Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Cao, Z.; Wang, E.; Xu, X.; Tong, C.; Zhao, X.; Song, X.; Wang, J.; Wang, H.; Xia, X.; Zhao, Y. Signaling Pathway-Mediated Strategies for Plant Thermotolerance. Encyclopedia. Available online: https://encyclopedia.pub/entry/50567 (accessed on 28 June 2026).
Cao Z, Wang E, Xu X, Tong C, Zhao X, Song X, et al. Signaling Pathway-Mediated Strategies for Plant Thermotolerance. Encyclopedia. Available at: https://encyclopedia.pub/entry/50567. Accessed June 28, 2026.
Cao, Zidan, Enbo Wang, Xinyi Xu, Chenheng Tong, Xia Zhao, Xiqiang Song, Jian Wang, Hou-Ling Wang, Xinli Xia, Ying Zhao. "Signaling Pathway-Mediated Strategies for Plant Thermotolerance" Encyclopedia, https://encyclopedia.pub/entry/50567 (accessed June 28, 2026).
Cao, Z., Wang, E., Xu, X., Tong, C., Zhao, X., Song, X., Wang, J., Wang, H., Xia, X., & Zhao, Y. (2023, October 19). Signaling Pathway-Mediated Strategies for Plant Thermotolerance. In Encyclopedia. https://encyclopedia.pub/entry/50567
Cao, Zidan, et al. "Signaling Pathway-Mediated Strategies for Plant Thermotolerance." Encyclopedia. Web. 19 October, 2023.
Copy Citation
Cell membrane is mainly composed of phospholipids, which are elastic and semi-permeable membranes with a thickness of 7-8 nm. For animal cells, the outer side of the membrane is in contact with the external environment. Its main functions are to selectively exchange substances, absorb nutrients, excrete metabolic waste, secrete and transport proteins.
Reactive oxygen species (ROS) refer to the general term for oxygen-containing free radicals and peroxides that are easily formed in organisms related to oxygen metabolism. Examples include peroxides, superoxides, hydroxyl radicals, singlet oxygen, and α-oxygen.
Calmodulin (CaM) is a multifunctional protein that is widely present in various eukaryotic cells and can bind to calcium ions. Calmodulin participates in various intracellular signaling pathways and plays a key role in Ca2+ dependent signaling pathways. It is a dynamic Ca2+ sensor that can respond to a wide range of Ca2+ concentrations and transmit signals downstream.
Heat shock transcription factors (HSF) usually exist in non-stressed cells as monomers and bind to a small number of heat shock proteins. Under heat stress, they dissociate from the heat shock proteins, polymerize from monomers into trimers, move to the nucleus, and bind to the promoter sequence upstream of the heat shock gene to initiate the transcription activity of the gene, leading to an increase in heat shock proteins and thus protecting other proteins.
heat stress
signal transduction
botany
heat stress response
1. Introduction
With the greenhouse effect, more frequent and intense high temperatures have a very detrimental impact on the survival of plants. Plants with different morphological structures and physiological characteristics have different tolerance ranges for heat stress (HS). When the environmental temperature exceeds its tolerance range, extreme temperatures can adversely affect cells and inhibit their normal physiological functions [1]. When the temperature exceeds the tolerance range, plants will inhibit reproductive development, invest more energy to resist external pressure, stop seed germination, delay flower and fruit development, and even inhibit nutritional growth [1]. In order to cope with high temperatures, plants will change their morphological structure and internal physiological and biochemical processes: by curling leaves to reduce light exposure area, closing stomata to reduce water loss, increasing intracellular reactive oxygen species (ROS) to activate downstream regulatory pathways, making membrane ion channels imbalanced, and increasing various heat shock proteins to perform functions.
When plants are exposed to high temperatures for a long time, they will alter their metabolism to adapt to the environment, with stronger resistance and adaptability to high temperatures. This adaptation is called heat tolerance. Heat tolerance involves multiple biological regulatory pathways: a series of complex physiological and biochemical processes, such as the reduction of membrane lipid molecular mobility, the production and removal of reactive oxygen species, and the regulation of heat shock protein production [2,3,4].
1.1. Adaptation of Plants to High Temperatures under Different Climatic Conditions
There are many plants living on the earth. Due to different climatic conditions and growth environments, plants have formed different living habits. It can be said that the diversity of plants is caused by climate.
1.1.1. Desert Plants
The ecosystems in arid regions suffer from intense evaporation, strong winds and sand, intense sunlight and high temperatures, and infertile and salinized soils, creating extremely harsh environments. It is difficult to grow plants in this area, or the growth conditions are extremely harsh. During the evolution of plants, plant leaves are more sensitive to environmental changes and have stronger plasticity. Their structural and functional characteristics show the corresponding strategies of plants for the environment and the ability of plants to utilize resources [5]. In order to adapt to extreme drought and high salinity environments, some leaves of desert plants shrink, and even evolve into thin scales or hard spines. Some leaves are fleshy, and their shapes are mostly irregular cylinders or cones. Some leaves have strong divisions, white hairs, deep stomata, etc., to reduce water loss [6]. Some leaves have degenerated, using buds from the previous year for photosynthesis.
As a bridge for nutrient transportation between plant roots and leaves, stems have developed unique structural characteristics to adapt to extreme environments. They are wrapped in peels to suppress transpiration of plants and develop transport tissues to transport water. It consists of a large number of parenchyma cells, which improves the water retention of cells [7]. In arid regions, most plants have grown roots to absorb water from deeper soil. However, in some desert areas with more precipitation, the roots undergo significant changes: the main roots are suppressed and the lateral roots develop. This is because the shallow soil can carry more water [8]. In arid regions, there is less water, high temperatures, and more saline soil. Plants living in this region will have degraded cone-shaped leaves, reduce water loss caused by transpiration, and have more developed roots to absorb water [9]. Its molecular regulation also tends to be more resistant to salt and heat mechanisms; for example, cells contain more sodium and potassium ions.
In addition to the evolution of special forms, there are also very complex regulatory mechanisms for abiotic stress in plants at the molecular level. These responses are controlled by the expression of only a few genes, and the products of some genes are not only stress tolerance effector proteins, but also signal transduction factors and transcription factors in stress response: AREB, HSP, NAC, WRKY, MYB, ERF [10]. These gene families have been shown to regulate drought, high temperature, and salt stress in different ways. They form a large and complex network, carefully calculating the best way to survive [11].
1.1.2. Tropical Plants
Temperature at appropriate intervals affects the growth and development of plants, but extreme temperatures may endanger the life of plants. However, some plants can increase their tolerance range of temperature. In tropical regions, even during the rainy season, the temperature can exceed 40°C. Tropical plants have also evolved more comprehensively to withstand heat.
In order to adapt to high temperatures, plants have evolved many different ways and channels. Changes in plant tissue morphology are used to better adapt to high temperatures. The flat shape of leaves is designed to effectively absorb light energy, but sufficient light often comes with lethal heat, so leaves have evolved fur and scales to filter sunlight. Other plants also reflect sunlight through their leathery leaves, which can reduce the resulting excess energy and protect the plant body from heat damage. Some plant leaves are arranged vertically, allowing the leaf edges to face the light or folding the leaves under high temperature conditions to reduce the light absorption area [2]. The physiological adaptation of plants to high temperatures mainly involves reducing the water content of cells, increasing the concentration of soluble sugar or salt, which is beneficial for slowing down the metabolic rate and increasing the anticoagulant ability of the protoplasm. The second is to rely on intense transpiration to avoid damage to the plant body due to excessive heat [3].
Different plants have different defense mechanisms against high temperature stress, which vary with the duration and intensity of stress [4]. Some specific pathways are widely present in plants, such as the ROS regulatory system. Microscopic ROS act as signaling molecules to promote downstream defense responses, while excessive ROS can exacerbate membrane lipid oxidation, cause fatty acid decomposition, increase intracellular malondialdehyde (MDA) levels, and disrupt normal physiological functions of cells [12,13]. Hormones also widely regulate plant stress defense responses. The ethylene signaling pathway crosses with the heat stress response through the APETAL2/ethylene response element binding factor (AP2/ERF) transcription factor to produce heat shock proteins (HSP). Melatonin, as a signaling molecule, can clear altered intracellular free radicals (ROS/reactive nitrogen species (RNS)) in plants [14] and alter heat shock protein-dependent pathways, thereby enhancing plant resistance to abiotic stress [15]. In addition, under high temperature conditions, the content of various compounds such as proline and brassinolide in plants increases, regulating osmotic stress and improving plant heat tolerance [16]. For example, brassinolide regulates the activity of transcription factor BES1, promotes the production of heat shock proteins, and helps plants rebuild internal balance [17,18].
2. New Progress
2.1. Changes in Membrane Permeability
The plant heat stress signaling pathway is a complex collection of cellular signaling pathways and molecular mechanisms. The mutual regulation of these signaling pathways can help plants adapt and resist high temperature environments, ensuring their normal physiological functions. High temperature environments affect the stability of proteins, membranes, and cytoskeletons in plant cells to varying degrees. They also affect the activity of enzymes, altering the efficiency of enzyme catalysis, further causing metabolic disorders in plants.
The integrity and function of biological membranes are very sensitive to high temperatures, and can sense slight changes in temperature. Long-term exposure to high temperatures can alter the tertiary and quaternary structures of proteins, accelerate the movement of phospholipid molecules on the cell membrane, increase the concentration of unsaturated fatty acids, and enhance fluidity.
2.2. Transduction of Calcium Signals
When plants are exposed to high temperatures, sensitive calcium channels on the cell membrane are activated, and there is an influx of extracellular Ca2+. The influx of calcium ions can activate various downstream signaling pathways, completing the first step of the primary heat tolerance response in plants. However, in reality, calcium channels cannot directly sense temperature changes, but rather changes in plasma membrane fluidity and permeability caused by high temperatures induce channel opening. In higher plants, the use of Ca2+-chelating agent anhydrous lanthanum chloride (LaCl3) has shown that calcium channels are also validated as one of the main heat sensors [1,2,4].
Free Ca2+ is a second messenger commonly found in plants and animals, participating in various physiological reactions. When cell sensors capture changes in the external environment, open calcium channel proteins, and intracellular Ca2+ concentrations increase, the calcium sensor signal is transmitted through the calcium sensor to different substrates to different receptors, activating different regulatory pathways (Figure 1) [19].
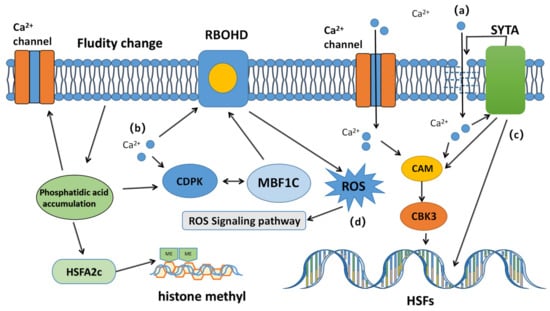
Figure 1. Schematic diagram of the calcium ion conductance signaling pathway triggered by high temperature in plants. Under heat stress, calcium ions interact with various signaling molecules. (a) High temperature changes the fluidity of the cell membrane, opens calcium channel proteins in the membrane, and activates calcium channels. Calcium ions enter the cell from membrane damage or channel proteins, and directly bind to CaM3 and calmodulin-binding kinase (CBK3) on HSF, thereby regulating the expression of HSPs. (b) Calcium ions also control ROS accumulation by directly interacting with RBOHD or calcium-dependent protein kinase (CDPK). (c) Calcium also initiates membrane repair through SYTA, and regulates the expression of CaM3, heat shock factor, and heat shock protein. (d) Calcium regulates the activity of major HSFs and ROS accumulation, initiates the expression of other HSFs and HSPs, and regulates enzyme activity, enabling plants to cope with high temperatures.
First, intracellular calcium activates calmodulin 3 (CaM3) and calcium-dependent protein kinase (CDPK).After the binding of calcium ions and calmodulin, the released signaling molecules begin to activate calmodulin-binding kinase (CBK3). Ca2+ binds to CaM3 and activates and phosphorylates heat shock factor (HSF). The heat shock element binding to the promoter region of HSP genes activates the transcription regulation of HSP production and participates in the regulation of HSP production. Multiprotein bridging factor 1c (MBF1c), as a co-activator of CDPK, stimulates the production of respiratory burst oxidase homolog D (RBOHD) (ROS) and regulates the expression of downstream antioxidant enzymes [20]. Next, the activated CDPK activates mitogen-activated protein kinases (MAPKs) and RBOHD [21],phosphorylates HSFA2 and other transcription factors, regulates the expression of HSPs, and then antagonizes HS [22].
Calcium ions also activate many other pathways to enhance heat tolerance. Synaptotagmin A (SYTA) is a Ca2+-driven transmembrane protein that repairs damaged cell membranes and regulates CaM3 expression. The intracellular calcium concentration also affects the production of heat shock molecules (HSFA2C) (Figure 1) [23]. At high temperatures, phospholipids are hydrolyzed into phosphatidic acid, which plays an important role as a second messenger in signal transduction, enhancing the expression of HSFA2C, thereby regulating the expression of HSP genes. In wheat, calmodulin is activated by calcium ions, which participate in regulating the expression of heat shock proteins and establishing stress defense [23]。At the same time, phospholipid signaling molecules can further regulate the flow of calcium ions.
2.3. The Generation of Reactive Oxygen Species (ROS) under Heat Stress
In the early stage of HS, the content of reactive oxygen species in cells gradually increases, and the expression of HSFs and the accumulation of HSPs play a decisive role in this stage. Low levels of ROS act as signaling molecules to promote downstream HS reactions, but excessive ROS accumulation can produce toxicity, and the detoxification mechanism of ROS protects plants from HT stress [24].
Small changes in temperature can significantly affect plant photosynthesis [1,25]. High ambient temperatures can inhibit the activity of photosystem Ⅱ, photosystem Ⅰ, and rubisco enzymes on plant thylakoid membranes, thereby affecting their mediated light and electron transport chains, leading to oxidative and metabolic disorders, leaving a large amount of light energy to produce more ROS [26]. Under stress conditions, ROS often play a role in the establishment of plant defense. Low levels of ROS participate in signal transduction to promote downstream reactions, while high levels of ROS cause membrane peroxidation [23],leading to cellular oxidative damage, resulting in metabolic imbalances, protein denaturation and deformation, which in turn lead to protein toxic stress, disruption of membrane stability and cytoskeletal integrity, and cell structure collapse or even programmed cell death [27].
Therefore, the signaling network of ROS needs to be highly conserved in plants to control a series of physiological and metabolic reactions. In previous studies, the focus was on the toxicity of ROS to cells. However, recent research has focused on its role as a signaling molecule. This requires plants to maintain a dynamic balance of ROS, allowing it to function as a signaling molecule without harming the plant.
Under abiotic stress, plants generate ROS signals and induce downstream responses. One of these is the activation of defense genes, leading to specific stress responses. For example, Arabidopsis thaliana upregulates the expression of antioxidant genes to enhance the antioxidant capacity of plants in response to ROS signaling transduction. However, the mechanisms involved in ROS induction remain unclear.
2.4. Production and Function of Heat Stress Protein
Heat shock proteins are key players in plant heat stress. Typically, heat stress genes (HSGs) are induced by high environmental temperatures. They further encode HSPs, which protect intracellular proteins from denaturation and maintain their stability and function through protein folding, thereby helping plants to rebuild homeostasis and resist high temperatures [28] (Figure 2).
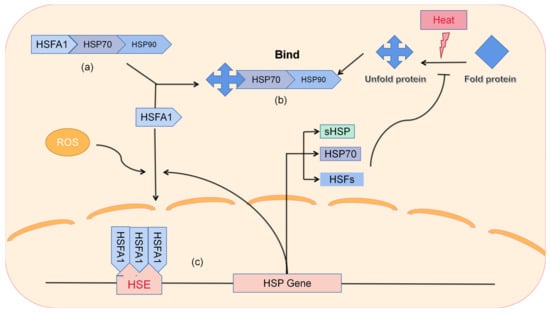
Figure 2. The working pathway of heat shock proteins. (a) Under normal conditions, HSFA1 acts as a major regulator and is sequestered by HSP70/90, inhibiting its function. (b) After high temperature stress, intracellular proteins unfold, leading to the aggregation of HSP70/90 into them and releasing HSFA1. (c) Under high ROS conditions in the cell, HSFA1 forms a trimer and binds to HSE in the nucleus, activating the expression of downstream HSPs. Induced expression of proteins can rebuild cellular homeostasis. HSE is a motif located upstream of the HSP gene promoter.
In plants, heat stress proteins are usually divided into five families based on their molecular weights: HSP100, HSP90, HSP70, HSP60, and small molecular weight HSP20 (sHSPs) [29]. HSPs can not only solve the problem of protein misfolding in cells to a certain extent, but also protect the stability of other proteins as molecular chaperones.
The function of proteins is related to their structure, and proteins with the same three-dimensional structure have the same function [24]. When the three-dimensional structure is destroyed, proteins will lose their original function. Timperio pointed out that HSP, as a molecular binding protein, can bind to substrate proteins to maintain their stability and molecular function; on the other hand, it can reduce the expression of other proteins to a certain extent [30]. QU (2013) also demonstrated how to perform the function of heat shock proteins. HSFA1, as a dominant factor, triggers the heat stress effect by producing co-activators HSFA1b and HSFA2, forming a homotrimer that binds to heat shock promoter elements and induces the expression of various HSPs, HSF70, HSF101, and sHSPs [25].
The regulatory expression of HSP is largely controlled by HSFs [31]. An HSF does not normally participate in heat tolerance (Figure 2). After changes in environmental temperature, HSFs are activated by various signaling molecules through various pathways. In the whole HSF gene family, HSFA1 is responsible for the main regulatory function. Studies have shown that if this gene is inhibited in tomatoes, the plants will not express HSPs or the expression level will be very small, affecting the normal growth and development of the plants and showing more serious symptoms of heat damage [32]. The HSF family, with HSFA1 as the main regulator, presents a complex network structure and co-regulates the expression of HSP [33,34]. Despite extensive research on HSFs, no direct upstream transcription factors with a role in thermal signal transduction have been found. Further research is needed on both HSFs and other novel transcription factors.
2.5. Heat-Resistant Mechanism of Unfolded Protein
Unfolded protein response (UPR) is a signal pathway activated in cells, which responds to the stress of damaged protein stability in the endoplasmic reticulum (ER) [33,35]. Secreted proteins and membrane proteins are folded and modified in the endoplasmic reticulum. Under high-temperature conditions, a large number of secreted proteins gather in the endoplasmic reticulum waiting for modification. When the folding capacity of the system is exceeded, a large number of unfolded proteins will accumulate, and the normal biological function of the endoplasmic reticulum will be blocked [36]. In plants, two UPR pathways have been reported, one in the ER and the other in the cytoplasm [37,38]. When the organism is subjected to stress disasters such as high temperature, high salt, and drought, the protein that needs to be folded exceeds the capacity of the system to fold, and the UPR response is activated [39]. In the process of endoplasmic reticulum folding, the regulation of cofactors is required, and the basic leucine zipper (bZIP) and transcription factors can help other proteins prevent misfolding. Under external stress, bZIP and transcription factors in the endoplasmic reticulum are released [40]. The released transcription factors enter the nucleus, activating the transcription of specific genes within the nucleus, leading to the accumulation of ER partner transcripts and the activation of brassinosteroid signaling [40]. In Arabidopsis thaliana, bZIP28 and bZIP60 are upregulated by high-temperature stress and play a role in the regulatory pathway of UPR, whereas the decrease in their expression levels will show serious high-temperature damage and reduce plant resistance [40].
Different from the UPR in the ER, the UPR in the cytoplasm is triggered by the unfolded protein in the cytoplasm, which is mainly regulated by HSF and HSFA2. HSFA2 binds to the HSF binding element in the heat shock response (HSR) gene promoter [40]. However, in the early stage of heat stress temperature rise, secreted proteins do not accumulate, and UPR response is slow, unlike calcium ion channels. However, heat stress proteins can accumulate in plant cells at warmer temperatures [41], and the activation of UPR seems to require plasmalemma to emit specific calcium signals [42], indicating that UPR acts as a downstream heat stress signal transduction rather than a heat receptor.
2.6. Carbohydrates Can Be Used as Heat Conduction Resistance Signals
In plants, as an organism’s source of carbon and energy, carbohydrates carry out important biological functions. According to research, glucose metabolism can also act as signaling molecules to regulate other physiological processes [42]. Intracellular accumulation of soluble sugars can regulate osmotic pressure, reduce abnormal ion loss, and maintain membrane stability to ensure physiological function [43,44]. Therefore, changes in glucose metabolism caused by HS can have adverse effects on sugar-sensitive systems.
In the whole development process of plants, the reproductive stage is extremely sensitive to high temperature, and even a small temperature rise will affect the development of pollen tubes [45].
At high temperatures, the resistance mechanism of plants is activated, the rate of sugar metabolism is accelerated to provide energy for the body, and the expression of glycolysis- and citric acid cycle-related proteins are increased [46]. Glucose is the most common sugar with a stable molecular structure. When the ambient temperature rises, glucose will reduce the peroxidation concentration of membrane lipids and then protect the process of photosynthesis to cope with heat stress [47]. At the same time, it was found that MAPKs in the heat-resistant signal transduction pathway were activated after being treated with Aspergillus pinosus, whereas those treated with sucrose did not respond, indicating that plants may have different signal transduction pathways under heat stress [48].
2.7. Relationship between Proline and Heat Stress
Proline is involved in the regulation of heat stress under heat stress. Many studies have shown that proline performs a variety of functions, helping to regulate osmotic pressure, remove reactive oxygen species, and stabilize cell structure [49]. Early studies have shown that the accumulation ability of proline is related to stress tolerance [50,51]. Proline is a kind of amino acid that plays an important role under stress conditions, serving as an osmotic pressure solute and signal molecule in plants [52]. In broad beans, the biosynthesis of proline and the involvement of nitric oxide and proline in the heat tolerance mechanism have been reported [52]. Proline can also effectively remove hydroxyl free radicals in cells and reduce or eliminate oxidative stress caused by reactive oxygen species under heat stress [53]. Many studies have shown that proline accumulation is positively related to the stress degree of plants, and the higher the plant stress resistance, the more proline accumulation in cells; however, some people believe that the increase in proline concentration under stress is a result of coercion and is not a response to stress [51,52,54,55]. Proline, as a molecular chaperone, regulates the integrity of proteins and enhances the activities of different enzymes. Its functions include preventing protein aggregation, clearing hydroxyl-reactive oxygen species, and regulating intracellular osmotic potential [56]. Many studies attribute antioxidant properties to proline, certificating its ability to scavenge reactive oxygen species and its role as a singlet oxygen quencher [57].
2.8. miRNAs Involved in the Construction of Plant Heat Tolerance Networks
miRNA is a small endogenous non-coding single-stranded RNA molecule that is involved in the heat-resistant defense construction of plants [1]. miRNA can be used as a signal molecule to regulate the corresponding genes of plant heat tolerance and affect plant heat tolerance. In response to heat stress, upregulated miRNAs may downregulate their specific genes, where miRNAs act as negative regulators of heat tolerance. Conversely, downregulated miRNAs may upregulate their target genes and affect heat tolerance [58].
In addition, miRNAs also play a major role in plant growth and development [59], hormone response [13], and stress defense [60]. A series of miRNAs that regulate heat stress responses have been discovered in different plants such as Arabidopsis [61]. In poplar, 12 miRNA families were identified in response to high-temperature stress. Most of their target genes were drought stress response factors or signal transduction proteins and different families had different induction patterns [62]. Under heat stress conditions, miRNA regulates a large number of stress-responsive genes, which are involved in various biological pathways for plants to acquire heat tolerance defense mechanisms [63].
Information
Subjects:
Plant Sciences
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
639
Revisions:
3 times
(View History)
Update Date:
31 Oct 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No