation of sleep and circadian rhythm: Importance for headache. J. Headache Pain 2018, 19, 20.
- Maugeri, G.; D’Amico, A.G.; Musumeci, G.; Reglodi, D.; D’Agata, V. Effects of PACAP on Schwann Cells: Focus on Nerve Injury. Int. J. Mol. Sci. 2020, 21, 8233.
- Johnson, G.C.; May, V.; Parsons, R.L.; Hammack, S.E. Parallel signaling pathways of pituitary adenylate cyclase activating polypeptide (PACAP) regulate several intrinsic ion channels. Ann. N. Y. Acad. Sci. 2019, 1455, 105–112.
- Clement, A.; Guo, S.; Jansen-Olesen, I.; Christensen, S.L. ATP-Sensitive Potassium Channels in Migraine: Translational Findings and Therapeutic Potential. Cells 2022, 11, 2406.
- Miyata, A.; Arimura, A.; Dahl, R.R.; Minamino, N.; Uehara, A.; Jiang, L.; Culler, M.D.; Coy, D.H. Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells. Biochem. Biophys. Res. Commun. 1989, 164, 567–574.
- Denes, V.; Geck, P.; Mester, A.; Gabriel, R. Pituitary Adenylate Cyclase-Activating Polypeptide: 30 Years in Research Spotlight and 600 Million Years in Service. J. Clin. Med. 2019, 8, 1488.
- Vaudry, D.; Nakamachi, T.; Basille, M.; Wurtz, O.; Fournier, A.; Shoida, S. Hypophysis Adenylate Cyclase Activating Polypeptide. In Handbook of Biologically Active Peptides, 2nd ed.; Academic Press: Boston, MA, USA, 2013; Available online: https://www.sciencedirect.com/topics/pharmacology-toxicology-and-pharmaceutical-science/hypophysis-adenylate-cyclase-activating-polypeptide (accessed on 28 August 2023).
- Tam, J.K.; Lee, L.T.; Chow, B.K. PACAP-related peptide (PRP)—Molecular evolution and potential functions. Peptides 2007, 28, 1920–1929.
- Köves, K.; Szabó, E.; Kántor, O.; Heinzlmann, A.; Szabó, F.; Csáki, Á. Current State of Understanding of the Role of PACAP in the Hypothalamo-Hypophyseal Gonadotropin Functions of Mammals. Front. Endocrinol. 2020, 11, 88.
- Lee, E.H.; Seo, S.R. Neuroprotective roles of pituitary adenylate cyclase-activating polypeptide in neurodegenerative diseases. BMB Rep. 2014, 47, 369–375.
- Sadanandan, N.; Cozene, B.; Park, Y.J.; Farooq, J.; Kingsbury, C.; Wang, Z.J.; Moscatello, A.; Saft, M.; Cho, J.; Gonzales-Portillo, B.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide: A Potent Therapeutic Agent in Oxidative Stress. Antioxidants 2021, 10, 354.
- Waschek, J.A. VIP and PACAP: Neuropeptide modulators of CNS inflammation, injury, and repair. Br. J. Pharmacol. 2013, 169, 512–523.
- Xiao, X.; Qiu, P.; Gong, H.Z.; Chen, X.M.; Sun, Y.; Hong, A.; Ma, Y. PACAP ameliorates hepatic metabolism and inflammation through up-regulating FAIM in obesity. J. Cell Mol. Med. 2019, 23, 5970–5980.
- Gonkowski, S. Vasoactive Intestinal Polypeptide in the Carotid Body—A History of Forty Years of Research. A Mini Review. Int. J. Mol. Sci. 2020, 21, 4692.
- Jiang, W.; Wang, H.; Li, Y.S.; Luo, W. Role of vasoactive intestinal peptide in osteoarthritis. J. Biomed. Sci. 2016, 23, 63.
- Cao, S.G.; Wu, W.C.; Han, Z.; Wang, M.Y. Effects of psychological stress on small intestinal motility and expression of cholecystokinin and vasoactive intestinal polypeptide in plasma and small intestine in mice. World J. Gastroenterol. 2005, 11, 737–740.
- Jacobs, B.; Dussor, G. Neurovascular contributions to migraine: Moving beyond vasodilation. Neuroscience 2016, 338, 130–144.
- Langer, I.; Jeandriens, J.; Couvineau, A.; Sanmukh, S.; Latek, D. Signal Transduction by VIP and PACAP Receptors. Biomedicines 2022, 10, 406.
- Fizanne, L.; Sigaudo-Roussel, D.; Saumet, J.L.; Fromy, B. Evidence for the involvement of VPAC1 and VPAC2 receptors in pressure-induced vasodilatation in rodents. J. Physiol. 2004, 554 Pt 2, 519–528.
- Parsons, R.L.; May, V. PACAP-Induced PAC1 Receptor Internalization and Recruitment of Endosomal Signaling Regulate Cardiac Neuron Excitability. J. Mol. Neurosci. 2019, 68, 340–347.
- Bill, C.A.; Vines, C.M. Phospholipase C. Adv. Exp. Med. Biol. 2020, 1131, 215–242.
- Barloese, M.; Chitgar, M.; Hannibal, J.; Møller, S. Pituitary adenylate cyclase-activating peptide: Potential roles in the pathophysiology and complications of cirrhosis. Liver Int. 2020, 40, 2578–2589.
- Makhinson, M.; Chotiner, J.K.; Watson, J.B.; O’Dell, T.J. Adenylyl cyclase activation modulates activity-dependent changes in synaptic strength and Ca2+/calmodulin-dependent kinase II autophosphorylation. J. Neurosci. 1999, 19, 2500–2510.
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912.
- Lu, J.; Piper, S.J.; Zhao, P.; Miller, L.J.; Wootten, D.; Sexton, P.M. Targeting VIP and PACAP Receptor Signaling: New Insights into Designing Drugs for the PACAP Subfamily of Receptors. Int. J. Mol. Sci. 2022, 23, 8069.
- Hirabayashi, T.; Nakamachi, T.; Shioda, S. Discovery of PACAP and its receptors in the brain. J. Headache Pain 2018, 19, 28.
- Watanabe, J. Vasoactive Intestinal Polypeptide Receptor 1. In Methods in Enzymology; Academic Press: Boston, MA, USA, 2013; Available online: https://www.sciencedirect.com/topics/medicine-and-dentistry/vasoactive-intestinal-polypeptide-receptor-1 (accessed on 28 August 2023).
- Vasoactive Intestinal Polypeptide Receptor. Autonomic Neuroscience. 2007. Available online: https://www.sciencedirect.com/topics/neuroscience/vasoactive-intestinal-polypeptide-receptor (accessed on 28 August 2023).
- May, V.; Buttolph, T.R.; Girard, B.M.; Clason, T.A.; Parsons, R.L. PACAP-induced ERK activation in HEK cells expressing PAC1 receptors involves both receptor internalization and PKC signaling. Am. J. Physiol. Cell Physiol. 2014, 306, C1068–C1079.
- Hou, X.; Yang, D.; Yang, G.; Li, M.; Zhang, J.; Zhang, J.; Zhang, Y.; Liu, Y. Therapeutic potential of vasoactive intestinal peptide and its receptor VPAC2 in type 2 diabetes. Front. Endocrinol. 2022, 13, 984198.
- Sundrum, T.; Walker, C.S. Pituitary adenylate cyclase-activating polypeptide receptors in the trigeminovascular system: Implications for migraine. Br. J. Pharmacol. 2018, 175, 4109–4120.
- Liu, J.; Wang, G.; Dan, Y.; Liu, X. CGRP and PACAP-38 play an important role in diagnosing pediatric migraine. J. Headache Pain 2022, 23, 68.
- Schytz, H.W.; Olesen, J.; Ashina, M. The PACAP receptor: A novel target for migraine treatment. Neurotherapeutics 2010, 7, 191–196.
- Kuburas, A.; Russo, A.F. Shared and independent roles of CGRP and PACAP in migraine pathophysiology. J. Headache Pain 2023, 24, 34.
- Ernstsen, C.; Christensen, S.L.; Rasmussen, R.H.; Nielsen, B.S.; Jansen-Olesen, I.; Olesen, J.; Kristensen, D.M. The PACAP pathway is independent of CGRP in mouse models of migraine: Possible new drug target? Brain 2022, 145, 2450–2460.
- Christensen, C.E.; Ashina, M.; Amin, F.M. Calcitonin Gene-Related Peptide (CGRP) and Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) in Migraine Pathogenesis. Pharmaceuticals 2022, 15, 1189.
- Anapindi, K.D.B.; Yang, N.; Romanova, E.V.; Rubakhin, S.S.; Tipton, A.; Dripps, I.; Sheets, Z.; Sweedler, J.V.; Pradhan, A.A. PACAP and Other Neuropeptide Targets Link Chronic Migraine and Opioid-induced Hyperalgesia in Mouse Models. Mol. Cell Proteomics 2019, 18, 2447–2458.
- Silvestro, M.; Iannone, L.F.; Orologio, I.; Tessitore, A.; Tedeschi, G.; Geppetti, P.; Russo, A. Migraine Treatment: Towards New Pharmacological Targets. Int. J. Mol. Sci. 2023, 24, 12268.
- Pellesi, L.; Chaudhry, B.A.; Vollesen, A.L.H.; Snoer, A.H.; Baumann, K.; Skov, P.S.; Jensen, R.H.; Ashina, M. PACAP38- and VIP-induced cluster headache attacks are not associated with changes of plasma CGRP or markers of mast cell activation. Cephalalgia 2022, 42, 687–695.
- Rasmussen, L.; Deligianni, C.; Christensen, C.E.; Karlsson, W.K.; Al-Khazali, H.M.; Van de Casteele, T.; Granhall, C.; Amin, F.M.; Ashina, M. The effect of Lu AG09222 on PACAP38- and VIP-induced vasodilation, heart rate increase, and headache in healthy subjects: An interventional, randomized, double-blind, parallel-group, placebo-controlled study. J. Headache Pain 2023, 24, 60.
- Pellesi, L.; Al-Karagholi, M.A.; De Icco, R.; Coskun, H.; Elbahi, F.A.; Lopez-Lopez, C.; Snellman, J.; Hannibal, J.; Amin, F.M.; Ashina, M. Effect of Vasoactive Intestinal Polypeptide on Development of Migraine Headaches: A Randomized Clinical Trial. JAMA Netw. Open 2021, 4, e2118543.
- Schytz, H.W.; Birk, S.; Wienecke, T.; Kruuse, C.; Olesen, J.; Ashina, M. PACAP38 induces migraine-like attacks in patients with migraine without aura. Brain 2009, 132 Pt 1, 16–25.
- Vu, J.P.; Germano, P.M.; Pisegna, J.R. Adenylate Cyclase. Adenylate Cyclases Are Enzymes That Catalyze the Conversion of ATP to cAMP and Pyrophosphate. In From: The Senses: A Comprehensive Reference; Academic Press: Boston, MA, USA, 2008; Available online: https://www.sciencedirect.com/topics/agricultural-and-biological-sciences/adenylate-cyclase (accessed on 29 August 2023).
- Roberts, R.E. The extracellular signal-regulated kinase (ERK) pathway: A potential therapeutic target in hypertension. J. Exp. Pharmacol. 2012, 4, 77–83.
- Lund, A.M.; Hannibal, J. Localization of the neuropeptides pituitary adenylate cyclase-activating polypeptide, vasoactive intestinal peptide, and their receptors in the basal brain blood vessels and trigeminal ganglion of the mouse CNS; an immunohistochemical study. Front. Neuroanat. 2022, 16, 991403.
- Ivic, I.; Balasko, M.; Fulop, B.D.; Hashimoto, H.; Toth, G.; Tamas, A.; Juhasz, T.; Koller, A.; Reglodi, D.; Solymár, M. VPAC1 receptors play a dominant role in PACAP-induced vasorelaxation in female mice. PLoS ONE 2019, 14, e0211433.
- VIP and PACAP Receptors—IUPHAR/BPS Guide to PHARMACOLOGY. Available online: https://www.guidetopharmacology.org/GRAC/FamilyDisplayForward?familyId=67 (accessed on 29 August 2023).
- Datki, Z.; Sinka, R. Translational biomedicine-oriented exploratory research on bioactive rotifer-specific biopolymers. Adv. Clin. Exp. Med. 2022, 31, 931–935.
- Palotai, M.; Telegdy, G.; Tanaka, M.; Bagosi, Z.; Jászberényi, M. Neuropeptide AF induces anxiety-like and antidepressant-like behavior in mice. Behav. Brain Res. 2014, 274, 264–269.
- Lieb, A.; Thaler, G.; Fogli, B.; Trovato, O.; Posch, M.A.; Kaserer, T.; Zangrandi, L. Functional Characterization of Spinocerebellar Ataxia Associated Dynorphin A Mutant Peptides. Biomedicines 2021, 9, 1882.
- Skobeleva, K.; Shalygin, A.; Mikhaylova, E.; Guzhova, I.; Ryazantseva, M.; Kaznacheyeva, E. The STIM1/2-Regulated Calcium Homeostasis Is Impaired in Hippocampal Neurons of the 5xFAD Mouse Model of Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 14810.
- Martos, D.; Tuka, B.; Tanaka, M.; Vécsei, L.; Telegdy, G. Memory Enhancement with Kynurenic Acid and Its Mechanisms in Neurotransmission. Biomedicines 2022, 10, 849.
- Tanaka, M.; Szabó, Á.; Spekker, E.; Polyák, H.; Tóth, F.; Vécsei, L. Mitochondrial Impairment: A Common Motif in Neuropsychiatric Presentation? The Link to the Tryptophan–Kynurenine Metabolic System. Cells 2022, 11, 2607.
- Tanaka, M.; Bohár, Z.; Martos, D.; Telegdy, G.; Vécsei, L. Antidepressant-like effects of kynurenic acid in a modified forced swim test. Pharmacol. Rep. 2020, 72, 449–455.
- Tanaka, M.; Telegdy, G. Involvement of adrenergic and serotonergic receptors in antidepressant-like effect of urocortin 3 in a modified forced swimming test in mice. Brain Res. Bull. 2008, 77, 301–305.
- Tanaka, M.; Spekker, E.; Szabó, Á.; Polyák, H.; Vécsei, L. Modelling the neurodevelopmental pathogenesis in neuropsychiatric disorders. Bioactive kynurenines and their analogues as neuroprotective agents-in celebration of 80th birthday of Professor Peter Riederer. J. Neural. Transm. 2022, 129, 627–642.
- Reducha, P.V.; Edvinsson, L.; Haanes, K.A. Could Experimental Inflammation Provide Better Understanding of Migraines? Cells 2022, 11, 2444.
- Ojala, J.; Tooke, K.; Hsiang, H.; Girard, B.M.; May, V.; Vizzard, M.A. PACAP/PAC1 Expression and Function in Micturition Pathways. J. Mol. Neurosci. 2019, 68, 357–367.
- Tamas, A.; Reglodi, D.; Farkas, O.; Kovesdi, E.; Pal, J.; Povlishock, J.T.; Schwarcz, A.; Czeiter, E.; Szanto, Z.; Doczi, T.; et al. Effect of PACAP in central and peripheral nerve injuries. Int. J. Mol. Sci. 2012, 13, 8430–8448.
- Zhang, L.; Zhou, Y.; Yang, L.; Wang, Y.; Xiao, Z. PACAP6-38 improves nitroglycerin-induced central sensitization by modulating synaptic plasticity at the trigeminal nucleus caudalis in a male rat model of chronic migraine. J. Headache Pain 2023, 24, 66.
- Takács-Lovász, K.; Kun, J.; Aczél, T.; Urbán, P.; Gyenesei, A.; Bölcskei, K.; Szőke, É.; Helyes, Z. PACAP-38 Induces Transcriptomic Changes in Rat Trigeminal Ganglion Cells Related to Neuroinflammation and Altered Mitochondrial Function Presumably via PAC1/VPAC2 Receptor-Independent Mechanism. Int. J. Mol. Sci. 2022, 23, 2120.
- Frederiksen, S.D.; Haanes, K.A.; Warfvinge, K.; Edvinsson, L. Perivascular neurotransmitters: Regulation of cerebral blood flow and role in primary headaches. J. Cereb. Blood Flow Metab. 2019, 39, 610–632.
- Edvinsson, L.; Tajti, J.; Szalárdy, L.; Vécsei, L. PACAP and its role in primary headaches. J. Headache Pain 2018, 19, 21.
- Markovics, A.; Kormos, V.; Gaszner, B.; Lashgarara, A.; Szoke, E.; Sandor, K.; Szabadfi, K.; Tuka, B.; Tajti, J.; Szolcsanyi, J.; et al. Pituitary adenylate cyclase-activating polypeptide plays a key role in nitroglycerol-induced trigeminovascular activation in mice. Neurobiol. Dis. 2012, 45, 633–644.
- Edvinsson, L. PACAP and its receptors in migraine pathophysiology: Commentary on Walker et al. Br. J. Pharmacol. 171: 1521–1533. Br. J. Pharmacol. 2015, 172, 4782–4784.
- Saposnik, G.; Montalban, X.; Selchen, D.; Terzaghi, M.A.; Bakdache, F.; Montoya, A.; Fruns, M.; Caceres, F.; Oh, J. Therapeutic Inertia in Multiple Sclerosis Care: A Study of Canadian Neurologists. Front. Neurol. 2018, 9, 781.
- Harding, S.D.; Armstrong, J.F.; Faccenda, E.; Southan, C.; Alexander, S.P.H.; Davenport, A.P.; Pawson, A.J.; Spedding, M.; Davies, J.A.; NC-IUPHAR. The IUPHAR/BPS guide to PHARMACOLOGY in 2022: Curating pharmacology for COVID-19, malaria and antibacterials. Nucl. Acids Res. 2022, 50, D1282–D1294.
- Guo, S.; Jansen-Olesen, I.; Olesen, J.; Christensen, S.L. Role of PACAP in migraine: An alternative to CGRP? Neurobiol. Dis. 2023, 176, 105946.
- Granoth, R.; Fridkin, M.; Gozes, I. VIP and the potent analog, stearyl-Nle(17)-VIP, induce proliferation of keratinocytes. FEBS Lett. 2000, 475, 78–83.
- Gourlet, P.; De Neef, P.; Cnudde, J.; Waelbroeck, M.; Robberecht, P. In vitro properties of a high affinity selective antagonist of the VIP1 receptor. Peptide 1997, 18, 1555–1560.
- Beebe, X.; Darczak, D.; Davis-Taber, R.A.; Uchic, M.E.; Scott, V.E.; Jarvis, M.F.; Stewart, A.O. Discovery and SAR of hydrazide antagonists of the pituitary adenylate cyclase-activating polypeptide (PACAP) receptor type 1 (PAC1-R). Bioorg. Med. Chem. Lett. 2008, 18, 2162–2166.
- Laburthe, M.; Couvineau, A.; Tan, V. Class II G protein-coupled receptors for VIP and PACAP: Structure, models of activation and pharmacology. Peptides 2007, 28, 1631–1639.
- Tasma, Z.; Siow, A.; Harris, P.W.R.; Brimble, M.A.; Hay, D.L.; Walker, C.S. Characterisation of agonist signalling profiles and agonist-dependent antagonism at PACAP-responsive receptors: Implications for drug discovery. Br. J. Pharmacol. 2022, 179, 435–453.
- Spekker, E.; Tanaka, M.; Szabó, Á.; Vécsei, L. Neurogenic Inflammation: The Participant in Migraine and Recent Advancements in Translational Research. Biomedicines 2022, 10, 76.
- Pinho-Ribeiro, F.A.; Verri, W.A., Jr.; Chiu, I.M. Nociceptor Sensory Neuron-Immune Interactions in Pain and Inflammation. Trends Immunol. 2017, 38, 5–19.
- Guo, S.; Vollesen, A.L.; Hansen, R.D.; Esserlind, A.L.; Amin, F.M.; Christensen, A.F.; Olesen, J.; Ashina, M. Part I: Pituitary adenylate cyclase-activating polypeptide-38 induced migraine-like attacks in patients with and without familial aggregation of migraine. Cephalalgia 2017, 37, 125–135.
- ClinicalTrials.gov. PACAP Induced Migraine Attacks in Patients with High and Low Genetic Load. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT02158221 (accessed on 29 August 2023).
- Togha, M.; Ghorbani, Z.; Ramazi, S.; Zavvari, F.; Karimzadeh, F. Evaluation of Serum Levels of Transient Receptor Potential Cation Channel Subfamily V Member 1, Vasoactive Intestinal Polypeptide, and Pituitary Adenylate Cyclase-Activating Polypeptide in Chronic and Episodic Migraine: The Possible Role in Migraine Transformation. Front. Neurol. 2021, 12, 770980.
- Körtési, T.; Tuka, B.; Tajti, J.; Bagoly, T.; Fülöp, F.; Helyes, Z.; Vécsei, L. Kynurenic Acid Inhibits the Electrical Stimulation Induced Elevated Pituitary Adenylate Cyclase-Activating Polypeptide Expression in the TNC. Front. Neurol. 2018, 8, 745.
- Guo, S.; Vollesen, A.L.; Hansen, Y.B.; Frandsen, E.; Andersen, M.R.; Amin, F.M.; Fahrenkrug, J.; Olesen, J.; Ashina, M. Part II: Biochemical changes after pituitary adenylate cyclase-activating polypeptide-38 infusion in migraine patients. Cephalalgia 2017, 37, 136–147.
- Amin, F.M.; Asghar, M.S.; Guo, S.; Hougaard, A.; Hansen, A.E.; Schytz, H.W.; van der Geest, R.J.; de Koning, P.J.; Larsson, H.B.; Olesen, J.; et al. Headache and prolonged dilatation of the middle meningeal artery by PACAP38 in healthy volunteers. Cephalalgia 2012, 32, 140–149.
- Maasz, G.; Zrinyi, Z.; Reglodi, D.; Petrovics, D.; Rivnyak, A.; Kiss, T.; Jungling, A.; Tamas, A.; Pirger, Z. Pituitary adenylate cyclase-activating polypeptide (PACAP) has a neuroprotective function in dopamine-based neurodegeneration in rat and snail parkinsonian models. Dis. Model Mech. 2017, 10, 127–139.
- Rubio-Beltrán, E.; Correnti, E.; Deen, M.; Kamm, K.; Kelderman, T.; Papetti, L.; Vigneri, S.; MaassenVanDenBrink, A.; Edvinsson, L.; European Headache Federation School of Advanced Studies (EHF-SAS). PACAP38 and PAC1 receptor blockade: A new target for headache? J. Headache Pain 2018, 19, 64.
- Ashina, M.; Doležil, D.; Bonner, J.H.; Zhou, L.; Klatt, J.; Picard, H.; Mikol, D.D. A phase 2, randomized, double-blind, placebo-controlled trial of AMG 301, a pituitary adenylate cyclase-activating polypeptide PAC1 receptor monoclonal antibody for migraine prevention. Cephalalgia 2021, 41, 33–44.
- Study to Evaluate the Efficacy and Safety of AMG 301 in Migraine Prevention 2020. Available online: https://ClinicalTrials.gov/show/NCT03238781 (accessed on 4 September 2023).
- Lundbeck News Room: Lundbeck Announced the Start of a Phase II Clinical Study to Assess Lu AG09222 for Migraine Prevention. Available online: https://newsroom.lundbeckus.com/news-release/2021/lundbeck-announced-start-of-phase-ii-clinical-study-for-migraine-prevention (accessed on 29 August 2023).
- A Study With Lu AG09222 in Adults With Migraine Who Have Not Been Helped by Prior Preventive Treatments 2023. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT05133323 (accessed on 4 September 2023).
- A Study of LY3451838 in Participants with Migraine 2023. Available online: https://ClinicalTrials.gov/show/NCT04498910 (accessed on 4 September 2023).
- ClinicalTrials.gov. The Effects of a Long-Lasting Infusion of Vasoactive Intestinal Peptide (VIP) in Episodic Migraine Patients. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT04260035 (accessed on 29 August 2023).

