 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Laurent Dufossé | -- | 9230 | 2023-10-06 08:39:35 | | | |
| 2 | Peter Tang | Meta information modification | 9230 | 2023-10-07 05:07:54 | | | | |
| 3 | Peter Tang | Meta information modification | 9230 | 2023-10-07 05:09:14 | | |
Video Upload Options
Anthraquinones and their derivatives constitute a large group of quinoid compounds with about 700 molecules described. They are widespread in fungi and their chemical diversity and biological activities attracted attention of industries in such fields as pharmaceuticals, clothes dyeing, and food colorants. As marine microorganisms appeared as producers of an astonishing variety of structurally unique secondary metabolites, they may represent a promising resource for identifying new candidates for therapeutic drugs or daily additives.
1. Introduction
2. Anthraquinones from Marine-Derived Fungi
2.1. Anthraquinone′s Basic Structure
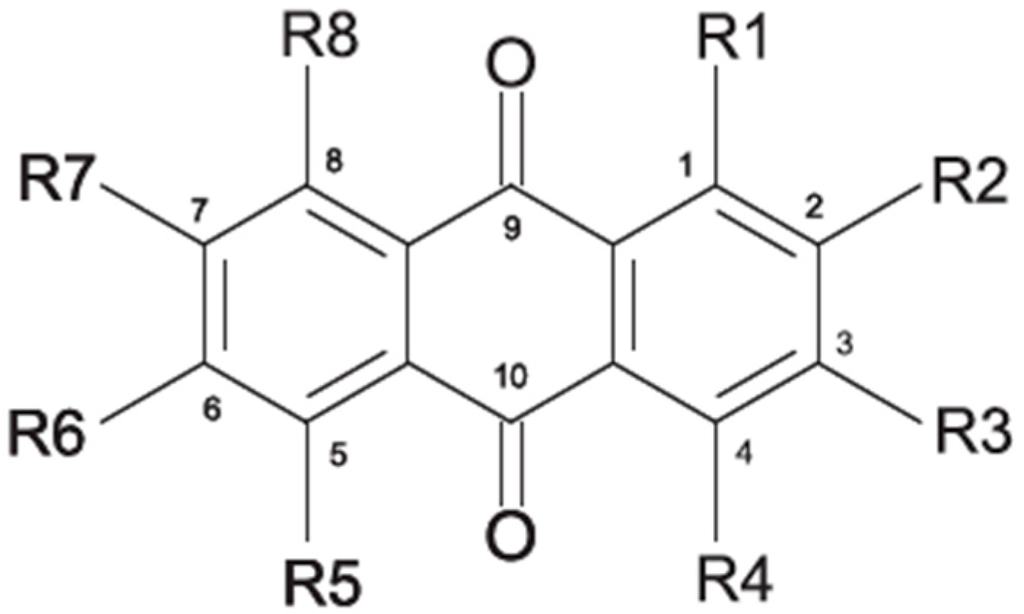
2.2. Ecology of Marine-Derived Fungal Anthraquinones Producers
-
obligate marine fungi (true ones) that grow and sporulate only in seawater. Their spores are able to germinate and form new thalli in salted environment.
-
transitional marine fungi (marine-derived fungi) that come from terrestrial or freshwater media and have undergone physiological adaptation to survive, grow, or reproduce in the marine environment.

2.3. Structural Diversity and Colors of Anthraquinoid Extrolites from Marine-Derived Fungi
2.3.1. Present Knowledge about Anthraquinonoid Compounds from Fungi
2.3.2. Nature and Colors of Compounds from Marine-Derived Fungi
Genera and Species
|
Genus |
Species/Strain No |
Name of Compounds Produced |
Source of Isolation |
Refs. |
|---|---|---|---|---|
|
Alternaria |
Al. eichorniae |
4-deoxyBostrycin, Bostrycin |
Mar. Plant pathogen |
[46] |
|
Al. (SK11) |
(+) α S-alterporriol C, 6-methylquinizarin, Alterporriol S, Austrocortinin |
Mangrove Plant end. |
[47] |
|
|
Al. sp. ZJ-2008003 |
Alterporriol C, K–R, Altersolanol B and C, Macrosporin |
Mar. Org end. |
[48] |
|
|
Al. sp. ZJ9-6B |
Alterporriols C–M, Altersolanol A, Dactylariol, Macrosporin, Physcion, TetrahydroAltersolanol B |
Mar. Plant end. |
[49] |
|
|
Aspergillus |
A. glaucus |
10,10′-dimer of Emodin and Physcion, Catenarin, Cynodontin, Emodin, Erythroglaucin, Helminthosporin, Physcion, Questin, Rubrocristin, Tritisporin, Variecolorquinone A |
Mangrove sed. |
|
|
A. sp. 05F16 |
1-deoxytetrahydrobostrycin, Tetrahydrobostrycin |
Algal end. |
[54] |
|
|
A. sp. SCSIOF063 |
(1′S)-7-chloroaverantin, 1′-O-methylaverantin 1′-O-methyl-7-chloroaverantin, 6-O-methyl-7-chloroaverantin, 6-O-methyl-7-chloroaverythrin, 6-O-methyl-7-bromoaverantin, 6,1′-O,O-dimethyl-7-chloroaverantin, 6,1′-O,O-dimethyl-7-bromoaverantin, 6,1′-O,O-dimethylaverantin, 7-chloroaverantin-1′-butyl ether, 7-chloroaverythrin |
Sed. |
[55] |
|
|
A. variecolor B-17 |
(2S)-2,3-dihydroxypropyl1,6,8-trihydroxy-3-methyl-9,10-dioxoanthracene-2carboxylate, Catenarin, Emodin, Fallacinol, Physcion, Erythroglaucin, Questin, Questinol, Rubrocristin, Variecolorquinone A, |
Sed. |
[56] |
|
|
A. versicolor |
7-hydroxyemodin 6,8-methyl ether, Emodin, Isorhodoptilometrin-methyl ether, Methyl emodin |
Algal end. |
[57] |
|
|
A. versicolor EN-7 (Genbank no EU042148) |
6,8-di-O-methylversiconol 6,8-di-O-methylnidurufin 6,8-di-O-methylaverantin 6,8-di-O-methylversicolorin A, Aversin: (−)-isomer |
Algal end. |
[58] |
|
|
Curvularia |
C. lunata |
Cytoskyrin A, Lunatin |
Mar. Org end. |
|
|
Eurotium |
E. cristatum (ECE) |
Catenarin, Emodin, Erythroglaucyn, Physcion, Physcion anthrone, Questin, Rubrocristin |
Mar. Org end. |
|
|
E. repens |
Catenarin, Erythroglaucyn, Physcion, Physcion anthrone |
Mar. Org end. |
||
|
E. rubrum |
6,3-O-(α-d-ribofuranosyl)-questin, Questin |
Mar. Plant end. |
[63] |
|
|
Unidentified |
Fungus Isolate 1850 and 2526 |
Averufin, Nidurufin, versicolorin C |
Mar. Plant end. |
[64] |
|
Fungus ZSUH-36 |
1′-O-methyl averantin, 6,8-di-O-methyl averufanin, 6,8-di-O-methyl averufin, 6,8,1′-tri-O-methyl averantin, Versicolorin C |
Mar. Plant end. |
[65] |
|
|
Fusarium |
F. sp. No. B77 |
5-acetyl-2-methoxy-1,4,6-trihydroxy-anthraquinone |
Mangrove Plant end. |
[66] |
|
F. sp. ZZF60 |
6,8-dimethoxy-1-methyl-2-(3-oxobutyl) anthraquinone |
Mangrove Plant end. |
[67] |
|
|
F. sp. No. ZH-210 |
Fusaquinon A,B,C |
Mangrove sed. |
[68] |
|
|
F. sp. PSU-F14, F. sp. PSU-F135 |
Austrocortirubin, Bostrycin |
Mar. Org end. |
[69] |
|
|
Halorosellinia |
H. sp. (No. 1403) |
1,4,5,6,7,9-hexahydroxy-2-methoxy-7-methyl-5β,9β, 8aβ,6α,10aα—hexahydroanthracene10(10aH)-one, Austrocortirubin, Demethoxyaustrocortirubin, Hydroxy-9,10-anthraquinone, SZ-685C |
Mar. Plant-derived |
|
|
Lichens |
Arthonia elegans, Biatorella conspersa, B. ochrophora, Pyrenula cerina, Sphaerophorus fragilis, Stereocaulon corticatulum,v. procerum, Trypethelium aeneum, T. aureomaculata, etc. |
Physcion |
Lichens |
[72] |
|
Caloplaca sp. |
Phallacinol (=Teloschistin=Fallacinol)) |
Lichen |
||
|
Caloplaca ehrenbergii, C. schaereri, C. spitsbergensis, etc. |
1-O-methyl-7-chloroemodin, 7-chloro-1,6,8-trihydroxy-3-methyl-10-anthrone, 7-chlorocitreorosein, 7-chloroemodic acid, 7-chloroemodin, 7-chloroemodinal, Emodin, Phallacinol, Physcion |
Lichens |
||
|
Gliocladium sp. T 31 |
Citreorosein, Emodin, Isorhodoptilometrin |
Lichen |
[78] |
|
|
Letrouitia hafellneri, L. leprolytoides |
7-chloroemodinal, 7-chloroemodin, Fragilin, Physcion |
Lichens |
||
|
Microsphaeropsis |
M. sp. |
1,3,6,8-tetrahydroxyanthraquinone, 1,3,6,8-tetrahydroxy-2-(1-hydroxyethyl)anthraquinone 1,3,6,8-tetrahydroxy-2-(1-methoxyethyl)anthraquinone 1,2,3,6,8-pentahydroxy-7-(1-methoxyethyl)anthraquinone |
Mar. Org end. |
[81] |
|
Monodictys |
M. sp. |
Chrysophanol, Emodin, Monodictyquinone A, Pachybasin |
Mar. Org end. |
|
|
Nigrospora |
N. spp. |
1-deoxytetrahydrobostrycin, 4-deoxybostrycin, Bostrycin, 4a-epi-9α-methoxydihydrodeoxybostrycin, 10-deoxybostrycin |
Mar. Plant/Org end. |
|
|
N. sp. MA75 |
4-deoxybostrycin, Bostrycin |
Marine |
[85] |
|
|
N. sp. 1403 |
4-deoxybostrycin, Bostrycin |
Mangrove |
[86] |
|
|
Paecilomyces |
P. sp. (Tree 1-7) |
Chrysophanol, Emodin |
Mangrove |
[87] |
|
Penicillium |
P. citrinum PSU-F51 (Accession no JQ66600) |
Chrysophanol, Citreorosein, Emodin, Penicillanthranins A and B |
Mar. Org end. |
[88] |
|
P. chrysogenum |
Skyrin |
Salt lake |
[89] |
|
|
P. flavidorsum SHK1-27 |
6,8-O-dimethylaverufin, 8-O-methylaverufin, Averufin, Averantin, Versiconol, Versicolorin A&B, Nidurufin |
Marine |
[90] |
|
|
P. oxalicum 2-HL-M-6 |
Aloe emodin, Chrysophanol, Citreorosein, Citreorosein-3-O-sulfate, Emodin, Emodin-3-O-sulfate, Isorhodoptilometrin |
Mangrove sed. |
[91] |
|
|
Phomopsis |
P. sp. PSU-MA214 |
Phomopsanthraquinone, 1-hydroxy-3-methoxy-6-methylanthraquinone, Ampelanol, Macrosporin |
Mangrove Plant endo |
[92] |
|
Stemphylium |
S. sp. 33231 |
2-O-acetylaltersolanol B, Alterporriol T–W, Altersolanol B&C, Auxarthrol C, Macrosporin |
Mangrove Plant end. |
[93] |
|
S. globuliferum |
6-O-methylalaternin, Acetylalterporriol D and E, Alterporriol D and E, Altersolanol A,B and C, Dihydroaltersolanol B and C, Macrosporin, Stemphylanthranol A and B |
Salt lake Plant end. |
[94] |
|
|
Trichoderma |
T. aureoviride PSU-F95 |
Coniothranthraquinone 1, Trichodermaquinone |
Mar. Org end. |
[88] |
|
Xylaria |
X. sp. 2508 |
Altersolanol A, Bostrycin, Deoxybostrycin, Xylanthraquinone |
Marine |
[95] |
| Mol. Formula/Mol. mass | Trivial name | Structure | IUPAC NAME | Source | Refs |
|---|---|---|---|---|---|
| C14H8O3/224 | Hydroxy-9,10-anthraquinone |
 |
1-hydroxy-3-methylanthraquinone | Halorosellinia sp. No. 1403 | [70] |
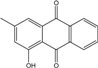
| C15H10O3/238 | Pachybasin |
 |
1-hydroxy-3-methylanthraquinone | Monodictys sp. | [14][82] |
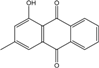
| C15H10O4/254 | Chrysophanol |
 |
1,8-dihydroxy-3-methylanthraquinone | Monodictys sp. Paecilomyces sp. P. citrinum PSU-F51 P. oxalicum 2-HL-M-6 | [14][82] [87] [88] [91] |
| C15H10O4/254 | 6-Methylquinizarin | 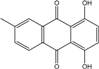 |
1,4-dihydroxy-methylanthraquinone | Al. sp. (SK11) | [47] |
| C16H12O4/268 | 1-Hydroxy-3-methoxy-6-methylanthraquinone | 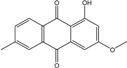 |
---- | Phomopsis sp. PSU-MA214 | [92] |
| C15H10O5/270 | Aloe emodin |
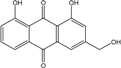 |
1,8-dihydroxy-3-(hydroxymethyl)anthraquinone | P. oxalicum 2-HL-M-6 | [91] |
| C15H10O5/270 | Emodin |
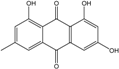 |
1,3,8-trihydroxy-6-methylanthraquinone | A. glaucus A. variecolor B-17 A. versicolor Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) Eurotium Cristatum Gliocladium sp. T31 Monodictys sp. Paecilomyces sp. P. citrinum PSU-F51 P. oxalicum 2-HL-M-6 | [91] [56] [61] [75][76][77][79][96] [36] [78] [14][82] [87] [88] [57] |
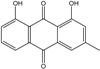
| C15H10O5/270 | Helminthosporin |
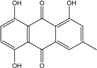 |
1,5,8-trihydroxy-3-methylanthraquinone | A. glaucus | [50][51][52][53] |
| C14H8O6/272 | 1,3,6,8-Tetrahydroxyanthraquinone |
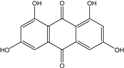 |
---- | A.versicolor Microsphaeropsis | [97] [81] |
| C15H14O5/274 | Coniothranthraquinone 1 | 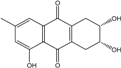 |
(2S,3R)-2,3,5-trihydroxy-7-methyl-1,2,3,4-tetrahydroanthraquinone | Trichoderma aureoviride (PSU-F95) | [88][98] |
| C16H12O5/284 | 1-Methylemodin |
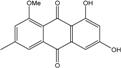 |
1,3-dihydroxy-8-methoxy-6-methylanthraquinone | A. versicolor | [57] |
| C16H12O5/284 | Austrocortinin |
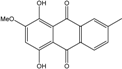 |
1,4-dihydroxy-2-methoxy-7-methylanthraquinone | Al. sp. (SK11) | [47] |
| C16H12O5/284 | Macrosporin |
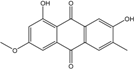 |
1,7-dihydroxy-3-methoxy-6-methylanthraquinone | Al. sp. ZJ9-6B Al. sp. ZJ-2008003 Phomopsis sp. PSU-MA214 Stemphylium globuliferum Stemphylium sp. 33231 | [49] [48] [92] [94] [93] |
| C16H12O5/284 | Marcrospin | 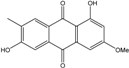 |
1,6-dihydroxy-3-methoxy-7-methylanthraquinone | Al. sp. ZJ9-6B | [49] |
| C16H12O5/284 | Monodictyquinone A |
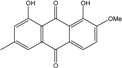 |
1,8-dihydroxy-2-methoxy-6-methylanthraquinone | Monodictys sp. | [14][82] |
| C16H12O5/284 | Phallacinol/Fallacinol |
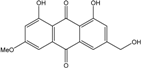 |
1,8-dihydroxy-3-(hydroxy-methyl)-6-methoxyanthraquinone | A. variecolor B-17 Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) | [99] [73][74][76][77][79][96] |
| C16H12O5/284 | Physcion |
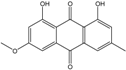 |
1,8-dihydroxy-3-methoxy-6-methylanthraquinone | Al. sp. ZJ9-6B A. glaucus A. variecolor B-17 Eurotium repens Eurotium cristatum Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) Letrouitia hafellneri, L. leprolytoides, Arthonia elegans, Biatorella conspersa, B. ochrophora, Pyrenula cerina, Sphaerophorus fragilis, Stereocaulon corticatulum, v. procerum, Trypethelium aeneum, T. aureomaculata | [49] [50][51][52][53] [56] [62] [36][61] [75][76][77][79][96] [72][80] |
| C16H12O5/284 | Questin |
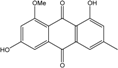 |
1,6-dihydroxy-8-methoxy-3-methylanthraquinone | A. glaucus A. variecolor B-17 Eurotium cristatum (ECE) Eurotium rubrum | [50],[51] [56] [52][53] [36][61][63] |
| C15H10O6/286 | Catenarin |
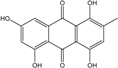 |
1,4,5,7-tetrahydroxy-2-methylanthraquinone | A. glaucus A. variecolor B-17 Eurotium cristatum (ECE) Eurotium repens | [50][51] [56] [52][53] [36][62] |
| C15H10O6/286 | Citreorosein |
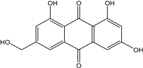 |
ω-hydroxyemodin (OHM) or 1,3,8-trihydroxy-6-(hydroxymethyl) anthraquinone | Gliocladium. sp. T31 P. citrinum PSU-F51 P. oxalicum 2-HL-M-6 | [78] [88] [91] |
| C15H10O6/286 | Cynodontin |
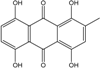 |
1,4,5,8-tetrahydroxy-2-methylanthraquinone | A. glaucus | [50][51][52][53] |
| C15H10O6/286 | Lunatin | 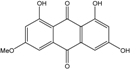 |
1,3,8-trihydroxy-6-methoxyanthraquinone | Curvularia lunata | [50][51][59][60] |
| C15H10O7/286 | Tritisporin |
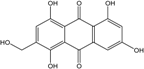 |
1,4,5,7-tetrahydroxy-2-(hydroxylmethyl) anthraquinone | A. glaucus | [50][51][52][53] |
| C15H14O6/290 | Trichodermaquinone | 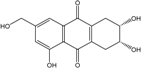 |
(2S,3R)-2,3,5-trihydroxy-7-(hydroxylmethyl)-1,2,3,4-tetrahydroanthraquinone | Trichoderma aureoviride (PSU-F95) | [88][98] |
| C15H14O6/290 | Demethoxyaustrocortirubin |
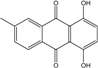 |
1,4-dihydroxy-6-methylanthraquinone | Halorosellinia sp. No. 1403 | [70][71] |
| C15H14O6/290 | 7-Hydroxyemodin 6,8-methyl ether | 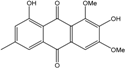 |
2,8-dihydroxy-1,3-dimethoxy-6-methyl anthraquinone | A. versicolor | [57] |
| C16H10O4/300 | Erythroglaucin |
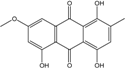 |
1, 4, 5-trihydroxy-7-methoxy-2-methylanthraquinone | A. glaucus A. variecolor B-17 Eurotium cristatum (ECE) Eurotium repens | [50][51][52][53] [56] [36][61] [36] |
| C16H12O6/300 | Carviolin | 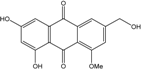 |
1,3-dihydroxy-6-(hydroxymethyl)-8-methoxyanthraquinone | P. dravuni | [100] |
| C16H12O6/300 | Questinol | 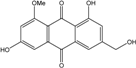 |
1,6-dihydroxy-3-(hydroxymethyl)-8-methoxyanthraquinone | A. variecolor B-17 | [56] |
| C16H12O6/300 | Rubrocristin |
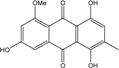 |
1,4,7-trihydroxy-5-methoxy-2-methylanthraquinone | A. glaucus A. variecolor B-17 Eurotium cristatum (ECE) | [50][51][52][53] [56] [36][61] |
| C16H12O6/300 | 6-O-methylalaternin |
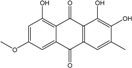 |
1,2,8-trihydroxy-6-methoxy-3-methylanthraquinone | Stemphylium globuliferum | [94] |
| C16H12O6/300 | 1,4,6-Trihydroxy-2-methoxy-7-methyl-anthraquinone |
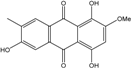 |
3,5,8-trihydroxy-7-methoxy-2-methylanthraquinone | Halorosellinia sp. No. 1403 | [70] |
| C15H9O5Cl/304 | 7-Chloroemodin | 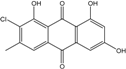 |
---- | Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) | [75][76][77][79][96] |
| C16H16O6/304 | Altersolanol B |
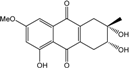 |
(2S,3R)-2,3,5-trihydroxy-7-methoxy-2-methyl-1,2,3,4-tetrahydroanthraquinone | Al. sp. ZJ-2008003 Stemphylium sp. 33231 | [48] [93] |
| C16H18O6/306 | Fusaquinon A | 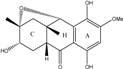 |
(2R,3S,4aR,9S,9aS)-3,5,8-trihydroxy-7-methoxy-2-methyl-2,3,4,4a,9,9a-hexahydro-2,9-epoxyanthracen-10(1H)-one | Fusarium sp. No. ZH-210 | [68] |
| C16H18O6/307 | Dihydroaltersolanol B |
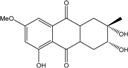 |
(2S,3R)-2,3,5-trihydroxy-7-methoxy-2-methyl-1,2,3,4,4a,9a-hexahydroanthraquinone | Stemphylium globuliferum | [94] |
| C15H19O7/311 | Xylanthraquinone | 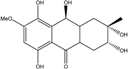 |
---- | Xylaria sp. 2508 | [95] |
| C17H14 O6/314 | Isorhodoptilometrin |
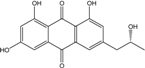 |
(R)-1,3,8-trihydroxy-6-(2-hydroxypropyl)anthraquinone | Gliocladium sp. T31 P. oxalicum 2-HL-M-6 | [78] [91] |
| C16H12O7/317 | 1,3,6,8-Tetrahydroxy-2-(1-hydroxyethyl) anthraquinone |
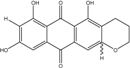 |
1,3,6,8-tetrahydroxy-2-(1-hydroxyethyl) anthracene-9,10-dione | Microsphaeropsis | [81] |
| C16H11O5Cl/318 | 1-O-Methyl-7-chloroemodin | 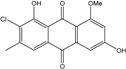 |
2-chloro-1,6-dihydroxy-8-methoxy-3-methylanthraquinone | Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) | [75][76][77][79][96] |
| C15H9O6Cl/320 | 7-Chlorocitreorosein | 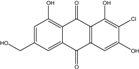 |
---- | Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) | [75][76][77][79][96] |
| C16H16O7/320 | Austrocortirubin |
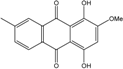 |
1,4-dihydroxy-2-methoxy-7-methylanthraquinone | Fusarium spp. PSU-F14 and PSU-F135 Halorosellinia sp. No. 1403 Nigrospora sp. ZJ-2010006 | [69] [70][71] [84] |
| C16H16O7/320 | Altersolanol C |
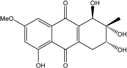 |
(1R,2R,3R)-1,2,3,5-tetra-hydroxy-7-methoxy-2-methyl-1,2,3,4-tetra-hydroanthraquinone | Al. sp. ZJ9-6B Al. sp. ZJ-2008003 Stemphylium sp. 33231 | [49] [48] [93] |
| C16H16O7/320 | 4-Deoxybostrycin |
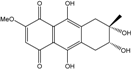 |
(2R,3S,4aS,9aS,10R)-2,3,5,8,10-pentahydroxy-6-methoxy-3-methyl-1,3,4,4a,9a,10-hexahydroanthracen-9(2H)-one | Nigrospora sp. 1403 Nigrospora sp. MA75 | [68][83][84][86][101] [85] |
| C16H16O7/320 | 2,3,5,8-Tetrahydroxy-7-methoxy-2-methyl-1,2,3,4-tetrahydroanthraquinone | ---- | 2,3,5,8-tetrahydroxy-7-methoxy-2-methyl-1,2,3,4-tetrahydroanthracene-9,10-dione | Al. eichorniae | [46] |
| C16H16O7/321 | 10-Deoxybostrycin |
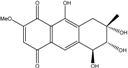 |
--- | Nigrospora sp. | [84] |
| C16H18O7/323 | Dihydroaltersolanol C |
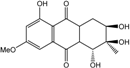 |
(1R,2R,3R)-1,2,3,5-tetra-hydroxy-7-methoxy-2-methyl-1,2,3,4,4a,9a-hexahydroanthraquinone | Stemphylium globuliferum | [94] |
| C16H20O7/324 | Fusaquinon C |
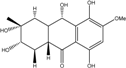 |
(2S,3R,4aR,9aR,10S)-2,3,5,8,10-pentahydroxy-6-methoxy-3-methyl-1,3,4,4a,9a,10-hexahydroanthracen-9(2H)-one | Fusarium sp. No. ZH-210 | [68] |
| C16H21O7/325 | 1-Deoxytetrahydrobostrycin |
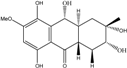 |
(2R,3S)-2,3,5,8,10-pentahydroxy-6-methoxy-3-methyl-1,3,4,4a,9a,10-hexahydroanthracen-9(2H)-one | A. sp. 05F16 Nigrospora sp. | [54] [83] |
| C15H20O8/328 | Fragilin |
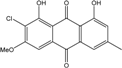 |
2-chloro-1,8-dihydroxy-3-methoxy-6-methylanthraquinone | Letrouitia hafellneri L. leprolytoides | [79][80] |
| C17H12O7/328 | 5-Acetyl-2-methoxy-1,4,6-trihydroxyanthraquinone | 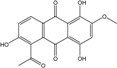 |
---- | Fuarium sp. B77 | [66] |
| C18H16O6/328 | Isorhodoptilometrin-1-methylether | 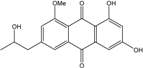 |
1,3-dihydroxy-6-2-hydroxypropyl-8-methoxyanthraquinone | A. versicolor | [57] |
| C17H14O7/329 | 1,3,6,8-Tetrahydroxy-2-(1-methoxyethyl)anthraquinone |
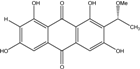 |
--- | Microsphaeropsis | [81] |
| C18H20O6/332 | Phomopsanthraquinone | 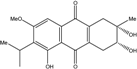 |
(2R,3S)-7-ethyl-1,2,3,4-tetrahydro-2,3,8-trihydroxy-6-methoxy-3-methylanthraquinone | Phomopsis sp. PSU-MA214 | [92] |
| C16H16O8/336 | Altersolanol A | 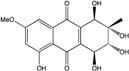 |
(1R,2S,3R,4S)-1,2,3,4,5-pentahydroxy-7-methoxy-2-methyl-1,2,3,4-tetrahydroanthraquinone | Stemphylium globuliferum Xylaria sp. 2508 | [94] [95] |
| C16H16O8/336 | Bostrycin | 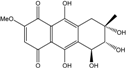 |
(5S,6R,7S)-5,6,7,9,10-pentahydroxy-2-methoxy-7-methyl-5,6,7,8-tetrahydroanthracene-1,4-dione | A. sp. strain 05F16 Al. eichorniae Fusarium spp. PSU-F14/PSU-F135 Halorosellinia sp. No. 1403 Nigrospora sp. Xylaria sp. 2508 | [46][102] [69] [68][86][101][103] [84] [54] [95] |
| C16H18O8/338 | SZ-685C | 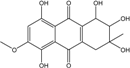 |
1,2,3,5,8-pentahydroxy-6-methoxy-3-methyl-1,2,3,4-tetrahydroanthraquinone | Halorosellinia sp. No. 1403 | [70][101][104][105][106][107] |
| C16H20O8/340 | Fusaquinon B |
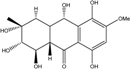 |
(1R,2S,3R,4aR,9aS,10S)-1,2,3,5,8,10-hexahydroxy-6-methoxy-3-methyl-1,3,4,4a,9a,10-hexahydroanthracen-9(2H)-one | Fusarium sp. No. ZH-210 | [68] |
| C16H21O8/340 | Tetrahydroxybostrycin | 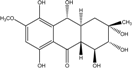 |
1,2,3,5,8,10-hexahydroxy-6-methoxy-3-methyl-1,3,4,4a,9a,10-hexahydroanthracen-9(2H)-one | A. sp. 05F16 Nigrospora sp. MA75 | [54] [85] |
| C18H12O7/340 | Versicolorin C | 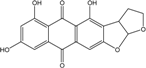 |
4,6,8-trihydroxy-3,3a-dihydroanthra[2,3-b]furo[3,2-d]furan-5,10(2H,12aH)-dione | Fungus ZSUH-36 Fungus Isolate 1850 and isolate 2526 | [65] [64] |
| C18H18O7/346 | 2-O-Acetylaltersolanol B |
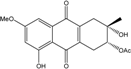 |
(2R,3S)-3,8-dihydroxy-6-methoxy-3-methyl-9,10-dioxo-1,2,3,4,9,10-hexahydroanthracen-2-yl acetate | Stemphylium sp. 33231 | [93] |
| C17H14O8/347 | 1,2,3,6,8-Pentahydroxy-7-(1-methoxyethyl)anthraquinone |
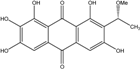 |
1,2,3,6,8-pentahydroxy-7-(1-methoxyethyl)anthracene-9,10-dione | Microsphaeropsis | [81] |
| C15H10O8S/350 | Emodin-3-O Sulfate |
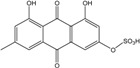 |
4,5-dihydroxy-7-methyl-9,10-dioxo-9,10-dihydroanthracen-2-yl hydrogen sulfate | P. oxalicum 2-HL-M-6 | [91] |
| C16H15O9/351 | Auxarthrol C |
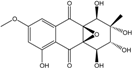 |
(1S,2R,3R,4R,4aR,9aS)-1,2,3,4,5-pentahydroxy-7-methoxy-2-methyl-1,2,3,4-tetrahydro-4a,9a-epoxyanthraquinone | Stemphylium sp. 33231 | [93] |
| C19H11O7/351 | 8-O-MethylversicolorinB |
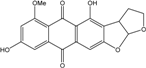 |
4,8-dihydroxy-6-methoxy-3,3a-dihydroanthra[2,3-b]furo[3,2-d]furan-5,10(2H,12aH)-dione | A. versicolor endolichenic | [108] |
| C21H20O5/352 | 6,8-Dimethoxy-1-methyl-2-(3-oxobutyl)anthraquinone | 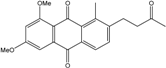 |
---- | Fusarium sp. ZZF60 | [67] |
| C19H13O7/353 | 8-O-Methylversicolorin A |
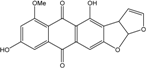 |
4,8-dihydroxy-6-methoxyanthra[2,3-b]furo[3,2-d]furan-5,10(3aH,12aH)-dione | A. versicolor endolichenic | [108] |
| C20H18O6/354 | Averythrin |
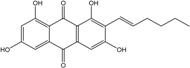 |
(E)-2-(hex-1-en-1-yl)-1,3,6,8-tetrahydroxyanthraquinone | A. sp. SCSIO F063 A. versicolor endolichenic | [55] [108] |
| C30H18O10/358 | Skyrin |
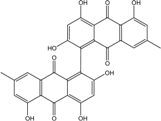 |
2,2′,4,4′,5,5′-hexahydroxy-7,7′-dimethyl-[1,1′-bianthracene]-9,9′,10,10′-tetraone | P. chrysogenum | [89] |
| C16H12O8S/364 | Macrosporin-7-O-sulfate |
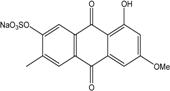 |
Sodium 8-hydroxy-6-methoxy-3-methyl-9,10-dioxo-9,10-dihydroanthracen-2-yl sulfate | Stemphylium sp. 33231 | [93] |
| C15H9O9S/365 | Citreorosein-3-O-sulfate |
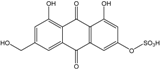 |
4,5-dihydroxy-7-(hydroxymethyl)-9,10-dioxo-9,10-dihydroanthracen-2-yl hydrogen sulfate | P. oxalicum 2-HL-M-6 | [91] |
| C20H14O7/365 | 6,8-di-O-methylversico-lorinA | 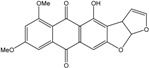 |
4-hydroxy-6,8-dimethoxyanthra[2,3-b]furo[3,2-d]furan-5,10(3aH,12aH)-dione | A. versicolor endolichenic A. versicolor EN-7 (Genbank no EU042148) | [108] [58] |
| C21H19O6/367 | 8-O-Methylaverythrin |
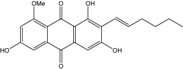 |
(E)-2-(hex-1-en-1-yl)-1,3,6-trihydroxy-8-methoxyanthraquinone | A. versicolor endolichenic | [108] |
| C20H16O7/368 | Aversin | 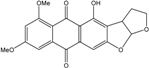 |
4-hydroxy-6,8-dimethoxy-3,3a-dihydroanthra[2,3-b]furo[3,2-d]furan-5,10(2H,12aH)-dione | A. versicolor endolichenic | [108] |
| C20H16O7/368 | Aversin : (−)-isomer | 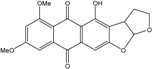 |
4-hydroxy-6,8-dimethoxy-3,3a-dihydroanthra[2,3-b]furo[3,2-d]furan-5,10(2H,12aH)-dione | A. versicolor EN-7 (Genbank no EU042148) | [58] |
| C20H20O7/372 | Averantin | 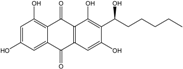 |
(S)-1,3,6,8-tetrahydroxy-2-(1-hydroxyhexyl)anthraquinone | A. sp. SCSIO F063 | [55] |
| C20H20O7/372 | Averantin = (S)-(−)-averantin | 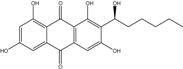 |
(S)-1,3,6,8-tetrahydroxy-2-(1-hydroxyhexyl)anthraquinone | A. sp. SCSIO F063 | [55] |
| C21H18O7/382 | 6-O-Methylaverufin | 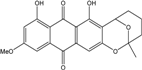 |
7,9-dihydroxy-11-methoxy-2-methyl-3,4,5,6-tetrahydro-2H-2,6-epoxyanthra[2,3-b]oxocine-8,13-dione | Fungus ZSUH-36 A. versicolor EN-7 | [65] [58] |
| C20H16O8/384 | Nidurufin | 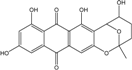 |
5,7,9,11-tetrahydroxy-2-methyl-3,4,5,6-tetrahydro-2H-2,6-epoxyanthra[2,3-b]oxocine-8,13-dione | Fungus Isolate 1850 and isolate 2526 | [64] |
| C20H15O8/386 | Averufin | 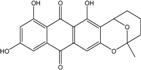 |
7,9,11-trihydroxy-2-methyl-3,4,5,6-tetrahydro-2H-2,6-epoxyanthra[2,3-b]oxocine-8,13-dione | A. versicolor Fungus ZSUH-36 Fungus Isolate 1850 and isolate 2526 | [109] [65] [64] |
| C21H22O7/386 | 1′-O-Methylaverantin |
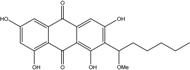 |
1,3,6,8-tetrahydroxy-2-(1-methoxyhexyl)anthraquinone | A. sp. SCSIO F063 Fungus ZSUH-36 | [55] [65] |
| C19H15O9/388 | (2S)-2,3-Dihydroxy-propyl-1,6,8-trihydroxy-3-methyl-9,10-dioxoanthracene-2-carboxylate | 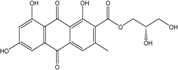 |
(1S,5′S,6′R,7′S,8′R)-1′,2,5′,6′,7′,8,8′-heptahydroxy-3′,6-dimethoxy-3,6′-dimethyl-5′,6′,7′,8′,8′a,10′a-hexahydro-[1,2′-bianthracene]-9,9′,10,10′-tetraone | A. variecolor B-17 | [56] |
| C20H17ClO6/388 | 7-Chloroaverythrin | 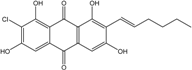 |
(E)-2-chloro-7-(hex-1-en-1-yl)-1,3,6,8-tetrahydroxyanthraquinone | A. sp. SCSIO F063 | [55] |
| C20H20O8/388 | 6,8-Di-O-methylversiconol | 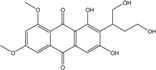 |
2-(1,4-dihydroxybutan-2-yl)-1,3-dihydroxy-6,8-dimethoxyanthraquinone | A. versicolor EN-7 (Genbank no EU042148) | [58] |
| C22H20O7/396 | 6,8-Di-O-methylaverufin |
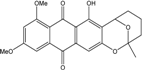 |
7-hydroxy-9,11-dimetho-xy-2-methyl-3,4,5,6-tetra-hydro-2H-2,6-epoxyanthra[2,3-b]oxocine-8,13-dione | A. versicolor endolichenic | [108] |
| C22H20O7/396 | 6,8-Di-O-methylnidurufin | 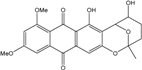 |
5,7-dihydroxy-9,11-dime-thoxy-2-methyl-3,4,5,6-tetrahydro-2H-2,6-epoxyanthra[2,3-b]oxocine-8,13-dione | A. versicolor endolichenic A. versicolor EN-7 (Genbank no EU042148) | [108] [58] |
| C22H22O7/398 | 6,8-Di-O-methylaverufanin |
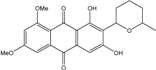 |
1,3-dihydroxy-6,8-dimethoxy-2-(6-methyltetra-hydro-2H-pyran-2-yl)anthracene-9,10-dione | Fungus ZSUH-36 | [65] |
| C22H24O7/400 | 6,1′-O,O-Dimethylaverantin |
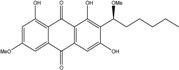 |
(S)-1,3,8-trihydroxy-6-methoxy-2-(1-methoxyhexyl)anthraquinone | A. sp. SCSIO F063 | [55] |
| C20H17O9/401 | Variecolorquinone A |
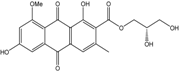 |
(S)-2,3-dihydroxypropyl 1,6-dihydroxy-8-methoxy-3-methyl-9,10-dioxo-9,10-dihydroanthracene-2-carboxylate | A.glaucus A. variecolor B-17 | [50][51][52][53] [56] |
| C22H24O7/401 | 6,8-Di-O-methylaveran-tin |
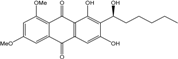 |
(S)-1,3-dihydroxy-2-(1-hydroxyhexyl)-6,8-dimetho-xyanthraquinone | A. versicolor EN-7 (Genbank no EU042148) A. sp. SCSIO F063 | [58] [55] |
| C21H19ClO6/402 | 6-O-Methyl-7-chloroaverythrin |
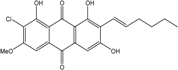 |
(E)-2-chloro-7-(hex-1-en-1-yl)-1,6,8-trihydroxy-3-methoxyanthraquinone | A. sp. SCSIO F063 | [55] |
| C20H19ClO7/407 | (1′S)-7-Chloroaverantin |
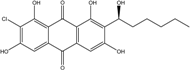 |
(S)-2-chloro-1,3,6,8-tetrahydroxy-7-(1-hydroxyhexyl)anthraquinone | A. sp. SCSIO F063 | [55] |
| C22H23ClO7/407 | 6,1′-O,O-Dimethyl-7-chloroaverantin |
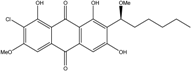 |
(S)-2-chloro-1,6,8-trihydroxy-3-methoxy-7-(1-methoxyhexyl)anthraquinone | A. sp. SCSIO F063 | [55] |
| C23H26O7/414 | 6,8,1′-Tri-O-methyl-averantin |
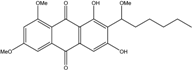 |
1,3-dihydroxy-6,8-dimethoxy-2-(1-methoxyhexyl)anthraquinone | A.versicolor endolichenic Fungus ZSUH-36 | [108] [65] |
| C21H19O9/415 | 6-3-O-(Ribofuranosyl)questin | 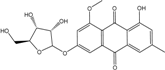 |
1,6-dihydroxy-6-O-(ribofuranosyl)-8-methoxy-3-methylanthraquinone | Eurotium rubrum | [63] |
| C21H21ClO7/420 | 1′-O-methyl-7-chloro averantin |
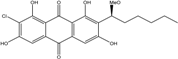 |
(S)-2-chloro-1,3,6,8-tetrahydroxy-7-(1-methoxy-hexyl)anthraquinone | A. sp. SCSIO F063 | [55] |
| C21H21ClO7/421 | 6-O-methyl-7-chloro-averantin |
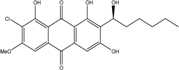 |
(S)-2-chloro-1,6,8-trihydroxy-7-(1-hydroxyhexyl)-3-methoxyanthraquinone | A. sp. SCSIO F063 | [55] |
| C24H28O8/444 | Averantin-1′-butyl ether | 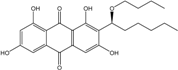 |
(S)-2-(1-butoxyhexyl)-1,3,6,8-tetrahydroxy-anthraquinone | A. sp. SCSIO F063 | [55] |
| C24H27ClO7/463 | 7-Chloroaverantin-1′-butyl ether |
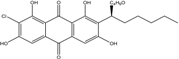 |
(S)-2-(1-butoxyhexyl)-7-chloro-1,3,6,8-tetrahy-droxyanthraquinone | A. sp. SCSIO F063 | [55] |
| C21H21BrO7/465 | 6-O-Methyl-7-bromoaverantin |
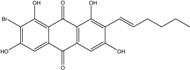 |
(S)-2-bromo-1,6,8-trihydroxy-7-(1-hydroxyhexyl)-3-methoxyanthraquinone | A. sp. SCSIO F063 | [55] |
| C22H23BrO7/479 | 6,1′-O,O-Dimethyl-7-bromoaverantin |
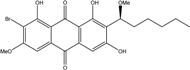 |
(S)-2-bromo-1,6,8-trihydroxy-3-methoxy-7-(1-methoxyhexyl)anthraquinone | A. sp. SCSIO F063 | [55] |
| C24H23O11/487 | Macrosporin2-O-(6′-acetyl)-a-d-glucopyranoside |
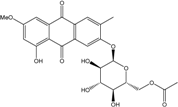 |
((2R,3S,4S,5R,6R)-3,4,5-trihydroxy-6-((8-hydroxy-6-methoxy-3-methyl-9,10-dioxo-9,10-dihydroanthracen-2-yl)oxy)tetrahydro-2H-pyran-2-yl)methyl acetate | Stemphylium sp. 33231 | [93] |
| C28H24O10/520 | Penicillanthranin A |
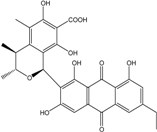 |
(1S,3R,4S)-1-(6-ethyl-1,3,8-trihydroxy-9,10-dioxo-9,10-dihydroanthracen-2-yl)-6,8-dihy-droxy-3,4,5-trimethyliso-chroman-7-carboxylic acid | P. citrinum PSU-F51 | [88] |
| C28H24O11/536 | Penicillanthranin B |
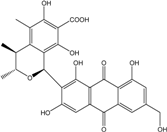 |
(1S,3R,4S)-6,8-dihydroxy-3,4,5-trimethyl-1-(1,3,8-trihydroxy-6-(hydroxy-methyl)-9,10-dioxo-9,10-dihydroanthracen-2-yl)isochroman-7-carboxylic acid | P. citrinum PSU-F51 | [88] |
| C32H24O8/536 | (trans)-R (cis)-Emodin-Physcion bianthrone | 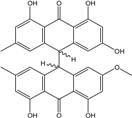 |
2,4,4′,5,5′-pentahydroxy-2′-methoxy-7,7′-dimethyl-[9,9′-bianthracene]-10,10′(9H,9′H)-dione | A. glaucus | [50][51][52][53] |
| C32H21O10/565 | Alterporriol Q |
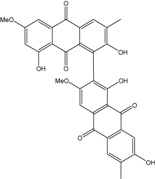 |
1′,2,7′,8-tetrahydroxy-3′,6-dimethoxy-3,6′-dimethyl-[1,2′-bianthracene]-9,9′,10,10′-tetraone | Al. sp. ZJ-2008003 | [48] |
| C32H21O10/565 | Alterporriol R |
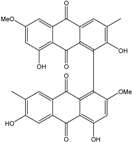 |
2,4′,6′,8-tetrahydroxy-2′,6-dimethoxy-3,7′-dimethyl-[1,1′-bianthracene]-9,9′,10,10′-tetraone | Al. sp. ZJ-2008003 | [48] |
| C32H22O10/566 | Alterporriol V |
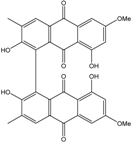 |
2,2′,8,8′-tetrahydroxy-6,6′-dimethoxy-3,3′-dimethyl-[1,1′-bianthracene]-9,9′,10,10′-tetraone | Stemphylium sp. 33231 | [93] |
| C30H22O12/574 | Cytoskyrin A | 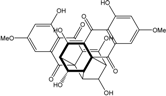 |
(6R,14R,17S,18R,19R,20S)-1,7,9,15,17,20-hexahydroxy-3,11-dimethoxy-6,13a,5a,14-(epibutane[1][2][3][4]tetrayl)cycloocta[1,2-b:5,6-b′]dinaphtha-lene-5,8,13,16(6H,14H)-tetraone | Curvularia lunata | [50][51][59][60] |
| C32H26O10/586 | Alterporriol K |
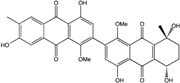 |
(5S,8R)-4,4′,5,7′,8-penta-hydroxy-1,1′-dimethoxy-6′,8-dimethyl-5,6,7,8-tetrahydro-[2,2′-bianthracene]-9,9′,10,10′-tetraone | Al. sp. ZJ9-6B Al. sp. ZJ-2008003 | [49] [48] |
| C32H30O13/590 | Alterporriol T |
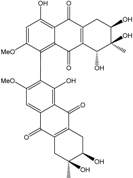 |
(6R,6′S,7R,7′R,8R)-1′,4,6,6′,7,7′,8-heptahydroxy-2,3′-dimethoxy-6′,7-dime-thyl-5,5′,6,6′,7,7′,8,8′-octahydro-[1,2′-bianthracene]-9,9′,10,10′-tetraone | Stemphylium sp. 33231 | [93] |
| C32H25O12/601 | Alterporriol L |
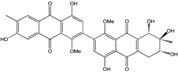 |
(6S,7R,8R)-4,4′,6,7,7′,8-hexahydroxy-1,1′-dimethoxy-6′,7-dimethyl-5,6,7,8-tetrahydro-[2,2′-bianthracene]-9,9′,10,10′-tetraone | Al. sp. ZJ9-6B Al. sp. ZJ-2008003 | [49] [48] |
| C32H25O12/601 | Alterporriol M |
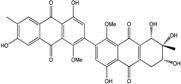 |
(6S,7S,8R)-4,4′,6,7,7′,8-hexahydroxy-1,1′-dimethoxy-6′,7-dimethyl-5,6,7,8-tetrahydro-[2,2′-bianthracene]-9,9′,10,10′-tetraone | Al. sp. ZJ9-6B Al. sp. ZJ-2008003 | [49] [48] |
| C32H25O12/601 | Alterporriol P |
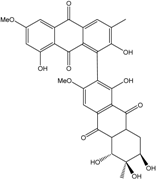 |
(5′R,6′R,7′R)-1′,2,5′,6′,7′,8-hexahydroxy-3′,6-dimethoxy-3,6′-dimethyl-5′,6′,7′,8′,8′a,10′a-hexahydro-[1,2′-bianthracene]-9,9′,10,10′-tetraone | Al. sp. ZJ-2008003 | [48] |
| C32H26O12/602 | Alterporriol W |
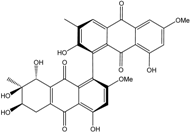 |
(1′R,6R,7R,8R)-2′,4,6,7,8,8′-hexahydroxy-2,6′-dimethoxy-3′,7-dimethyl-5,6,7,8-tetrahydro-[1,1′-bianthracene]-9,9′,10,10′-tetraone | Stemphylium sp. 33231 | [93] |
| C32H30O12/606 | Alterporriol U |
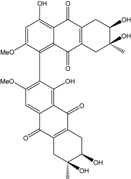 |
(6R,6′S,7S,7′R)-1′,4,6,6′,7,7′-hexahydroxy-2,3′-di-methoxy-6′,7-dimethyl-5,5′,6,6′,7,7′,8,8′-octahy-dro-[1,2′-bianthracene]-9,9′,10,10′-tetraone | Stemphylium sp. 33231 | [93] |
| C31H32O13/612 | Alterporriol S |
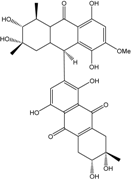 |
(2′S,3′R,4′S,6R,7S,9′R)-1,2′,3′,4,5′,6,7,8′-octahy-droxy-7′-methoxy-2′,4′,7-trimethyl-2′,3′,4′,4′a,5,6,7,8,9′,9′a-decahydro-[2,9′-bianthracene]-9,10,10′(1′H)-trione | Al. sp. (SK11) | [47] |
| C32H25O13/617 | (+)-aS-alterporriol C |
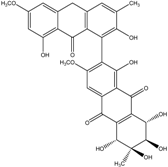 |
(1S,5′S,6′R,7′S,8′R)-1′,2,5′,6′,7′,8,8′-heptahydroxy-3′,6-dimethoxy-3,6′-dime-thyl-5′,6′,7′,8′,8′a,10′a-hexahydro-[1,2′-bianthracene]-9,9′,10,10′-tetraone | Al. sp. (SK11) | [47] |
| C32H25O13/617 | Alterporriol C |
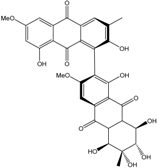 |
(1S,5′R,6′S,7′R,8′S)-1′,2,5′,6′,7′,8,8′-heptahydroxy-3′,6-dimethoxy-3,6′-dime-thyl-5′,6′,7′,8′,8′a,10′a-hexahydro-[1,2′-bianthracene]-9,9′,10,10′-tetraone | Al. sp. (SK11) Al. sp. ZJ-2008003 | [47] [48] |
| C32H29O14/637 | Alterporriol N |
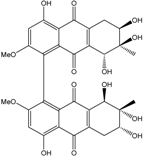 |
(6R,6′R,7R,7′R,8R,8′R)-4,4′,6,6′,7,7′,8,8′-octahy-droxy-2,2′-dimethoxy-7,7′-dimethyl-5,5′,6,6′,7,7′,8,8′-octahydro-[1,1′-bianthracene]-9,9′,10,10′-tetraone | Al. sp. ZJ-2008003 | [48] |
| C32H29O14/637 | Alterporriol O |
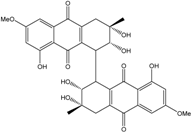 |
(2R,2′R,3S,3′S)-2,2′,3,3′,8,8′-hexahydroxy-6,6′-dimethoxy-3,3′-dimethyl-1,1′,2,2′,3,3′,4,4′-octahy-dro-[1,1′-bianthracene]-9,9′,10,10′-tetraone | Al. sp. ZJ-2008003 | [48] |
| C34H33O17/713 | Acetylalterporriol D |
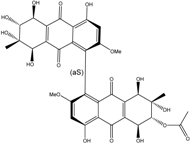 |
(1′S,5S,5′S,6R,6′R,7S,7′S,8R,8′R)-4,4′,5,5′,6′,7,7′,8,8′-nonahydroxy-2,2′-dimethoxy-7,7′-dimethyl-9,9′,10,10′-tetraoxo-5,5′,6,6′,7,7′,8,8′,9,9′,10,10′-dodecahydro-[1,1′-bianthracen]-6-yl acetate | Stemphylium globuliferum | [94] |
| C34H33O17/713 | Acetylalterporriol E |
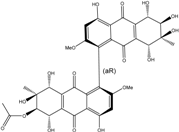 |
(1′R,5S,5′S,6R,6′R,7S,7′S,8R,8′R)-4,4′,5,5′,6′,7,7′,8,8′-nonahydroxy-2,2′-dimethoxy-7,7′-dimethyl-9,9′,10,10′-tetraoxo-5,5′,6,6′,7,7′,8,8′,9,9′,10,10′-dodecahydro-[1,1′-bianthracen]-6-yl acetate | Stemphylium globuliferum | [94] |
| C34H33O17/713 | Alterporriol D |
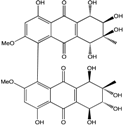 |
(1S,5S,5′S,6R,6′R,7S,7′S,8R,8′R)-4,4′,5,5′,6,6′,7,7′,8,8′-decahydroxy-2,2′-dimethoxy-7,7′-dimethyl-5,5′,6,6′,7,7′,8,8′-octahy-dro-[1,1′-bianthracene]-9,9′,10,10′-tetraone | Stemphylium globuliferum | [94] |
| C34H33O17/713 | Alterporriol E |
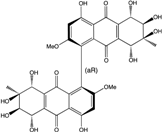 |
(1R,5S,5′S,6R,6′R,7S,7′S,8R,8′R)-4,4′,5,5′,6,6′,7,7′,8,8′-decahydroxy-2,2′-dimethoxy-7,7′-dimethyl-5,5′,6,6′,7,7′,8,8′-octahy-dro-[1,1′-bianthracene]-9,9′,10,10′-tetraone | Stemphylium globuliferum | [94] |
| C48H40O21/952 | Stemphylanthranol A | 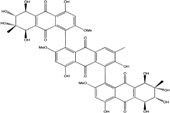 |
(5S,5′′S,6R,6′′R,7S,7′′S,8R,8′′R)-2′,4,4′′,5,5′′,6,6′′,7,7′′,8,8′,8′′-dodecahydroxy-2,2′′,6′-trimethoxy-3′,7,7′′-trimethyl-5,5′′,6,6′′,7,7′′,8,8′′-octahydro-[1,1′:5′,1′′-teranthracene]-9,9′,9′′,10,10′,10′′-hexaone | Stemphylium globuliferum | [94] |
| C48H40O21/952 | Stemphylanthranol B | 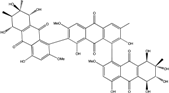 |
(5S,5′′R,6R,6′′R,7S,7′′R,8R,8′′R)-2′,4,4′′,5,5′′,6,7,7′′,8,8′,8′′-undecahydroxy-2,2′′,6′-trimethoxy-3′,6′′,7,7′′-tetramethyl-5,5′′,6,6′′,7,7′′,8,8′′-octahydro-[1,1′:7′,1′′-tetranthracene]-9,9′,9′′,10,10′,10′′-hexaone | Stemphylium globuliferum | [94] |
| ---- | 7-Chloroemodic acid | ---- | ---- | Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) | [75][76][77][79][96] |
| ---- | 7-Chloroemodinal | ---- | ---- | Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.)L. hafellneri L. leprolytoides | [75][76][77][79][96] [80] |
| ---- | 7-Chloro-1,6,8-trihydroxy-3-methyl-10-anthrone | ---- | ---- | Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) | [75][76][77][79][96] |
 ; Orange:
; Orange:  ; Yellow:
; Yellow:  ; Red:
; Red:  ; Bronze:
; Bronze:  .
.
Ubiquitous Fungi
Endophytes and/or Pathogens
Lichens
3. Biosynthesis and Known Roles for Anthraquinones in Fungi
3.1. Biosynthetic Route and Genes Involved
3.2. Roles in the Biology of Fungi
References
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2012, 29, 144–222.
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2015, 32, 116–211.
- Montaser, R.; Luesch, H. Marine natural products: A new wave of drugs? Future Med. Chem. 2011, 3, 1475–1489.
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69–85.
- Bhatnagar, I.; Kim, S.-K. Marine antitumor drugs: Status, shortfalls and strategies. Mar. Drugs 2010, 8, 2702–2720.
- Bugni, T.S.; Ireland, C.M. Marine diversity. Nat. Prod. Rep. 2004, 21, 143–163.
- Damare, S. Deep-Sea Fungi: Occurrence and Adaptations. Ph.D Thesis, Goa University, Goa, India, 2006.
- Kohlmeyer, J.; Kohlmeyer, E. Marine Mycology; Elsvier: London, UK, 1979; p. 704.
- Blackwell, M. The Fungi: 1, 2, 3 … 5.1 million species? Am. J. Bot. 2011, 98, 426–438.
- Jones, E.B. G. Des champignons dans l'océan ! Biofutur 1998, 1998, 18–20.
- Yu, Z.; Lang, G.; Kajahn, I.; Schmaljohann, R.; Imhoff, J.F. Scopularides A and B, cyclodepsipeptides from a marine sponge-derived fungus, Scopulariopsis brevicaulis. J. Nat. Prod. 2008, 71, 1052–1054.
- Imhoff, J.; Yu, Z.; Lang, G.; Wiese, J.; Kalthoff, H.; Klose, S. Production and Use of Antitumoral Cyclodepsipeptides. U.S. Patent 8,765,907 B2, 11 June 2009.
- Newman, D.; Cragg, G. Marine-sourced anti-cancer and cancer pain control agents in clinical and late preclinical development. Mar. Drugs 2014, 12, 255.
- Ebel, R. Natural product diversity from marine fungi. In Comprehensive Natural Products II: Chemistry and Biology; Mander, L., Liu, H.-W., Eds.; Elsevier: Oxford, UK, 2010; Volume 2, pp. 223–262.
- Hanson, J.R. Natural Products: The Secondary Metabolites; Royal Society of Chemistry: Cambridge, UK, 2003; p. 147.
- Caro, Y.; Anamale, L.; Fouillaud, M.; Laurent, P.; Petit, T.; Dufosse, L. Natural hydroxyanthraquinoid pigments as potent food grade colorants: An overview. Nat. Prod. Bioprospect. 2012, 2, 174–193.
- Gessler, N.N.; Egorova, A.S.; Belozerskaia, T.A. Fungal anthraquinones (review). Appl. Biochem. Microbiol. 2013, 49, 85–99.
- Anslow, W.K.; Raistrick, H. Studies in the biochemistry of micro-organisms: Synthesis of cynodontin (1:4:5:8-tetrahydroxy-2-methylanthraquinone), a metabolic product of species of Helminthosporium. Biochem. J. 1940, 34, 1546–1548.
- Taylor, G.W. Natural dyes in textile applications. Rev. Progress Color. Relat. Top. 1986, 16, 53–61.
- Shore, J. Colorants and Auxiliaries; Society of Dyers and Colourists: Bradford, UK, 1990; Volume 1, p. 372.
- Stoessl, A. Some metabolites of Alternaria solani. Can. J. Chem. 1969, 47, 767–776.
- Kühlwein, F.; Polborn, K.; Beck, W. Metallkomplexe von farbstoffen. VIII übergangsmetallkomplexe des curcumins und seiner derivate. Z. Anorg. Allg. Chem. 1997, 623, 1211–1219.
- Yuzhen, Z.; Wenfu, H. The influence of methyl groups on the colour and dyeing properties of acid dyes derived from 1-amino-4-bromo-anthraquinone-2-sulphonic acid and arylamines. Dyes Pigm. 1996, 30, 283–289.
- Yuzhen, Z.; Wenfu, H.; Yueping, T. Structure and dyeing properties of some anthraquinone violet acid dyes. Dyes Pigments 1997, 34, 25–35.
- Kim, S.-K. Marine Microbiology: Bioactive Compounds and Biotechnological Applications; Wiley & Sons: Weinheim, Germany, 2013; p. 550.
- Kjer, J.; Debbab, A.; Aly, A.H.; Proksch, P. Methods for isolation of marine-derived endophytic fungi and their bioactive secondary products. Nat. Protoc. 2010, 5, 479–490.
- Dufossé, L. Anthraquinones, the Dr Jekyll and Mr Hyde of the food pigment family. Food Res. Int. 2014, 65, 132–136.
- Fain, V.Y.; Zaitsev, B.E.; Ryabov, M.A. Metal complexes with 1,5- and 1,8-Dihydroxy-9,10-anthraquinones: Electronic absorption spectra and structure of ligands. Russ. J. Coord. Chem. 2004, 30, 360–364.
- Fain, V.Y.; Zaitsev, B.E.; Ryabov, M.A. Tautomerism of the natural 1,8-dihydroxy-9,10-anthraquinones chrysophanol, aloe-emodin, and rhein. Chem. Nat. Compd. 2005, 41, 146–152.
- Thomson, R.H. Anthraquinones. In Naturally Occurring Quinones; Academic Press: London, UK, 1971; pp. 367–535.
- Panno, L.; Bruno, M.; Voyron, S.; Anastasi, A.; Gnavi, G.; Miserere, L.; Varese, G.C. Diversity, ecological role and potential biotechnological applications of marine fungi associated to the seagrass Posidonia oceanica. New Biotechnol. 2013, 30, 685–694.
- Mtui, G.; Nakamura, Y. Lignocellulosic enzymes from Flavodon flavus, a fungus isolated from Western Indian Ocean off the coast of Dar es Salaam, Tanzania. Afr. J. Biotechnol. 2008, 7, 3066–3072.
- Singh, P.; Raghukumar, C.; Meena, R.M.; Verma, P.; Shouche, Y. Fungal diversity in deep-sea sediments revealed by culture-dependent and culture-independent approaches. Fungal Ecol. 2012, 5, 543–553.
- Raghukumar, C. Marine fungal biotechnology: An ecological perspective. Fungal Divers. 2008, 31, 19–35.
- Kolesnikov, M.P.; Mirchink, T.G.; Lur’ ye, N.Y. Pigments of Penicillium funiculosum strains isolated from various types of soils. Sov. Soil Sci. 1984, 16, 28–34.
- Anke, H.; Kolthoum, I.; Zähner, H.; Laatsch, H. Metabolic products of microorganisms. 185. The anthraquinones of the Aspergillus glaucus group. I. Occurrence, isolation, identification and antimicrobial activity. Arch. Microbiol. 1980, 126, 223–230.
- Saleem, M.; Nazir, M. Bioactive natural products from marine-derived fungi: An update. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, Netherlands, 2015; Volume 45, pp. 297–361.
- Kathiresan, K.; Bingham, B.L. Biology of mangroves and mangrove ecosystems. In Advances in Marine Biology; Southward, A., Young, C., Fuiman, L., Tyler, P., Eds.; Academic Press: London, UK, 2001; Volume 40, pp. 81–251.
- Hawksworth, D.L. Hypocrealean fungi revisited. Mycol. Res. 2000, 104, 117–118.
- Bick, I.R.C.; Rhee, C. Anthraquinone pigments from Phoma foveata Foister. Biochem. J. 1966, 98, 112–116.
- Fain, V.Y.; Zaitsev, B.E.; Ryabov, M.A. Proportional response method for the assignment of π1, π* adsorption bands of ionized hydroxyanthraquinones. Žurnal fizičeskoj himii 2003, 77, 563–565.
- Fujitake, N.; Suzuki, T.; Fukumoto, M.; Oji, Y. Predomination of dimers over naturally occurring anthraquinones in soil. J. Nat. Prod. 1998, 61, 189–192.
- Fujitake, N.; Azuma, J.; Hamasaki, T.; Nakajima, H.; Saiki, K. Chrysotalunin, a most prominent soil anthraquinone pigment in Japanese soils. Sci. Total Environ. 1992, 117–118, 219–226.
- Suzuki, T.; Fujitake, N.; Oji, Y. Horizontal Distribution of main hydroxyanthraquinones in soil. Soil Sci. Plant Nutr. (Tokyo Jpn.) 1999, 45, 297–306.
- Velmurugan, P. Studies on the Production and Dyeing Properties of Water Soluble Pigments from Filamentous Fungi. Ph.D. Thesis, Bharathiar University, Coimbatore, India, 2008.
- Charudattan, R.; Rao, K.V. Bostrycin and 4-deoxybostrycin: Two nonspecific phytotoxins produced by Alternaria eichhorniae. Appl. Environ. Microbiol. 1982, 43, 846–849.
- Xia, G.; Li, J.; Li, H.; Long, Y.; Lin, S.; Lu, Y.; He, L.; Lin, Y.; Liu, L.; She, Z. Alterporriol-type dimers from the mangrove endophytic fungus, Alternaria sp. (SK11), and their MptpB inhibitions. Mar. Drugs 2014, 12, 2953–2969.
- Zheng, C.J.; Shao, C.L.; Guo, Z.Y.; Chen, J.F.; Deng, D.S.; Yang, K.L.; Chen, Y.Y.; Fu, X.M.; She, Z.G.; Lin, Y.C.; et al. Bioactive hydroanthraquinones and anthraquinone dimers from a soft coral-derived Alternaria sp. fungus. J. Nat. Prod. 2012, 75, 189–197.
- Huang, C.H.; Pan, J.H.; Chen, B.; Yu, M.; Huang, H.B.; Zhu, X.; Lu, Y.J.; She, Z.G.; Lin, Y.C. Three bianthraquinone derivatives from the mangrove endophytic fungus Alternaria sp. ZJ9–6B from the South China Sea. Mar. Drugs 2011, 9, 832–843.
- Duran, N.; Teixeira, M.F.S.; de Conti, R.; Esposito, E. Ecological-friendly pigments from fungi. Crit. Rev. Food Sci. Nutr. 2002, 42, 53–66.
- Mapari, S.A.S.; Nielsen, K.F.; Larsen, T.O.; Frisvad, J.C.; Meyer, A.S.; Thrane, U. Exploring fungal biodiversity for the production of water-soluble pigments as potential natural food colorants. Curr. Opin. Biotechnol. 2005, 16, 231–238.
- Frisvad, J.C.; Filtenborg, O. Terverticillate Penicillia: Chemotaxonomy and mycotoxin production. Mycologia 1989, 81, 837–861.
- Du, L.; Zhu, T.; Liu, H.; Fang, Y.; Zhu, W.; Gu, Q. Cytotoxic Polyketides from a Marine-derived Fungus Aspergillus glaucus. J. Nat. Prod. 2008, 71, 1837–1842.
- Xu, J.; Nakazawa, T.; Ukai, K.; Kobayashi, H.; Mangindaan, R.E.P.; Wewengkang, D.S.; Rotinsulu, H.; Namikoshi, M. Tetrahydrobostrycin and 1-deoxytetrahydrobostrycin, two new hexahydroanthrone derivatives, from a marine-derived fungus Aspergillus sp. J. Antibiot. 2008, 61, 415–419.
- Huang, H.; Wang, F.; Luo, M.; Chen, Y.; Song, Y.; Zhang, W.; Zhang, S.; Ju, J. Halogenated anthraquinones from the marine-derived fungus Aspergillus sp. SCSIO F063. J. Nat. Prod. 2012, 75, 1346–1352.
- Wang, W.; Zhu, T.; Tao, H.; Lu, Z.; Fang, Y.; Gu, Q.; Zhu, W. Two new cytotoxic quinone type compounds from the halotolerant fungus Aspergillus variecolor. J. Antibiot. 2007, 60, 603–607.
- Hawas, U.W.; El-Beih, A.A.; El-Halawany, A.M. Bioactive anthraquinones from endophytic fungus Aspergillus versicolor isolated from red sea algae. Arch. Pharm. Res. 2012, 35, 1749–1756.
- Zhang, Y.; Li, X.M.; Wang, B.G. Anthraquinone derivatives produced by Marine-Derived Fungus Aspergillus versicolor EN-7. Biosci. Biotechnol. Biochem. 2012, 76, 1774–1776.
- Räisänen, R. Dyes from lichens and mushrooms. In Handbook of Natural Colorants; Bechtold, T., Mussak, R.A.M., Eds.; John Wiley & Sons: Chippenham, Wiltshire, UK, 2009; pp. 183–200.
- Jadulco, R.; Brauers, G.; Edrada, R.A.; Ebel, R.; Wray, V.; Sudarsono, S.; Proksch, P. New metabolites from sponge-derived fungi Curvularia lunata and Cladosporium herbarum. J. Nat. Prod. 2002, 65, 730–733.
- Almeida, A.P.; Dethoup, T.; Singburaudom, N.; Lima, R.; Vasconcelos, M.H.; Pinto, M.; Kijjoa, A. The in vitro anticancer activity of the crude extract of the sponge-associated fungus Eurotium cristatum and its secondary metabolites. J. Nat. Pharm. 2010, 1, 25.
- Smetanina, O.F.; Kalinovskii, A.I.; Khudyakova, Y.V.; Slinkina, N.N.; Pivkin, M.V.; Kuznetsova, T.A. Metabolites from the marine fungus Eurotium repens. Chem. Nat. Compd. 2007, 43, 395–398.
- Li, D.; Li, X.; Wang, B. Natural anthraquinone derivatives from a marine mangrove plant-derived endophytic fungus Eurotium rubrum: Structural elucidation and DPPH radical scavenging activity. J. Microbiol. Biotechnol. 2009, 19, 675–680.
- Zhu, F.; Chen, G.; Chen, X.; Yuan, Y.; Huang, M.; Xiang, W.; Sun, H. Structural elucidation of three anthraquinones from a marine-derived mangrove endophytic fungus (isolate 1850). In Proceeding of the International Conference on BioMedical Engineering and Informatics, Sanya, China, 27–30 May 2008; pp. 664–667.
- Shao, C.; Wang, C.; Wei, M.; Li, S.; She, Z.; Gu, Y.; Lin, Y. Structural and spectral assignments of six anthraquinone derivatives from the mangrove fungus (ZSUH-36). Magn. Reson. Chem. 2008, 46, 886–889.
- Shao, C.L.; Wang, C.Y.; Zheng, C.J.; She, Z.G.; Gu, Y.C.; Lin, Y.C. A new anthraquinone derivative from the marine endophytic fungus Fusarium sp. (No. b77). Nat. Prod. Res. 2010, 24, 81–85.
- Huang, Z.-J.; Yang, R.-Y.; Guo, Z.-Y.; She, Z.-G.; Lin, Y.-C. New anthraquinone derivative produced by cultivation of mangrove endophytic fungus Fusarium sp. ZZF60 from the South China Sea. Chin. J. Appl. Chem. 2010, 27, 394–397.
- Chen, Y.; Cai, X.; Pan, J.; Gao, J.; Li, J.; Yuan, J.; Fu, L.; She, Z.; Lin, Y. Structure elucidation and NMR assignments for three anthraquinone derivatives from the marine fungus Fusarium sp. (No. ZH-210). Magn. Reson. Chem. 2009, 47, 362–365.
- Trisuwan, K.; Khamthong, N.; Rukachaisirikul, V.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Anthraquinone, cyclopentanone, and naphthoquinone derivatives from the sea fan-derived fungi Fusarium spp. PSU-F14 and PSU-F135. J. Nat. Prod. 2010, 1507–1511.
- Xia, X.K.; Huang, H.R.; She, Z.G.; Shao, C.L.; Liu, F.; Cai, X.L.; Vrijmoed, L.L.P.; Lin, Y.C. 1H and 13C NMR assignments for five anthraquinones from the mangrove endophytic fungus Halorosellinia sp. (No. 1403). Magn. Reson. Chem. 2007, 45, 1006–1009.
- Xiang, Z.S.; Zhou, W.Q.; Cai, M.H.; Zhang, Y. Method for Fermental Cultivation of Marine Fungi for Production of Anticancer Compound 1403 C. International Patent Number CN 104250657 A, 31 December 2014.
- Santesson, J. Anthraquinones in Caloplaca. Phytochemistry 1970, 9, 2149–2166.
- Manojlovic, N.; Novakovic, M.; Stevovic, V.; Solujic, S. Antimicrobial metabolites from three Serbian Caloplaca. Pharm. Biol. 2005, 43, 718–722.
- Manojlović, N.; Marković, Z.; Gritsanapan, W.; Boonpragob, K. High-performance liquid chromatographic analysis of anthraquinone compounds in the Laurera benguelensis. Russ. J. Phys. Chem. 2009, 83, 1554–1557.
- Manojlovic, N.T.; Vasiljevic, P.J.; Markovic, Z.S. Antimicrobial activity of extracts and various fractions of chloroform extract from the lichen Laurera benguelensis. J. Biol. Res. Thessalon. 2002, 13, 27–34.
- Søchting, U.; Figueras, G. Caloplaca lenae sp. nov., and other Caloplaca species with caloploicin and vicanicin. Lichenologist 2007, 39, 7–14.
- Yosioka, I.; Hino, K.; Fujio, M.; Kitagawa, I. The structure of caloploicin, a new lichen trichloro-depsidone. Chem. Pharm. Bull. 1973, 21, 1547–1553.
- Ren, H.; Tian, L.; Gu, Q.; Zhu, W. Secalonic acid D; A cytotoxic constituent from marine lichen-derived fungus Gliocladium sp. T31. Arch. Pharm. Res. 2006, 29, 59–63.
- Elix, J. Lichen phytochemistry: Additions and amendments I John A. Elix. Australasian Lichenol. 2008, 63, 20–25.
- Johansson, S.; Søchting, U.; Elix, J.; Wardlaw, J. Chemical variation in the lichen genus Letrouitia (Ascomycota, Letrouitiaceae). Mycol. Prog. 2005, 4, 139–148.
- Brauers, G.; Edrada, R.A.; Ebel, R.; Proksch, P.; Wray, V.; Berg, A.; Grafe, U.; Schachtele, C.; Totzke, F.; Finkenzeller, G.; et al. Anthraquinones and betaenone derivatives from the sponge-associated fungus Microsphaeropsis species: Novel inhibitors of protein kinases. J. Nat. Prod. 2000, 63, 739–745.
- El-Beih, A.A.; Kawabata, T.; Koimaru, K.; Ohta, T.; Tsukamoto, S. Monodictyquinone A: A new antimicrobial anthraquinone from a sea urchin-derived fungus Monodictys sp. Chem. Pharm. Bull. 2007, 55, 1097–1098.
- Kjer, J. New Natural Products from Endophytic Fungi from Mangrove Plants: Structure Elucidation and Biological Screening; Heinrich-Heine-Universität: Düsseldorf, Germany, 2009.
- Yang, K.L.; Wei, M.Y.; Shao, C.L.; Fu, X.M.; Guo, Z.Y.; Xu, R.F.; Zheng, C.J.; She, Z.G.; Lin, Y.C.; Wang, C.Y. Antibacterial anthraquinone derivatives from a sea anemone-derived fungus Nigrospora sp. J. Nat. Prod. 2012, 75, 935–941.
- Deshmukh, S.K.; Verekar, S.A.; Bhave, S.V. Endophytic fungi: A reservoir of antibacterials. Front. Microbiol. 2014, 5, 715.
- Xia, X.; Li, Q.; Li, J.; Shao, C.; Zhang, J.; Zhang, Y.; Liu, X.; Lin, Y.; Liu, C.; She, Z. Two new derivatives of griseofulvin from the mangrove endophytic fungus Nigrospora sp. (strain No. 1403) from Kandelia candel (L.) Druce. Planta Med. 2011, 77, 1735–1738.
- Wen, L.; Lin, Y.-C.; She, Z.-G.; Du, D.-S.; Chan, W.-L.; Zheng, Z.-H. Paeciloxanthone, a new cytotoxic xanthone from the marine mangrove fungus Paecilomyces sp. (Tree1–7). J. Asian Nat. Prod. Res. 2008, 10, 133–137.
- Khamthong, N.; Rukachaisirikul, V.; Tadpetch, K.; Kaewpet, M.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Tetrahydroanthraquinone and xanthone derivatives from the marine-derived fungus Trichoderma aureoviride PSU-F95. Arch. Pharm. Res. 2012, 35, 461–468.
- Brunati, M.; Luis Rojas, J.; Sponga, F.; Ciciliato, I.; Losi, D.; Goettlich, E.; de Hoog, S.; Genilloud, O.; Marinelli, F. Diversity and pharmaceutical screening of fungi from benthic mats of Antarctic lakes. Mar. Genom. 2009, 2, 43–50.
- Ren, H.; Gu, Q.-Q.; Cui, C.-B. Anthraquinone derivatives produced by marine-derived Penicillium flavidorsum SHK1–27 and their antitumor activities. Chin. J. Med. Chem. 2007, 17, 148–154.
- Wang, P.L.; Li, D.Y.; Xie, L.R.; Wu, X.; Hua, H.M.; Li, Z.L. Two new compounds from a marine-derived fungus Penicillium oxalicum. Nat. Prod. Res. 2014, 28, 290–293.
- Klaiklay, S.; Rukachaisirikul, V.; Phongpaichit, S.; Pakawatchai, C.; Saithong, S.; Buatong, J.; Preedanon, S.; Sakayaroj, J. Anthraquinone derivatives from the mangrove-derived fungus Phomopsis sp. PSU-MA214. Phytochem. Lett. 2012, 5, 738–742.
- Zhou, X.-M.; Zheng, C.-J.; Chen, G.-Y.; Song, X.-P.; Han, C.-R.; Li, G.-N.; Fu, Y.-H.; Chen, W.-H.; Niu, Z.-G. Bioactive anthraquinone derivatives from the mangrove-derived fungus Stemphylium sp. 33231. J. Nat. Prod. 2014, 77, 2021–2028.
- Liu, Y.; Marmann, A.; Abdel-Aziz, M.S.; Wang, C.Y.; Müller, W.E.G.; Lin, W.H.; Mándi, A.; Kurtán, T.; Daletos, G.; Proksch, P. Tetrahydroanthraquinone Derivatives from the Endophytic Fungus Stemphylium globuliferum. Eur. J. Org. Chem. 2015, 2015, 2646–2653.
- Huang, X.; Sun, X.; Lin, S.; Xiao, Z.; Li, H.; Bo, D.; She, Z. Xylanthraquinone, a new anthraquinone from the fungus Xylaria sp. 2508 from the South China Sea. Nat. Prod. Res. 2014, 28, 111–114.
- Elix, J.A.; Wardlaw, J.H.; SØchting, U. Two new anthraquinones from the lichen Caloplaca spitsbergensis. Herzogia 2000, 14, 27–30.
- Berger, Y. 1, 3, 6, 8-tetrahydroxyanthraquinone from Aspergillus versicolor. Phytochemistry 1980, 19, 2779–2780.
- Xu, L.; Meng, W.; Cao, C.; Wang, J.; Shan, W.; Wang, Q. Antibacterial and antifungal compounds from Marine Fungi. Mar. Drugs 2015, 13, 3479–3513.
- Wang, W.-L.; Zhu, T.-J.; Tao, H.-W.; Lu, Z.-Y.; Fang, Y.-C.; Gu, Q.-Q.; Zhu, W.-M. Three novel, structurally unique spirocyclic alkaloids from the Halotolerant B-17 Fungal Strain of Aspergillus variecolor. Chem. Biodivers. 2007, 4, 2913–2919.
- Janso, J.E.; Bernan, V.S.; Greenstein, M.; Bugni, T.S.; Ireland, C.M. Penicillium dravuni, a new marine-derived species from an alga in Fiji. Mycologia 2005, 97, 444–453.
- Wang, X.; Mao, Z.G.; Song, B.B.; Chen, C.H.; Xiao, W.W.; Hu, B.; Wang, J.W.; Jiang, X.B.; Zhu, Y.H.; Wang, H.J. Advances in the study of the structures and bioactivities of metabolites isolated from mangrove-derived fungi in the South China Sea. Mar. Drugs 2013, 11, 3601–3616.
- Stevens, K.L.; Badar Ud, D.; Ahmad, A.; Ahmad, M. The antibiotic bostrycin from Alternaria eichhorniae. Phytochemistry 1979, 18, 1579–1580.
- Xu, C.; Wang, J.; Gao, Y.; Lin, H.; Du, L.; Yang, S.; Long, S.; She, Z.; Cai, X.; Zhou, S.; et al. The anthracenedione compound bostrycin induces mitochondria-mediated apoptosis in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2010, 10, 297–308.
- Xie, G.; Zhu, X.; Li, Q.; Gu, M.; He, Z.; Wu, J.; Li, J.; Lin, Y.; Li, M.; She, Z.; et al. SZ-685C, a marine anthraquinone, is a potent inducer of apoptosis with anticancer activity by suppression of the Akt/FOXO pathway. Br. J. Pharmacol. 2010, 159, 689–697.
- Kang, L.; Cai, M.; Yu, C.; Zhang, Y.; Zhou, X. Improved production of the anticancer compound 1403C by glucose pulse feeding of marine Halorosellinia sp. (No. 1403) in submerged culture. Bioresour. Technol. 2011, 102, 10750–10753.
- Zhu, X.; He, Z.; Wu, J.; Yuan, J.; Wen, W.; Hu, Y.; Jiang, Y.; Lin, C.; Zhang, Q.; Lin, M.; et al. A marine anthraquinone SZ-685C overrides adriamycin-resistance in breast cancer cells through suppressing Akt signaling. Mar. Drugs 2012, 10, 694–711.
- Wang, X.Y.; Tan, T.; Mao, Z.G.; Lei, N.; Wang, Z.M.; Hu, B.; Chen, Z.Y.; She, Z.G.; Zhu, Y.H.; Wang, H.J. The marine metabolite SZ-685C induces apoptosis in primary human nonfunctioning pituitary adenoma cells by inhibition of the Akt pathway in vitro. Mar. Drugs 2015, 13, 1569–1580.
- Dou, Y.; Wang, X.; Jiang, D.; Wang, H.; Jiao, Y.; Lou, H.; Wang, X. Metabolites from Aspergillus versicolor, an endolichenic fungus from the lichen Lobaria retigera. Drug Discov. Ther. 2014, 8, 84–88.
- Lee, Y.M.; Li, H.; Hong, J.; Cho, H.Y.; Bae, K.S.; Kim, M.A.; Kim, D.-K.; Jung, J.H. Bioactive metabolites from the sponge-derived fungus Aspergillus versicolor. Arch. Pharm. Res. 2010, 33, 231–235.
- Fouillaud, M.; Université de La Réunion, Saint Denis, France. Personal communication, 2016.
- Butinar, L.; Zalar, P.; Frisvad, J.C.; Gunde-Cimerman, N. The genus Eurotium—Members of indigenous fungal community in hypersaline waters of salterns. FEMS Microbiol. Ecol. 2005, 51, 155–166.
- Ren-Yi, G.; Lei, X.; Yi, K.; Iii-Ming, C.; Jian-Chun, Q.; Li, L.; Sheng-Xiang, Y.; Li-Chun, Z. Chaetominine, (+)-alantrypinone, questin, isorhodoptilometrin, and 4-hydroxybenzaldehyde produced by the endophytic fungus Aspergillus sp. YL-6 inhibit wheat (Triticum aestivum) and radish (Raphanus sativus) germination. J. Plant Interact. 2015, 10, 87–92.
- Debbab, A.; Aly, A.H.; Edrada-Ebel, R.; Wray, V.; Mueller, W.E.G.; Totzke, F.; Zirrgiebel, U.; Schaechtele, C.; Kubbutat, M.H.G.; Lin, W.H.; et al. Bioactive metabolites from the endophytic fungus Stemphylium globuliferum isolated from Mentha pulegium. J. Nat. Prod. 2009, 72, 626–631.
- Yang, W.J.; Yang, C.S.; Huang, C.J.; Chen, K.S.; Lin, S.F. Bostrycin, a novel coupling agent for protein immobilization and prevention of biomaterial-centered infection produced by Nigrospora sp. No. 407. Enzyme Microb. Technol. 2012, 50, 287–292.
- Furuya, K.; Shirasaka, M. Antibiotics from fungi. IV.production of rhodosporin (bostrycin) by Nigrospora oryzae. Sankyo Kenkyusho Nempo 1969, 21, 165–168.
- Crous, P.W.; Groenewald, J.Z. A phylogenetic re-evaluation of Athrinium. IMA Fungus 2013, 4, 133–154.
- van Eijk, G.W. Anthraquinones in the fungus Talaromyces stipitatus. Experientia 1973, 29, 522–523.
- Thomson, R.H. Naturally Occurring Quinones III: Recent Advances; Chapman & Hall: London, UK; New York, NY, USA, 1987.
- Noda, T.; Take, T.; Watanabe, T.; Abe, J. The structure of bostrycin. Tetrahedron 1970, 26, 1339–1346.
- Noda, T.; Take, T.; Otani, M.; Miyauchi, K.; Watanabe, T.; Abe, J. Structure of bostrycin. Tetrahedron Lett. 1968, 58, 6087–6090.
- Takenaka, A.; Furusaki, A.; Watanabe, T. The crystal and molecular structure of bostrycin pi-bromobenzoate, A derivative of bostrycin. Tetrahedron Lett. 1968, 6091–6094.
- Kelly, T.R.; Kim, M.H.; Curtis, A.D.M. Structure correction and synthesis of the naturally occurring benzothiazinone BMY 40662. J. Org. Chem. 1993, 58, 5855–5857.
- Wang, C.; Wang, J.; Huang, Y.; Chen, H.; Li, Y.; Zhong, L.; Chen, Y.; Chen, S.; Wang, J.; Kang, J.; et al. Anti-mycobacterial activity of marine fungus-derived 4-deoxybostrycin and nigrosporin. Molecules 2013, 18, 1728–1740.
- Kuhnert, E.; Heitkämper, S.; Fournier, J.; Surup, F.; Stadler, M. Hypoxyvermelhotins A–C, new pigments from Hypoxylon lechatii sp. nov. Fungal Biol. 2014, 118, 242–252.
- Hawksworth, D.L. Freshwater and marine lichen-forming fungi. In Aquatic Mycology across the Millennium; Hyde, K.D., Ho, W.H., Pointing, S.B., Eds.; Fungal Diversity Press: Hong Kong, China, 2000; Volume 5, pp. 1–7.
- Gauslaa, Y.; McEvoy, M. Seasonal changes in solar radiation drive acclimation of the sun-screening compound parietin in the lichen Xanthoria parietina. Basic Appl. Ecol. 2005, 6, 75–82.
- Larsson, P.; Večeřová, K.; Cempírková, H.; Solhaug, K.A.; Gauslaa, Y. Does UV-B influence biomass growth in lichens deficient in sun-screening pigments? Environ. Exp. Bot. 2009, 67, 215–221.
- Søchting, U.; Frödén, P. Chemosyndromes in the lichen genus Teloschistes (Teloschistaceae, Lecanorales). Mycol. Progress 2002, 1, 257–266.
- Søchting, U. Two major anthraquinone chemosyndromes in Teloschistaceae. Bibl. Lichenol. 1997, 68, 135–144.
- Søchting, U.; Lutzoni, F. Molecular phylogenetic study at the generic boundary between the lichen-forming fungi Caloplaca and Xanthoria (Ascomycota, Teloschistaceae). Mycol. Res. 2003, 107, 1266–1276.
- Søchting, U. Chemosyndromes with chlorinated anthraquinones in the lichen genus Caloplaca. In Lichenological Contributions in Honour of Jack Elix; McCarthy, P.M., Kantvilas, G., Louwhoff, S.H.J.J., Cramer, J., Eds.; Bibliotheca Lichenologica: Berlin, German, 2001; pp. 395–404.
- Frisvad, J.C.; Andersen, B.; Thrane, U. The use of secondary metabolite profiling in chemotaxonomy of filamentous fungi. Mycol. Res. 2008, 112, 231–240.
- Frisvad, J.C. Taxonomy, chemodiversity and chemoconsistency of Aspergillus, Penicillium and Talaromyces species. Front. Microbiol. 2015, 5.
- Gill, M.; Morgan, P.M. Pigments of fungi part 73—Absolute stereochemistry of fungal metabolites: Icterinoidins A1 and B1, and atrovirins B1 and B2. Arkivoc 2004, 152–165.
- Kurobane, I.; Vining, L.C.; McInnes, A.G.; Walter, J.A. Use of 13C in biosynthetic studies. The labeling pattern in dihydrofusarubin enriched from - and acetate in cultures of Fusarium solani. Can. J. Chem. 1980, 58, 1380–1385.
- Julia, P.; Martinkova, L.; Lolinski, J.; Machek, F. Ethanol as substrate for pigment production by the fungus Monascus purpureus. Enzyme Microb. Tech. 1994, 16, 996–1001.
- Cho, Y.J.; Hwang, H.J.; Kim, S.W.; Song, C.H.; Yun, J.W. Effect of carbon source and aeration rate on broth rheology and fungal morphology during red pigment production by Paecilomyces sinclairii in a batch bioreactor. J. Biotech. 2002, 95, 13–23.
- Cai, Y.; Din, Y.; Ta, G.; Lia, X. Production of 1,5-dihydroxy-3-methoxy-7-methylanthracene-9,10-dione by submerged culture of Shiraia bambusicola. J. Microbiol. Biotechnol. 2008, 18, 322–327.
- Yu, J.H.; Keller, N. Regulation of secondary metabolism in filamentous fungi. Annu. Rev. Phytopathol. 2005, 43, 437–458.
- Chooi, Y.H.; Tang, Y. Navigating the fungal polyketide chemical space: From genes to molecules. J. Org. Chem. 2012, 77, 9933–9953.
- Miethbauer, S.; Haase, S.; Schmidtke, K.; Gunther, W.; Heiser, I.; Liebermann, B. Biosynthesis of photodynamically active rubellins and structure elucidation of new anthraquinone derivatives produced by Ramularia collo-cygni. Phytochemistry 2006, 67, 1206–1213.
- Kroken, S.; Glass, N.L.; Taylor, J.W.; Yoder, O.C.; Turgeon, B.G. Phylogenomic analysis of type 1 polyketide synthase genes in pathogenic and saprobic ascomycetes. Natl. Acad. Sci. 2003, 100, 15670–15675.
- Brown, D.W.; Butchko, R.A.; Baker, S.E.; Proctor, R.H. Phylogenomic and functional domain analysis of polyketide synthases in Fusarium. Fungal Biol. 2012, 116, 318–331.
- Zabala, A.O.; Xu, W.; Chooi, Y.H.; Tang, Y. Characterization of a silent azaphilone gene cluster from Aspergillus niger ATCC 1015 reveals a hydroxylation-mediated pyran-ring formation. Chem. Biol. 2012, 19, 1049–1059.
- Cacho, R.A.; Tang, Y.; Chooi, Y.-H. Next-generation sequencing approach for connecting secondary metabolites to biosynthetic gene clusters in fungi. Front. Microbiol. 2014, 5, 774.
- Javidpour, P.; Korman, T.P.; Shakya, G.; Tsai, S.C. Structural and biochemical analyses of regio- and stereospecificities observed in a type II polyketide ketoreductase. Biochemistry 2011, 50, 4638–4649.
- Liu, L.; Zhang, Z.; Shao, C.-L.; Wang, J.-L.; Bai, H.; Wang, C.-Y. Bioinformatical analysis of the sequences, structures and functions of fungal polyketide synthase product template domains. Sci. Rep. 2015, 5, 10463.
- Bringmann, G.; Noll, T.F.; Gulder, T.A.; Grune, M.; Dreyer, M.; Wilde, C.; Pankewitz, F.; Hilker, M.; Payne, G.D.; Jones, A.L.; et al. Different polyketide folding modes converge to an identical molecular architecture. Nat. Chem. Biol. 2006, 2, 429–433.
- O’Hagan, D. Biosynthesis of polyketide metabolites. Nat. Prod. Rep. 1992, 9, 447–479.
- Elsworth, C.; Gill, M.; Gimenez, A.; Milanovic, N.M.; Raudies, E. Pigments of fungi. Part 50.1 Structure, biosynthesis and stereochemistry of new dimeric dihydroanthracenones of the phlegmacin type from Cortinarius sinapicolor Cleland. J. Chem. Soc. Perkin Trans. 1999, 1999, 119–126.
- Chen, Z.G.; Fujii, I.; Ebizuka, Y.; Sankawa, U. Emodin O-methyltransferase from Aspergillus terreus. Arch. Microbiol. 1992, 158, 29–34.
- Gill, M. Pigments of fungi (Macromycetes). Nat. Prod. Rep. 1999, 16, 301–317.
- Gill, M.; Morgan, P.M. New fungal anthraquinones. Arkivoc 2001, 2001, 145–156.
- Michal, G. Lysin-Stoffwechsel. In Biochemical pathways; Michal, G., Ed.; Spektrum Akademischer Verlag Heidelberg: Berlin, German, 1999; pp. 51–54.
- Henry, K.M.; Townsend, C.A. Synthesis and fate of o-carboxybenzophenones in the biosynthesis of aflatoxin. J. Am. Chem. Soc. 2005, 127, 3300–3309.
- Sanchez, J.F.; Somoza, A.D.; Keller, N.P.; Wang, C.C. Advances in Aspergillus secondary metabolite research in the post-genomic era. Nat. Prod. Rep. 2012, 29, 351–371.
- Bringmann, G.; Irmer, A.; Feineis, D.; Gulder, T.A.M.; Fiedler, H.-P. Convergence in the biosynthesis of acetogenic natural products from plants, fungi, and bacteria. Phytochemistry 2009, 70, 1776–1786.
- Kakule, T.B.; Lin, Z.; Schmidt, E.W. Combinatorialization of fungal polyketide synthase-peptide synthetase hybrid proteins. J. Am. Chem. Soc. 2014, 136, 17882–17890.
- Mapari, S.A.S. Chemotaxonomic Exploration of Fungal Biodiversity for Polyketide Natural Food Colorants… Discovery & Evaluation of cell Factories, and Characterization of Pigments. Ph.D. Thesis, Technical University of Denmark, Lyngby, Denmark, 2009.
- Bu’Lock, J.D.; Detroy, R.W.; Hošťálek, Z.; Munim-Al-Shakarchi, A. Regulation of secondary biosynthesis in Gibberella fujikuroi. Trans. Br. Mycol. Soc. 1974, 62, 377–389.
- Pagano, M.C.; Dhar, P.P. Fungal Pigments: An Overview. In Fungal Biomolecules: Sources, Applications and Recent Developments; Gupta, V.K., Mach, R.L., Sreenivasaprasad, S., Eds.; Wiley-Blackwell: Pondichery, India, 2015; pp. 173–181.
- Hiort, J.; Maksimenka, K.; Reichert, M.; Perović-Ottstadt, S.; Lin, W.H.; Wray, V.; Steube, K.; Schaumann, K.; Weber, H.; Proksch, P.; et al. New natural products from the sponge-derived fungus Aspergillus niger. J. Nat. Prod. 2004, 67, 1532–1543.
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459.
- Margalith, P. Pigment Microbiology; Springer Netherlands: London, UK; New York, NY, USA, 1992; p. 156.
- Duran, N.; de Conti, R.; Teixeira, M.F.S. Pigments from fungi : Industrial perspective. In Advances in Fungal Biotechnology; Rai, M., Ed.; I.K. International Publishing House Pvt. Ltd.: New Delhi, India, 2009; pp. 185–225.

