Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Enbo Wang and Version 1 by Enbo Wang.
气候变化引起的频繁高温对植物在不同生长阶段的生长发育产生了非常不利的影响。植物中几乎所有的细胞过程都对高温高度敏感,但植物有其独特的耐热策略。质膜通常通过膜流动性的变化来感知温度变化。受损蛋白质和活性氧在生物膜中的积累也有助于钙离子和热传感器将信号级联到转录因子,从而抵抗高温。The frequent high temperatures caused by climate change have had a very detrimental impact on the growth and development of plants at different stages of growth. Almost all cellular processes in plants are highly sensitive to high temperatures, but plants have their own unique heat-tolerant strategies. The plasma membrane usually senses temperature changes through changes in membrane fluidity. The accumulation of damaged proteins and reactive oxygen species in the biological membrane also helps calcium ions and heat sensors cascade signals to transcription factors, thereby resisting high temperatures.
- heat stress
- signal transduction
- botany
- heat stress response
1. 简介Introduction
随着温室效应,更频繁和强烈的高温对植物的生存产生了非常不利的影响。具有不同形态结构和生理特征的植物对热胁迫(HS)的耐受范围不同。当环境温度超过其耐受范围时,极端温度会对细胞产生不利影响并抑制其正常的生理功能With the greenhouse effect, more frequent and intense high temperatures have a very detrimental impact on the survival of plants. Plants with different morphological structures and physiological characteristics have different tolerance ranges for heat stress (HS). When the environmental temperature exceeds its tolerance range, extreme temperatures can adversely affect cells and inhibit their normal physiological functions [1]。当温度超过耐受范围时,植物会抑制生殖发育,投入更多的能量抵抗外界压力,种子停止发芽,花果延迟发育,甚至营养生长也会受到抑制. When the temperature exceeds the tolerance range, plants will inhibit reproductive development, invest more energy to resist external pressure, stop seed germination, delay flower and fruit development, and even inhibit nutritional growth [1]。为了应对高温,植物会改变自身的形态结构和内部生理生化过程:通过卷曲叶子减少受光面积,关闭气孔以减少水分流失,增加细胞内活性氧(ROS)激活下游调控途径,使膜上的离子通道不平衡,增加各种热休克蛋白来执行功能。
当植物长时间暴露在高温下时,它们会改变新陈代谢以适应环境,对高温有更强的抵抗力和适应能力。这种适应称为耐热性。耐热性涉及多种生物体调控途径:一系列复杂的生理生化过程,如膜脂质分子流动性的降低、活性氧的产生和去除、热休克蛋白产生的调节[. In order to cope with high temperatures, plants will change their morphological structure and internal physiological and biochemical processes: by curling leaves to reduce light exposure area, closing stomata to reduce water loss, increasing intracellular reactive oxygen species (ROS) to activate downstream regulatory pathways, making membrane ion channels imbalanced, and increasing various heat shock proteins to perform functions.
When plants are exposed to high temperatures for a long time, they will alter their metabolism to adapt to the environment, with stronger resistance and adaptability to high temperatures. This adaptation is called heat tolerance. Heat tolerance involves multiple biological regulatory pathways: a series of complex physiological and biochemical processes, such as the reduction of membrane lipid molecular mobility, the production and removal of reactive oxygen species, and the regulation of heat shock protein production [
2,3,4]。].
2. 植物在不同气候条件下对高温的适应Adaptation of Plants to High Temperatures under Different Climatic Conditions
地球上生活着许多植物。由于气候条件和生长环境的不同,植物形成了不同的生活习性,可以说植物的多样性是气候造成的。There are many plants living on the earth. Due to different climatic conditions and growth environments, plants have formed different living habits. It can be said that the diversity of plants is caused by climate.2.1. 沙漠植物Desert Plants
干旱地区的生态系统遭受强烈蒸发、强风和沙子、强烈的阳光和高温以及贫瘠和盐碱化的土壤,创造了极其恶劣的环境。在这个地区很难种植植物,或者生长条件极其恶劣。在植物进化过程中,植物叶片对环境变化更敏感,可塑性更强。它们的结构和功能特征显示了植物对环境的相应对策和植物利用资源的能力The ecosystems in arid regions suffer from intense evaporation, strong winds and sand, intense sunlight and high temperatures, and infertile and salinized soils, creating extremely harsh environments. It is difficult to grow plants in this area, or the growth conditions are extremely harsh. During the evolution of plants, plant leaves are more sensitive to environmental changes and have stronger plasticity. Their structural and functional characteristics show the corresponding strategies of plants for the environment and the ability of plants to utilize resources [5]。为了适应极端干旱和高盐度的环境,荒漠植物的一些叶子会萎缩,甚至进化成薄鳞片或硬刺。有些叶子是肉质的,形状大多是不规则的圆柱体或圆锥体。有些叶子有强烈的分裂,白色绒毛,深气孔等,以减少水分的损失. In order to adapt to extreme drought and high salinity environments, some leaves of desert plants shrink, and even evolve into thin scales or hard spines. Some leaves are fleshy, and their shapes are mostly irregular cylinders or cones. Some leaves have strong divisions, white hairs, deep stomata, etc., to reduce water loss [6]。一些叶子已经退化,使用前一年的芽进行光合作用。. Some leaves have degenerated, using buds from the previous year for photosynthesis. 作为植物根叶之间的养分运输桥梁,茎已经发展出独特的结构特征,以适应极端环境。它们被果皮包裹以抑制植物的蒸腾作用,并发展出运输组织来运输水分。它由大量的实质细胞组成,提高了细胞的保水性As a bridge for nutrient transportation between plant roots and leaves, stems have developed unique structural characteristics to adapt to extreme environments. They are wrapped in peels to suppress transpiration of plants and develop transport tissues to transport water. It consists of a large number of parenchyma cells, which improves the water retention of cells [7]。在干旱地区,大多数植物已经长出了根系,以便从更深的土壤中吸收水分。但是,在一些降水较多的沙漠地区,根系会发生很大的变化:主根会受到抑制,侧根会发育。这是因为浅层土壤可以携带更多的水分. In arid regions, most plants have grown roots to absorb water from deeper soil. However, in some desert areas with more precipitation, the roots undergo significant changes: the main roots are suppressed and the lateral roots develop. This is because the shallow soil can carry more water [8]。干旱地区的水少,温度高,盐碱土壤也比较多。生活在该地区的植物将具有退化的锥形叶子,减少蒸腾作用造成的水分损失,并具有更发达的根部来吸收水分. In arid regions, there is less water, high temperatures, and more saline soil. Plants living in this region will have degraded cone-shaped leaves, reduce water loss caused by transpiration, and have more developed roots to absorb water [9]。其分子调控也更倾向于耐盐和耐热的机制;例如,细胞含有更多的钠离子和钾离子。. Its molecular regulation also tends to be more resistant to salt and heat mechanisms; for example, cells contain more sodium and potassium ions. 除了特殊形态的演化外,植物中非生物胁迫在分子水平上也存在非常复杂的调控机制。这些反应仅由少数基因的表达控制,一些基因的产物不仅是应激耐受效应蛋白,而且是应激反应中的信号转导因子和转录因子:In addition to the evolution of special forms, there are also very complex regulatory mechanisms for abiotic stress in plants at the molecular level. These responses are controlled by the expression of only a few genes, and the products of some genes are not only stress tolerance effector proteins, but also signal transduction factors and transcription factors in stress response: AREB,HSP,NAC,WRKY,MYB,, HSP, NAC, WRKY, MYB, ERF [10]。这些基因家族已被证明以不同的方式调节干旱、高温和盐胁迫。它们形成了庞大而复杂的网络,精心计算着生存的最佳方式. These gene families have been shown to regulate drought, high temperature, and salt stress in different ways. They form a large and complex network, carefully calculating the best way to survive [11]。.2.2. 热带植物Tropical Plants
适当间隔的温度会影响植物的生长发育,但极端温度可能会危及植物的生命。然而,一些植物可以提高温度的耐受范围。在热带地区,即使在雨季,温度也超过40°C。Temperature at appropriate intervals affects the growth and development of plants, but extreme temperatures may endanger the life of plants. However, some plants can increase their tolerance range of temperature. In tropical regions, even during the rainy season, the temperature can exceed 40°C. 热带植物也进化得更全面,以抵御热量。Tropical plants have also evolved more comprehensively to withstand heat. 为了适应高温,植物进化出了许多不同的方式和渠道。植物组织形态的变化用于更好地适应高温。叶子的扁平形状旨在有效地吸收光能,但足够的光线往往伴随着致命的热量,因此叶子进化出了皮毛和鳞片来过滤阳光。其他植物也通过其革质叶子反射阳光,这可以减少随之而来的多余能量并保护植物身体免受热损伤。一些植物叶子垂直排列,使叶子边缘朝光或在高温条件下折叠叶子,减少光吸收面积In order to adapt to high temperatures, plants have evolved many different ways and channels. Changes in plant tissue morphology are used to better adapt to high temperatures. The flat shape of leaves is designed to effectively absorb light energy, but sufficient light often comes with lethal heat, so leaves have evolved fur and scales to filter sunlight. Other plants also reflect sunlight through their leathery leaves, which can reduce the resulting excess energy and protect the plant body from heat damage. Some plant leaves are arranged vertically, allowing the leaf edges to face the light or folding the leaves under high temperature conditions to reduce the light absorption area [2]。植物对高温的生理适应主要是降低细胞的含水量,增加可溶性糖或盐的浓度,有利于减慢代谢率,增加原生质的抗凝能力。二是依靠剧烈的蒸腾作用,避免因过热而对植物体造成损害. The physiological adaptation of plants to high temperatures mainly involves reducing the water content of cells, increasing the concentration of soluble sugar or salt, which is beneficial for slowing down the metabolic rate and increasing the anticoagulant ability of the protoplasm. The second is to rely on intense transpiration to avoid damage to the plant body due to excessive heat [3]。. 不同的植物对高温胁迫有不同的防御机制,这些机制随胁迫时间和强度而变化Different plants have different defense mechanisms against high temperature stress, which vary with the duration and intensity of stress [4]。一些特定的途径广泛存在于植物中,例如ROS调节系统。微量ROS作为信号分子促进下游防御反应,过量ROS会加重膜脂质氧化,引起脂肪酸分解,增加细胞内丙二醛(MDA)水平,破坏细胞正常生理功能. Some specific pathways are widely present in plants, such as the ROS regulatory system. Microscopic ROS act as signaling molecules to promote downstream defense responses, while excessive ROS can exacerbate membrane lipid oxidation, cause fatty acid decomposition, increase intracellular malondialdehyde (MDA) levels, and disrupt normal physiological functions of cells [12,13]。激素还广泛调控植物胁迫防御反应,乙烯信号转导途径通过APETAL2/乙烯响应元件结合因子(AP2/ERF)转录因子与热应激反应交叉,产生热休克蛋白(HSP)。褪黑激素作为一种信号分子,可以清除植物中改变的细胞内自由基(ROS/活性氮物种(RNS)). Hormones also widely regulate plant stress defense responses. The ethylene signaling pathway crosses with the heat stress response through the APETAL2/ethylene response element binding factor (AP2/ERF) transcription factor to produce heat shock proteins (HSP). Melatonin, as a signaling molecule, can clear altered intracellular free radicals (ROS/reactive nitrogen species (RNS)) in plants [14]和热休克蛋白依赖性途径的改变,从而增强植物非生物胁迫抵抗力 and alter heat shock protein-dependent pathways, thereby enhancing plant resistance to abiotic stress [15]。此外,在高温环境下,植物中脯氨酸和油菜素内酯等各种化合物的含量增加,调节渗透胁迫,提高植物耐热性. In addition, under high temperature conditions, the content of various compounds such as proline and brassinolide in plants increases, regulating osmotic stress and improving plant heat tolerance [16]。例如,油菜素内酯调节转录因子. For example, brassinolide regulates the activity of transcription factor BES1的活性,促进热休克蛋白的产生,帮助植物重建体内平衡, promotes the production of heat shock proteins, and helps plants rebuild internal balance [17,18]。.3. 细胞膜通透性的变化Changes in Membrane Permeability
The plant heat stress signaling pathway is a complex collection of cellular signaling pathways and molecular mechanisms. The mutual regulation of these signaling pathways can help plants adapt and resist high temperature environments, ensuring their normal physiological functions. High temperature environments affect the stability of proteins, membranes, and cytoskeletons in plant cells to varying degrees. They also affect the activity of enzymes, altering the efficiency of enzyme catalysis, further causing metabolic disorders in plants.
植物热胁迫信号通路是细胞信号通路和分子机制的复杂集合,这些信号通路的相互调控可以帮助植物适应和抵抗高温环境,保证其正常的生理功能。高温环境不同程度地影响植物细胞中蛋白质、膜和细胞骨架的稳定性。它们还影响酶的活性,改变酶催化的效率,进一步引起植物代谢紊乱。The integrity and function of biological membranes are very sensitive to high temperatures, and can sense slight changes in temperature. Long-term exposure to high temperatures can alter the tertiary and quaternary structures of proteins, accelerate the movement of phospholipid molecules on the cell membrane, increase the concentration of unsaturated fatty acids, and enhance fluidity.
生物膜的完整性和功能对高温非常敏感,可以感觉到温度的轻微变化。长时间暴露在高温下会改变蛋白质的三级和四级结构,加速细胞膜上磷脂分子的运动,增加不饱和脂肪酸的浓度,增强流动性。
4. 钙信号的传导Transduction of Calcium Signals
当植物受到高温时,细胞膜上的敏感钙通道被激活,并且存在细胞外钙When plants are exposed to high temperatures, sensitive calcium channels on the cell membrane are activated, and there is an influx of extracellular Ca2+流入。钙离子内流可以激活各种下游信号通路,完成植物初级耐热响应的第一步。然而,在现实中,钙离子通道不能直接感知温度变化,而是高温引起的质膜流动性和渗透性的变化诱导通道打开。在高等植物中,使用. The influx of calcium ions can activate various downstream signaling pathways, completing the first step of the primary heat tolerance response in plants. However, in reality, calcium channels cannot directly sense temperature changes, but rather changes in plasma membrane fluidity and permeability caused by high temperatures induce channel opening. In higher plants, the use of Ca的实验2+-螯合剂-无水氯化镧(chelating agent anhydrous lanthanum chloride (LaCl3)表明钙通道也被验证为主要的热传感器之一) has shown that calcium channels are also validated as one of the main heat sensors [1,2,4]。. 游离钙Free Ca2+是常存在于动植物中的第二信使,参与各种生理反应。当细胞传感器捕获外部环境的变化,打开钙通道蛋白,细胞内 is a second messenger commonly found in plants and animals, participating in various physiological reactions. When cell sensors capture changes in the external environment, open calcium channel proteins, and intracellular Ca浓度增加时,钙传感器传感器信号通过钙传感器传输到不同的底物到不同的接收器,从而激活不同的调节途径(图2+ concentrations increase, the calcium sensor signal is transmitted through the calcium sensor to different substrates to different receptors, activating different regulatory pathways (Figure 1)) [19]。.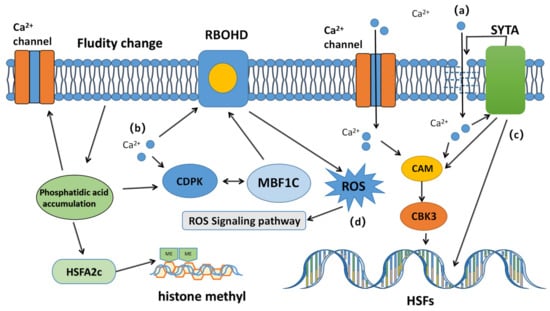
图Figure 1.高温引发植物钙离子电导信号通路示意图。在热应激下,钙离子与多种信号分子相互作用。( Schematic diagram of the calcium ion conductance signaling pathway triggered by high temperature in plants. Under heat stress, calcium ions interact with various signaling molecules. (a)高温会改变细胞膜的流动性,打开膜中的钙通道蛋白并激活钙通道。钙离子从膜损伤或通道蛋白进入细胞,并直接与HSF上的) High temperature changes the fluidity of the cell membrane, opens calcium channel proteins in the membrane, and activates calcium channels. Calcium ions enter the cell from membrane damage or channel proteins, and directly bind to CaM3 and calmodulin-binding kinase (CBK3) on HSF, thereby regulating the expression of HSPs. (b) CaM3和钙调蛋白结合激酶(CBK3)作用,进而调节Hlcium ions also control ROSP的表达。(b)钙离子还通过与 accumulation by directly interacting with RBOHD或钙依赖性蛋白激酶( or calcium-dependent protein kinase (CDPK)的直接相互作用来控制ROS积累。(). (c)钙还通过) Calcium also initiates membrane repair through SYTA启动膜修复,并调节CaM3,热休克因子和热休克蛋白的表达。(, and regulates the expression of CaM3, heat shock factor, and heat shock protein. (d)钙通过调节主要) Calcium regulates the activity of major HSF和ROS积累的活性,启动其他HSF和HSP的表达并调节酶活性,使植物能够应对高温。s and ROS accumulation, initiates the expression of other HSFs and HSPs, and regulates enzyme activity, enabling plants to cope with high temperatures.
5. 热应激下活性氧(The Generation of Reactive Oxygen Species (ROS)的产生) under Heat Stress
在HS早期,细胞中活性氧的含量逐渐增加,HSFs的表达和HSPs在这一阶段的积累起到了决定性的作用。低水平的ROS充当信号分子,促进下游HS反应,但过量的ROS积累会产生毒性,ROS的解毒机制保护植物免受HT胁迫In the early stage of HS, the content of reactive oxygen species in cells gradually increases, and the expression of HSFs and the accumulation of HSPs play a decisive role in this stage. Low levels of ROS act as signaling molecules to promote downstream HS reactions, but excessive ROS accumulation can produce toxicity, and the detoxification mechanism of ROS protects plants from HT stress [24]。. 温度的微小变化会显著影响植物的光合作用Small changes in temperature can significantly affect plant photosynthesis [1,25]。环境的高温会抑制光系统II、光系统I和植物类囊体膜上的. High ambient temperatures can inhibit the activity of photosystem Ⅱ, photosystem Ⅰ, and rubisco酶的活性,从而影响其介导的光和电子传递链,导致氧化还原和代谢紊乱,留下大量光能产生更多的活性氧(ROS) enzymes on plant thylakoid membranes, thereby affecting their mediated light and electron transport chains, leading to oxidative and metabolic disorders, leaving a large amount of light energy to produce more ROS [26].在胁迫环境下,ROS常在植物防御的建立中发挥作用。低水平的ROS参与信号转导以促进下游反应,高水平的ROS引起膜过氧化 Under stress conditions, ROS often play a role in the establishment of plant defense. Low levels of ROS participate in signal transduction to promote downstream reactions, while high levels of ROS cause membrane peroxidation [23],引起细胞氧化损伤,导致代谢失衡、蛋白质变性和变形,从而导致蛋白质毒性应激,破坏膜稳定性和细胞骨架完整性,并导致细胞结构崩溃甚至程序性死亡leading to cellular oxidative damage, resulting in metabolic imbalances, protein denaturation and deformation, which in turn lead to protein toxic stress, disruption of membrane stability and cytoskeletal integrity, and cell structure collapse or even programmed cell death [27]。. 因此,ROS的信号网络需要在植物中高度保守,以控制一系列生理代谢反应。在以前的研究中,重点是ROS对细胞的毒性。然而,最近的研究集中在它作为信号分子的方向上。这就要求植物保持ROS的动态平衡,使其在不毒害植物的情况下发挥信号分子的作用。Therefore, the signaling network of ROS needs to be highly conserved in plants to control a series of physiological and metabolic reactions. In previous studies, the focus was on the toxicity of ROS to cells. However, recent research has focused on its role as a signaling molecule. This requires plants to maintain a dynamic balance of ROS, allowing it to function as a signaling molecule without harming the plant. 在非生物胁迫下,植物产生ROS信号并诱导下游反应。其中之一是防御基因的激活,导致特定的应激反应。例如,拟南芥上调抗氧化基因的表达,提高植物响应ROS信号转导的抗氧化能力。然而,ROS诱导所涉及的机制尚不清楚。Under abiotic stress, plants generate ROS signals and induce downstream responses. One of these is the activation of defense genes, leading to specific stress responses. For example, Arabidopsis thaliana upregulates the expression of antioxidant genes to enhance the antioxidant capacity of plants in response to ROS signaling transduction. However, the mechanisms involved in ROS induction remain unclear.6. 热应激蛋白的产生及功能Production and Function of Heat Stress Protein
热休克蛋白是植物热胁迫的主要参与者。通常,热应激基因(Heat shock proteins are key players in plant heat stress. Typically, heat stress genes (HSGs) are induced by high environmental temperatures. They further encode HSG)通过环境高温诱导上调。进一步编码HSP,保护细胞内蛋白质免于变性,并通过蛋白质折叠保持其稳定性和功能,从而帮助植物重建稳态并抵抗高温s, which protect intracellular proteins from denaturation and maintain their stability and function through protein folding, thereby helping plants to rebuild homeostasis and resist high temperatures [28](图 (Figure 2)。).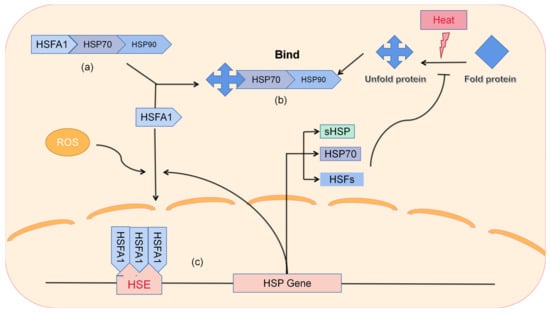
图Figure 2.热休克蛋白的工作途径。( The working pathway of heat shock proteins. (a)在正常情况下,) Under normal conditions, HSFA1作为主要调节剂,被 acts as a major regulator and is sequestered by HSP70/90螯合,抑制其功能。(, inhibiting its function. (b)高温应激后,细胞内蛋白质展开,导致) After high temperature stress, intracellular proteins unfold, leading to the aggregation of HSP70/90聚集到其中,释放HSFA1。( into them and releasing HSFA1. (c)在细胞内高) Under high ROS条件下,HSFA1形成三聚体并与细胞核中的HSE结合,激活下游HSP的表达。 conditions in the cell, HSFA1 forms a trimer and binds to HSE in the nucleus, activating the expression of downstream HSPs. Induced expression of proteins can rebuild cellular homeostasis. 蛋白质的诱导表达可以重建细胞稳态。HSE是位于HSP基因启动子上游的基序。HSE is a motif located upstream of the HSP gene promoter.