Anthraquinones and their derivatives constitute a large group of quinoid compounds with about 700 molecules described. They are widespread in fungi and their chemical diversity and biological activities attracted attention of industries in such fields as pharmaceuticals, clothes dyeing, and food colorants. As marine microorganisms appeared as producers of an astonishing variety of structurally unique secondary metabolites, they may represent a promising resource for identifying new candidates for therapeutic drugs or daily additives.
- anthraquinone
- marine
- fungi
- pigment
- biological activity
- antitumor
- antibiotic
- cytotoxicity
1. Introduction
2. Anthraquinones from Marine-Derived Fungi
2.1. Anthraquinone′s Basic Structure
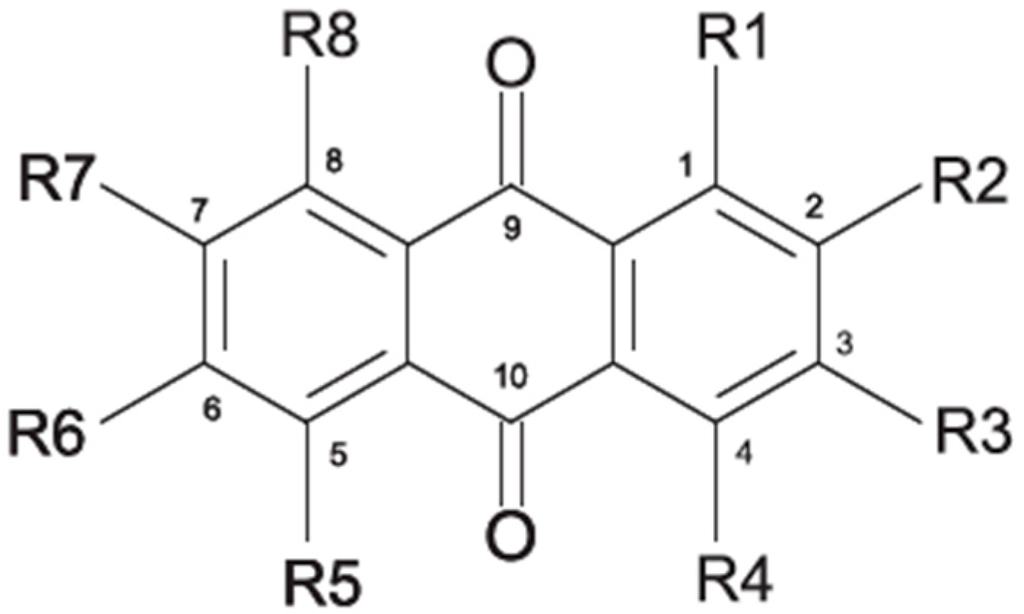
2.2. Ecology of Marine-Derived Fungal Anthraquinones Producers
-
obligate marine fungi (true ones) that grow and sporulate only in seawater. Their spores are able to germinate and form new thalli in salted environment.
-
transitional marine fungi (marine-derived fungi) that come from terrestrial or freshwater media and have undergone physiological adaptation to survive, grow, or reproduce in the marine environment.

2.3. Structural Diversity and Colors of Anthraquinoid Extrolites from Marine-Derived Fungi
2.3.1. Present Knowledge about Anthraquinonoid Compounds from Fungi
2.3.2. Nature and Colors of Compounds from Marine-Derived Fungi
Genera and Species
|
Genus |
Species/Strain No |
Name of Compounds Produced |
Source of Isolation |
Refs. |
|---|---|---|---|---|
|
Alternaria |
Al. eichorniae |
4-deoxyBostrycin, Bostrycin |
Mar. Plant pathogen |
[46] |
|
Al. (SK11) |
(+) α S-alterporriol C, 6-methylquinizarin, Alterporriol S, Austrocortinin |
Mangrove Plant end. |
[47] |
|
|
Al. sp. ZJ-2008003 |
Alterporriol C, K–R, Altersolanol B and C, Macrosporin |
Mar. Org end. |
[48] |
|
|
Al. sp. ZJ9-6B |
Alterporriols C–M, Altersolanol A, Dactylariol, Macrosporin, Physcion, TetrahydroAltersolanol B |
Mar. Plant end. |
[49] |
|
|
Aspergillus |
A. glaucus |
10,10′-dimer of Emodin and Physcion, Catenarin, Cynodontin, Emodin, Erythroglaucin, Helminthosporin, Physcion, Questin, Rubrocristin, Tritisporin, Variecolorquinone A |
Mangrove sed. |
|
|
A. sp. 05F16 |
1-deoxytetrahydrobostrycin, Tetrahydrobostrycin |
Algal end. |
[54] |
|
|
A. sp. SCSIOF063 |
(1′S)-7-chloroaverantin, 1′-O-methylaverantin 1′-O-methyl-7-chloroaverantin, 6-O-methyl-7-chloroaverantin, 6-O-methyl-7-chloroaverythrin, 6-O-methyl-7-bromoaverantin, 6,1′-O,O-dimethyl-7-chloroaverantin, 6,1′-O,O-dimethyl-7-bromoaverantin, 6,1′-O,O-dimethylaverantin, 7-chloroaverantin-1′-butyl ether, 7-chloroaverythrin |
Sed. |
[55] |
|
|
A. variecolor B-17 |
(2S)-2,3-dihydroxypropyl1,6,8-trihydroxy-3-methyl-9,10-dioxoanthracene-2carboxylate, Catenarin, Emodin, Fallacinol, Physcion, Erythroglaucin, Questin, Questinol, Rubrocristin, Variecolorquinone A, |
Sed. |
[56] |
|
|
A. versicolor |
7-hydroxyemodin 6,8-methyl ether, Emodin, Isorhodoptilometrin-methyl ether, Methyl emodin |
Algal end. |
[57] |
|
|
A. versicolor EN-7 (Genbank no EU042148) |
6,8-di-O-methylversiconol 6,8-di-O-methylnidurufin 6,8-di-O-methylaverantin 6,8-di-O-methylversicolorin A, Aversin: (−)-isomer |
Algal end. |
[58] |
|
|
Curvularia |
C. lunata |
Cytoskyrin A, Lunatin |
Mar. Org end. |
|
|
Eurotium |
E. cristatum (ECE) |
Catenarin, Emodin, Erythroglaucyn, Physcion, Physcion anthrone, Questin, Rubrocristin |
Mar. Org end. |
|
|
E. repens |
Catenarin, Erythroglaucyn, Physcion, Physcion anthrone |
Mar. Org end. |
||
|
E. rubrum |
6,3-O-(α-d-ribofuranosyl)-questin, Questin |
Mar. Plant end. |
[63] |
|
|
Unidentified |
Fungus Isolate 1850 and 2526 |
Averufin, Nidurufin, versicolorin C |
Mar. Plant end. |
[64] |
|
Fungus ZSUH-36 |
1′-O-methyl averantin, 6,8-di-O-methyl averufanin, 6,8-di-O-methyl averufin, 6,8,1′-tri-O-methyl averantin, Versicolorin C |
Mar. Plant end. |
[65] |
|
|
Fusarium |
F. sp. No. B77 |
5-acetyl-2-methoxy-1,4,6-trihydroxy-anthraquinone |
Mangrove Plant end. |
[66] |
|
F. sp. ZZF60 |
6,8-dimethoxy-1-methyl-2-(3-oxobutyl) anthraquinone |
Mangrove Plant end. |
[67] |
|
|
F. sp. No. ZH-210 |
Fusaquinon A,B,C |
Mangrove sed. |
[68] |
|
|
F. sp. PSU-F14, F. sp. PSU-F135 |
Austrocortirubin, Bostrycin |
Mar. Org end. |
[69] |
|
|
Halorosellinia |
H. sp. (No. 1403) |
1,4,5,6,7,9-hexahydroxy-2-methoxy-7-methyl-5β,9β, 8aβ,6α,10aα—hexahydroanthracene10(10aH)-one, Austrocortirubin, Demethoxyaustrocortirubin, Hydroxy-9,10-anthraquinone, SZ-685C |
Mar. Plant-derived |
|
|
Lichens |
Arthonia elegans, Biatorella conspersa, B. ochrophora, Pyrenula cerina, Sphaerophorus fragilis, Stereocaulon corticatulum,v. procerum, Trypethelium aeneum, T. aureomaculata, etc. |
Physcion |
Lichens |
[72] |
|
Caloplaca sp. |
Phallacinol (=Teloschistin=Fallacinol)) |
Lichen |
||
|
Caloplaca ehrenbergii, C. schaereri, C. spitsbergensis, etc. |
1-O-methyl-7-chloroemodin, 7-chloro-1,6,8-trihydroxy-3-methyl-10-anthrone, 7-chlorocitreorosein, 7-chloroemodic acid, 7-chloroemodin, 7-chloroemodinal, Emodin, Phallacinol, Physcion |
Lichens |
||
|
Gliocladium sp. T 31 |
Citreorosein, Emodin, Isorhodoptilometrin |
Lichen |
[78] |
|
|
Letrouitia hafellneri, L. leprolytoides |
7-chloroemodinal, 7-chloroemodin, Fragilin, Physcion |
Lichens |
||
|
Microsphaeropsis |
M. sp. |
1,3,6,8-tetrahydroxyanthraquinone, 1,3,6,8-tetrahydroxy-2-(1-hydroxyethyl)anthraquinone 1,3,6,8-tetrahydroxy-2-(1-methoxyethyl)anthraquinone 1,2,3,6,8-pentahydroxy-7-(1-methoxyethyl)anthraquinone |
Mar. Org end. |
[81] |
|
Monodictys |
M. sp. |
Chrysophanol, Emodin, Monodictyquinone A, Pachybasin |
Mar. Org end. |
|
|
Nigrospora |
N. spp. |
1-deoxytetrahydrobostrycin, 4-deoxybostrycin, Bostrycin, 4a-epi-9α-methoxydihydrodeoxybostrycin, 10-deoxybostrycin |
Mar. Plant/Org end. |
|
|
N. sp. MA75 |
4-deoxybostrycin, Bostrycin |
Marine |
[85] |
|
|
N. sp. 1403 |
4-deoxybostrycin, Bostrycin |
Mangrove |
[86] |
|
|
Paecilomyces |
P. sp. (Tree 1-7) |
Chrysophanol, Emodin |
Mangrove |
[87] |
|
Penicillium |
P. citrinum PSU-F51 (Accession no JQ66600) |
Chrysophanol, Citreorosein, Emodin, Penicillanthranins A and B |
Mar. Org end. |
[88] |
|
P. chrysogenum |
Skyrin |
Salt lake |
[89] |
|
|
P. flavidorsum SHK1-27 |
6,8-O-dimethylaverufin, 8-O-methylaverufin, Averufin, Averantin, Versiconol, Versicolorin A&B, Nidurufin |
Marine |
[90] |
|
|
P. oxalicum 2-HL-M-6 |
Aloe emodin, Chrysophanol, Citreorosein, Citreorosein-3-O-sulfate, Emodin, Emodin-3-O-sulfate, Isorhodoptilometrin |
Mangrove sed. |
[91] |
|
|
Phomopsis |
P. sp. PSU-MA214 |
Phomopsanthraquinone, 1-hydroxy-3-methoxy-6-methylanthraquinone, Ampelanol, Macrosporin |
Mangrove Plant endo |
[92] |
|
Stemphylium |
S. sp. 33231 |
2-O-acetylaltersolanol B, Alterporriol T–W, Altersolanol B&C, Auxarthrol C, Macrosporin |
Mangrove Plant end. |
[93] |
|
S. globuliferum |
6-O-methylalaternin, Acetylalterporriol D and E, Alterporriol D and E, Altersolanol A,B and C, Dihydroaltersolanol B and C, Macrosporin, Stemphylanthranol A and B |
Salt lake Plant end. |
[94] |
|
|
Trichoderma |
T. aureoviride PSU-F95 |
Coniothranthraquinone 1, Trichodermaquinone |
Mar. Org end. |
[88] |
|
Xylaria |
X. sp. 2508 |
Altersolanol A, Bostrycin, Deoxybostrycin, Xylanthraquinone |
Marine |
[95] |
|
Mol. Formula/Mol. mass |
Trivial name |
Structure |
IUPAC NAME |
Source |
Refs |
|---|---|---|---|---|---|
|
C14H8O3/224 |
Hydroxy-9,10-anthraquinone |
|
1-hydroxy-3-methylanthraquinone |
Halorosellinia sp. No. 1403 |
[70] |
|
C15H10O3/238 |
Pachybasin |
|
1-hydroxy-3-methylanthraquinone |
Monodictys sp. |
|
|
C15H10O4/254 |
Chrysophanol |
|
1,8-dihydroxy-3-methylanthraquinone |
Monodictys sp. Paecilomyces sp. P. citrinum PSU-F51 P. oxalicum 2-HL-M-6 |
|
|
C15H10O4/254 |
6-Methylquinizarin |
|
1,4-dihydroxy-methylanthraquinone |
Al. sp. (SK11) |
[47] |
|
C16H12O4/268 |
1-Hydroxy-3-methoxy-6-methylanthraquinone |
|
---- |
Phomopsis sp. PSU-MA214 |
[92] |
|
C15H10O5/270 |
Aloe emodin |
|
1,8-dihydroxy-3-(hydroxymethyl)anthraquinone |
P. oxalicum 2-HL-M-6 |
[91] |
|
C15H10O5/270 |
Emodin |
|
1,3,8-trihydroxy-6-methylanthraquinone |
A. glaucus A. variecolor B-17 A. versicolor Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) Eurotium Cristatum Gliocladium sp. T31 Monodictys sp. Paecilomyces sp. P. citrinum PSU-F51 P. oxalicum 2-HL-M-6 |
[91] [56] [61] [75,76,77,79,96] [36] [78] [14,82] [87] [88] [57] |
|
C15H10O5/270 |
Helminthosporin |
|
1,5,8-trihydroxy-3-methylanthraquinone |
A. glaucus |
|
|
C14H8O6/272 |
1,3,6,8-Tetrahydroxyanthraquinone |
|
---- |
A.versicolor Microsphaeropsis |
|
|
C15H14O5/274 |
Coniothranthraquinone 1 |
|
(2S,3R)-2,3,5-trihydroxy-7-methyl-1,2,3,4-tetrahydroanthraquinone |
Trichoderma aureoviride (PSU-F95) |
|
|
C16H12O5/284 |
1-Methylemodin |
|
1,3-dihydroxy-8-methoxy-6-methylanthraquinone |
A. versicolor |
[57] |
|
C16H12O5/284 |
Austrocortinin |
|
1,4-dihydroxy-2-methoxy-7-methylanthraquinone |
Al. sp. (SK11) |
[47] |
|
C16H12O5/284 |
Macrosporin |
|
1,7-dihydroxy-3-methoxy-6-methylanthraquinone |
Al. sp. ZJ9-6B Al. sp. ZJ-2008003 Phomopsis sp. PSU-MA214 Stemphylium globuliferum Stemphylium sp. 33231 |
|
|
C16H12O5/284 |
Marcrospin |
|
1,6-dihydroxy-3-methoxy-7-methylanthraquinone |
Al. sp. ZJ9-6B |
[49] |
|
C16H12O5/284 |
Monodictyquinone A |
|
1,8-dihydroxy-2-methoxy-6-methylanthraquinone |
Monodictys sp. |
|
|
C16H12O5/284 |
Phallacinol/Fallacinol |
|
1,8-dihydroxy-3-(hydroxy-methyl)-6-methoxyanthraquinone |
A. variecolor B-17 Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) |
|
|
C16H12O5/284 |
Physcion |
|
1,8-dihydroxy-3-methoxy-6-methylanthraquinone |
Al. sp. ZJ9-6B A. glaucus A. variecolor B-17 Eurotium repens Eurotium cristatum Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) Letrouitia hafellneri, L. leprolytoides, Arthonia elegans, Biatorella conspersa, B. ochrophora, Pyrenula cerina, Sphaerophorus fragilis, Stereocaulon corticatulum, v. procerum, Trypethelium aeneum, T. aureomaculata |
[49] [50,51,52,53] [56] [62] [36,61] [75,76,77,79,96] [72,80] |
|
C16H12O5/284 |
Questin |
|
1,6-dihydroxy-8-methoxy-3-methylanthraquinone |
A. glaucus A. variecolor B-17 Eurotium cristatum (ECE) Eurotium rubrum |
|
|
C15H10O6/286 |
Catenarin |
|
1,4,5,7-tetrahydroxy-2-methylanthraquinone |
A. glaucus A. variecolor B-17 Eurotium cristatum (ECE) Eurotium repens |
|
|
C15H10O6/286 |
Citreorosein |
|
ω-hydroxyemodin (OHM) or 1,3,8-trihydroxy-6-(hydroxymethyl) anthraquinone |
Gliocladium. sp. T31 P. citrinum PSU-F51 P. oxalicum 2-HL-M-6 |
|
|
C15H10O6/286 |
Cynodontin |
|
1,4,5,8-tetrahydroxy-2-methylanthraquinone |
A. glaucus |
|
|
C15H10O6/286 |
Lunatin |
|
1,3,8-trihydroxy-6-methoxyanthraquinone |
Curvularia lunata |
|
|
C15H10O7/286 |
Tritisporin |
|
1,4,5,7-tetrahydroxy-2-(hydroxylmethyl) anthraquinone |
A. glaucus |
|
|
C15H14O6/290 |
Trichodermaquinone |
|
(2S,3R)-2,3,5-trihydroxy-7-(hydroxylmethyl)-1,2,3,4-tetrahydroanthraquinone |
Trichoderma aureoviride (PSU-F95) |
|
|
C15H14O6/290 |
Demethoxyaustrocortirubin |
|
1,4-dihydroxy-6-methylanthraquinone |
Halorosellinia sp. No. 1403 |
|
|
C15H14O6/290 |
7-Hydroxyemodin 6,8-methyl ether |
|
2,8-dihydroxy-1,3-dimethoxy-6-methyl anthraquinone |
A. versicolor |
[57] |
|
C16H10O4/300 |
Erythroglaucin |
|
1, 4, 5-trihydroxy-7-methoxy-2-methylanthraquinone |
A. glaucus A. variecolor B-17 Eurotium cristatum (ECE) Eurotium repens |
|
|
C16H12O6/300 |
Carviolin |
|
1,3-dihydroxy-6-(hydroxymethyl)-8-methoxyanthraquinone |
P. dravuni |
[100] |
|
C16H12O6/300 |
Questinol |
|
1,6-dihydroxy-3-(hydroxymethyl)-8-methoxyanthraquinone |
A. variecolor B-17 |
[56] |
|
C16H12O6/300 |
Rubrocristin |
|
1,4,7-trihydroxy-5-methoxy-2-methylanthraquinone |
A. glaucus A. variecolor B-17 Eurotium cristatum (ECE) |
|
|
C16H12O6/300 |
6-O-methylalaternin |
|
1,2,8-trihydroxy-6-methoxy-3-methylanthraquinone |
Stemphylium globuliferum |
[94] |
|
C16H12O6/300 |
1,4,6-Trihydroxy-2-methoxy-7-methyl-anthraquinone |
|
3,5,8-trihydroxy-7-methoxy-2-methylanthraquinone |
Halorosellinia sp. No. 1403 |
[70] |
|
C15H9O5Cl/304 |
7-Chloroemodin |
|
---- |
Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) |
|
|
C16H16O6/304 |
Altersolanol B |
|
(2S,3R)-2,3,5-trihydroxy-7-methoxy-2-methyl-1,2,3,4-tetrahydroanthraquinone |
Al. sp. ZJ-2008003 Stemphylium sp. 33231 |
|
|
C16H18O6/306 |
Fusaquinon A |
|
(2R,3S,4aR,9S,9aS)-3,5,8-trihydroxy-7-methoxy-2-methyl-2,3,4,4a,9,9a-hexahydro-2,9-epoxyanthracen-10(1H)-one |
Fusarium sp. No. ZH-210 |
[68] |
|
C16H18O6/307 |
Dihydroaltersolanol B |
|
(2S,3R)-2,3,5-trihydroxy-7-methoxy-2-methyl-1,2,3,4,4a,9a-hexahydroanthraquinone |
Stemphylium globuliferum |
[94] |
|
C15H19O7/311 |
Xylanthraquinone |
|
---- |
Xylaria sp. 2508 |
[95] |
|
C17H14 O6/314 |
Isorhodoptilometrin |
|
(R)-1,3,8-trihydroxy-6-(2-hydroxypropyl)anthraquinone |
Gliocladium sp. T31 P. oxalicum 2-HL-M-6 |
|
|
C16H12O7/317 |
1,3,6,8-Tetrahydroxy-2-(1-hydroxyethyl) anthraquinone |
|
1,3,6,8-tetrahydroxy-2-(1-hydroxyethyl) anthracene-9,10-dione |
Microsphaeropsis |
[81] |
|
C16H11O5Cl/318 |
1-O-Methyl-7-chloroemodin |
|
2-chloro-1,6-dihydroxy-8-methoxy-3-methylanthraquinone |
Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) |
|
|
C15H9O6Cl/320 |
7-Chlorocitreorosein |
|
---- |
Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) |
|
|
C16H16O7/320 |
Austrocortirubin |
|
1,4-dihydroxy-2-methoxy-7-methylanthraquinone |
Fusarium spp. PSU-F14 and PSU-F135 Halorosellinia sp. No. 1403 Nigrospora sp. ZJ-2010006 |
|
|
C16H16O7/320 |
Altersolanol C |
|
(1R,2R,3R)-1,2,3,5-tetra-hydroxy-7-methoxy-2-methyl-1,2,3,4-tetra-hydroanthraquinone |
Al. sp. ZJ9-6B Al. sp. ZJ-2008003 Stemphylium sp. 33231 |
|
|
C16H16O7/320 |
4-Deoxybostrycin |
|
(2R,3S,4aS,9aS,10R)-2,3,5,8,10-pentahydroxy-6-methoxy-3-methyl-1,3,4,4a,9a,10-hexahydroanthracen-9(2H)-one |
Nigrospora sp. 1403 Nigrospora sp. MA75 |
|
|
C16H16O7/320 |
2,3,5,8-Tetrahydroxy-7-methoxy-2-methyl-1,2,3,4-tetrahydroanthraquinone |
---- |
2,3,5,8-tetrahydroxy-7-methoxy-2-methyl-1,2,3,4-tetrahydroanthracene-9,10-dione |
Al. eichorniae |
[46] |
|
C16H16O7/321 |
10-Deoxybostrycin |
|
--- |
Nigrospora sp. |
[84] |
|
C16H18O7/323 |
Dihydroaltersolanol C |
|
(1R,2R,3R)-1,2,3,5-tetra-hydroxy-7-methoxy-2-methyl-1,2,3,4,4a,9a-hexahydroanthraquinone |
Stemphylium globuliferum |
[94] |
|
C16H20O7/324 |
Fusaquinon C |
|
(2S,3R,4aR,9aR,10S)-2,3,5,8,10-pentahydroxy-6-methoxy-3-methyl-1,3,4,4a,9a,10-hexahydroanthracen-9(2H)-one |
Fusarium sp. No. ZH-210 |
[68] |
|
C16H21O7/325 |
1-Deoxytetrahydrobostrycin |
|
(2R,3S)-2,3,5,8,10-pentahydroxy-6-methoxy-3-methyl-1,3,4,4a,9a,10-hexahydroanthracen-9(2H)-one |
A. sp. 05F16 Nigrospora sp. |
|
|
C15H20O8/328 |
Fragilin |
|
2-chloro-1,8-dihydroxy-3-methoxy-6-methylanthraquinone |
Letrouitia hafellneri L. leprolytoides |
|
|
C17H12O7/328 |
5-Acetyl-2-methoxy-1,4,6-trihydroxyanthraquinone |
|
---- |
Fuarium sp. B77 |
[66] |
|
C18H16O6/328 |
Isorhodoptilometrin-1-methylether |
|
1,3-dihydroxy-6-2-hydroxypropyl-8-methoxyanthraquinone |
A. versicolor |
[57] |
|
C17H14O7/329 |
1,3,6,8-Tetrahydroxy-2-(1-methoxyethyl)anthraquinone |
|
--- |
Microsphaeropsis |
[81] |
|
C18H20O6/332 |
Phomopsanthraquinone |
|
(2R,3S)-7-ethyl-1,2,3,4-tetrahydro-2,3,8-trihydroxy-6-methoxy-3-methylanthraquinone |
Phomopsis sp. PSU-MA214 |
[92] |
|
C16H16O8/336 |
Altersolanol A |
|
(1R,2S,3R,4S)-1,2,3,4,5-pentahydroxy-7-methoxy-2-methyl-1,2,3,4-tetrahydroanthraquinone |
Stemphylium globuliferum Xylaria sp. 2508 |
|
|
C16H16O8/336 |
Bostrycin |
|
(5S,6R,7S)-5,6,7,9,10-pentahydroxy-2-methoxy-7-methyl-5,6,7,8-tetrahydroanthracene-1,4-dione |
A. sp. strain 05F16 Al. eichorniae Fusarium spp. PSU-F14/PSU-F135 Halorosellinia sp. No. 1403 Nigrospora sp. Xylaria sp. 2508 |
|
|
C16H18O8/338 |
SZ-685C |
|
1,2,3,5,8-pentahydroxy-6-methoxy-3-methyl-1,2,3,4-tetrahydroanthraquinone |
Halorosellinia sp. No. 1403 |
|
|
C16H20O8/340 |
Fusaquinon B |
|
(1R,2S,3R,4aR,9aS,10S)-1,2,3,5,8,10-hexahydroxy-6-methoxy-3-methyl-1,3,4,4a,9a,10-hexahydroanthracen-9(2H)-one |
Fusarium sp. No. ZH-210 |
[68] |
|
C16H21O8/340 |
Tetrahydroxybostrycin |
|
1,2,3,5,8,10-hexahydroxy-6-methoxy-3-methyl-1,3,4,4a,9a,10-hexahydroanthracen-9(2H)-one |
A. sp. 05F16 Nigrospora sp. MA75 |
|
|
C18H12O7/340 |
Versicolorin C |
|
4,6,8-trihydroxy-3,3a-dihydroanthra[2,3-b]furo[3,2-d]furan-5,10(2H,12aH)-dione |
Fungus ZSUH-36 Fungus Isolate 1850 and isolate 2526 |
|
|
C18H18O7/346 |
2-O-Acetylaltersolanol B |
|
(2R,3S)-3,8-dihydroxy-6-methoxy-3-methyl-9,10-dioxo-1,2,3,4,9,10-hexahydroanthracen-2-yl acetate |
Stemphylium sp. 33231 |
[93] |
|
C17H14O8/347 |
1,2,3,6,8-Pentahydroxy-7-(1-methoxyethyl)anthraquinone |
|
1,2,3,6,8-pentahydroxy-7-(1-methoxyethyl)anthracene-9,10-dione |
Microsphaeropsis |
[81] |
|
C15H10O8S/350 |
Emodin-3-O Sulfate |
|
4,5-dihydroxy-7-methyl-9,10-dioxo-9,10-dihydroanthracen-2-yl hydrogen sulfate |
P. oxalicum 2-HL-M-6 |
[91] |
|
C16H15O9/351 |
Auxarthrol C |
|
(1S,2R,3R,4R,4aR,9aS)-1,2,3,4,5-pentahydroxy-7-methoxy-2-methyl-1,2,3,4-tetrahydro-4a,9a-epoxyanthraquinone |
Stemphylium sp. 33231 |
[93] |
|
C19H11O7/351 |
8-O-MethylversicolorinB |
|
4,8-dihydroxy-6-methoxy-3,3a-dihydroanthra[2,3-b]furo[3,2-d]furan-5,10(2H,12aH)-dione |
A. versicolor endolichenic |
[108] |
|
C21H20O5/352 |
6,8-Dimethoxy-1-methyl-2-(3-oxobutyl)anthraquinone |
|
---- |
Fusarium sp. ZZF60 |
[67] |
|
C19H13O7/353 |
8-O-Methylversicolorin A |
|
4,8-dihydroxy-6-methoxyanthra[2,3-b]furo[3,2-d]furan-5,10(3aH,12aH)-dione |
A. versicolor endolichenic |
[108] |
|
C20H18O6/354 |
Averythrin |
|
(E)-2-(hex-1-en-1-yl)-1,3,6,8-tetrahydroxyanthraquinone |
A. sp. SCSIO F063 A. versicolor endolichenic |
|
|
C30H18O10/358 |
Skyrin |
|
2,2′,4,4′,5,5′-hexahydroxy-7,7′-dimethyl-[1,1′-bianthracene]-9,9′,10,10′-tetraone |
P. chrysogenum |
[89] |
|
C16H12O8S/364 |
Macrosporin-7-O-sulfate |
|
Sodium 8-hydroxy-6-methoxy-3-methyl-9,10-dioxo-9,10-dihydroanthracen-2-yl sulfate |
Stemphylium sp. 33231 |
[93] |
|
C15H9O9S/365 |
Citreorosein-3-O-sulfate |
|
4,5-dihydroxy-7-(hydroxymethyl)-9,10-dioxo-9,10-dihydroanthracen-2-yl hydrogen sulfate |
P. oxalicum 2-HL-M-6 |
[91] |
|
C20H14O7/365 |
6,8-di-O-methylversico-lorinA |
|
4-hydroxy-6,8-dimethoxyanthra[2,3-b]furo[3,2-d]furan-5,10(3aH,12aH)-dione |
A. versicolor endolichenic A. versicolor EN-7 (Genbank no EU042148) |
|
|
C21H19O6/367 |
8-O-Methylaverythrin |
|
(E)-2-(hex-1-en-1-yl)-1,3,6-trihydroxy-8-methoxyanthraquinone |
A. versicolor endolichenic |
[108] |
|
C20H16O7/368 |
Aversin |
|
4-hydroxy-6,8-dimethoxy-3,3a-dihydroanthra[2,3-b]furo[3,2-d]furan-5,10(2H,12aH)-dione |
A. versicolor endolichenic |
[108] |
|
C20H16O7/368 |
Aversin : (−)-isomer |
|
4-hydroxy-6,8-dimethoxy-3,3a-dihydroanthra[2,3-b]furo[3,2-d]furan-5,10(2H,12aH)-dione |
A. versicolor EN-7 (Genbank no EU042148) |
[58] |
|
C20H20O7/372 |
Averantin |
|
(S)-1,3,6,8-tetrahydroxy-2-(1-hydroxyhexyl)anthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C20H20O7/372 |
Averantin = (S)-(−)-averantin |
|
(S)-1,3,6,8-tetrahydroxy-2-(1-hydroxyhexyl)anthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C21H18O7/382 |
6-O-Methylaverufin |
|
7,9-dihydroxy-11-methoxy-2-methyl-3,4,5,6-tetrahydro-2H-2,6-epoxyanthra[2,3-b]oxocine-8,13-dione |
Fungus ZSUH-36 A. versicolor EN-7 |
|
|
C20H16O8/384 |
Nidurufin |
|
5,7,9,11-tetrahydroxy-2-methyl-3,4,5,6-tetrahydro-2H-2,6-epoxyanthra[2,3-b]oxocine-8,13-dione |
Fungus Isolate 1850 and isolate 2526 |
[64] |
|
C20H15O8/386 |
Averufin |
|
7,9,11-trihydroxy-2-methyl-3,4,5,6-tetrahydro-2H-2,6-epoxyanthra[2,3-b]oxocine-8,13-dione |
A. versicolor Fungus ZSUH-36 Fungus Isolate 1850 and isolate 2526 |
|
|
C21H22O7/386 |
1′-O-Methylaverantin |
|
1,3,6,8-tetrahydroxy-2-(1-methoxyhexyl)anthraquinone |
A. sp. SCSIO F063 Fungus ZSUH-36 |
|
|
C19H15O9/388 |
(2S)-2,3-Dihydroxy-propyl-1,6,8-trihydroxy-3-methyl-9,10-dioxoanthracene-2-carboxylate |
|
(1S,5′S,6′R,7′S,8′R)-1′,2,5′,6′,7′,8,8′-heptahydroxy-3′,6-dimethoxy-3,6′-dimethyl-5′,6′,7′,8′,8′a,10′a-hexahydro-[1,2′-bianthracene]-9,9′,10,10′-tetraone |
A. variecolor B-17 |
[56] |
|
C20H17ClO6/388 |
7-Chloroaverythrin |
|
(E)-2-chloro-7-(hex-1-en-1-yl)-1,3,6,8-tetrahydroxyanthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C20H20O8/388 |
6,8-Di-O-methylversiconol |
|
2-(1,4-dihydroxybutan-2-yl)-1,3-dihydroxy-6,8-dimethoxyanthraquinone |
A. versicolor EN-7 (Genbank no EU042148) |
[58] |
|
C22H20O7/396 |
6,8-Di-O-methylaverufin |
|
7-hydroxy-9,11-dimetho-xy-2-methyl-3,4,5,6-tetra-hydro-2H-2,6-epoxyanthra[2,3-b]oxocine-8,13-dione |
A. versicolor endolichenic |
[108] |
|
C22H20O7/396 |
6,8-Di-O-methylnidurufin |
|
5,7-dihydroxy-9,11-dime-thoxy-2-methyl-3,4,5,6-tetrahydro-2H-2,6-epoxyanthra[2,3-b]oxocine-8,13-dione |
A. versicolor endolichenic A. versicolor EN-7 (Genbank no EU042148) |
|
|
C22H22O7/398 |
6,8-Di-O-methylaverufanin |
|
1,3-dihydroxy-6,8-dimethoxy-2-(6-methyltetra-hydro-2H-pyran-2-yl)anthracene-9,10-dione |
Fungus ZSUH-36 |
[65] |
|
C22H24O7/400 |
6,1′-O,O-Dimethylaverantin |
|
(S)-1,3,8-trihydroxy-6-methoxy-2-(1-methoxyhexyl)anthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C20H17O9/401 |
Variecolorquinone A |
|
(S)-2,3-dihydroxypropyl 1,6-dihydroxy-8-methoxy-3-methyl-9,10-dioxo-9,10-dihydroanthracene-2-carboxylate |
A.glaucus A. variecolor B-17 |
|
|
C22H24O7/401 |
6,8-Di-O-methylaveran-tin |
|
(S)-1,3-dihydroxy-2-(1-hydroxyhexyl)-6,8-dimetho-xyanthraquinone |
A. versicolor EN-7 (Genbank no EU042148) A. sp. SCSIO F063 |
|
|
C21H19ClO6/402 |
6-O-Methyl-7-chloroaverythrin |
|
(E)-2-chloro-7-(hex-1-en-1-yl)-1,6,8-trihydroxy-3-methoxyanthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C20H19ClO7/407 |
(1′S)-7-Chloroaverantin |
|
(S)-2-chloro-1,3,6,8-tetrahydroxy-7-(1-hydroxyhexyl)anthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C22H23ClO7/407 |
6,1′-O,O-Dimethyl-7-chloroaverantin |
|
(S)-2-chloro-1,6,8-trihydroxy-3-methoxy-7-(1-methoxyhexyl)anthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C23H26O7/414 |
6,8,1′-Tri-O-methyl-averantin |
|
1,3-dihydroxy-6,8-dimethoxy-2-(1-methoxyhexyl)anthraquinone |
A.versicolor endolichenic Fungus ZSUH-36 |
|
|
C21H19O9/415 |
6-3-O-(Ribofuranosyl)questin |
|
1,6-dihydroxy-6-O-(ribofuranosyl)-8-methoxy-3-methylanthraquinone |
Eurotium rubrum |
[63] |
|
C21H21ClO7/420 |
1′-O-methyl-7-chloro averantin |
|
(S)-2-chloro-1,3,6,8-tetrahydroxy-7-(1-methoxy-hexyl)anthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C21H21ClO7/421 |
6-O-methyl-7-chloro-averantin |
|
(S)-2-chloro-1,6,8-trihydroxy-7-(1-hydroxyhexyl)-3-methoxyanthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C24H28O8/444 |
Averantin-1′-butyl ether |
|
(S)-2-(1-butoxyhexyl)-1,3,6,8-tetrahydroxy-anthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C24H27ClO7/463 |
7-Chloroaverantin-1′-butyl ether |
|
(S)-2-(1-butoxyhexyl)-7-chloro-1,3,6,8-tetrahy-droxyanthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C21H21BrO7/465 |
6-O-Methyl-7-bromoaverantin |
|
(S)-2-bromo-1,6,8-trihydroxy-7-(1-hydroxyhexyl)-3-methoxyanthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C22H23BrO7/479 |
6,1′-O,O-Dimethyl-7-bromoaverantin |
|
(S)-2-bromo-1,6,8-trihydroxy-3-methoxy-7-(1-methoxyhexyl)anthraquinone |
A. sp. SCSIO F063 |
[55] |
|
C24H23O11/487 |
Macrosporin2-O-(6′-acetyl)-a-d-glucopyranoside |
|
((2R,3S,4S,5R,6R)-3,4,5-trihydroxy-6-((8-hydroxy-6-methoxy-3-methyl-9,10-dioxo-9,10-dihydroanthracen-2-yl)oxy)tetrahydro-2H-pyran-2-yl)methyl acetate |
Stemphylium sp. 33231 |
[93] |
|
C28H24O10/520 |
Penicillanthranin A |
|
(1S,3R,4S)-1-(6-ethyl-1,3,8-trihydroxy-9,10-dioxo-9,10-dihydroanthracen-2-yl)-6,8-dihy-droxy-3,4,5-trimethyliso-chroman-7-carboxylic acid |
P. citrinum PSU-F51 |
[88] |
|
C28H24O11/536 |
Penicillanthranin B |
|
(1S,3R,4S)-6,8-dihydroxy-3,4,5-trimethyl-1-(1,3,8-trihydroxy-6-(hydroxy-methyl)-9,10-dioxo-9,10-dihydroanthracen-2-yl)isochroman-7-carboxylic acid |
P. citrinum PSU-F51 |
[88] |
|
C32H24O8/536 |
(trans)-R (cis)-Emodin-Physcion bianthrone |
|
2,4,4′,5,5′-pentahydroxy-2′-methoxy-7,7′-dimethyl-[9,9′-bianthracene]-10,10′(9H,9′H)-dione |
A. glaucus |
|
|
C32H21O10/565 |
Alterporriol Q |
|
1′,2,7′,8-tetrahydroxy-3′,6-dimethoxy-3,6′-dimethyl-[1,2′-bianthracene]-9,9′,10,10′-tetraone |
Al. sp. ZJ-2008003 |
[48] |
|
C32H21O10/565 |
Alterporriol R |
|
2,4′,6′,8-tetrahydroxy-2′,6-dimethoxy-3,7′-dimethyl-[1,1′-bianthracene]-9,9′,10,10′-tetraone |
Al. sp. ZJ-2008003 |
[48] |
|
C32H22O10/566 |
Alterporriol V |
|
2,2′,8,8′-tetrahydroxy-6,6′-dimethoxy-3,3′-dimethyl-[1,1′-bianthracene]-9,9′,10,10′-tetraone |
Stemphylium sp. 33231 |
[93] |
|
C30H22O12/574 |
Cytoskyrin A |
|
(6R,14R,17S,18R,19R,20S)-1,7,9,15,17,20-hexahydroxy-3,11-dimethoxy-6,13a,5a,14-(epibutane[1,2,3,4]tetrayl)cycloocta[1,2-b:5,6-b′]dinaphtha-lene-5,8,13,16(6H,14H)-tetraone |
Curvularia lunata |
|
|
C32H26O10/586 |
Alterporriol K |
|
(5S,8R)-4,4′,5,7′,8-penta-hydroxy-1,1′-dimethoxy-6′,8-dimethyl-5,6,7,8-tetrahydro-[2,2′-bianthracene]-9,9′,10,10′-tetraone |
Al. sp. ZJ9-6B Al. sp. ZJ-2008003 |
|
|
C32H30O13/590 |
Alterporriol T |
|
(6R,6′S,7R,7′R,8R)-1′,4,6,6′,7,7′,8-heptahydroxy-2,3′-dimethoxy-6′,7-dime-thyl-5,5′,6,6′,7,7′,8,8′-octahydro-[1,2′-bianthracene]-9,9′,10,10′-tetraone |
Stemphylium sp. 33231 |
[93] |
|
C32H25O12/601 |
Alterporriol L |
|
(6S,7R,8R)-4,4′,6,7,7′,8-hexahydroxy-1,1′-dimethoxy-6′,7-dimethyl-5,6,7,8-tetrahydro-[2,2′-bianthracene]-9,9′,10,10′-tetraone |
Al. sp. ZJ9-6B Al. sp. ZJ-2008003 |
|
|
C32H25O12/601 |
Alterporriol M |
|
(6S,7S,8R)-4,4′,6,7,7′,8-hexahydroxy-1,1′-dimethoxy-6′,7-dimethyl-5,6,7,8-tetrahydro-[2,2′-bianthracene]-9,9′,10,10′-tetraone |
Al. sp. ZJ9-6B Al. sp. ZJ-2008003 |
|
|
C32H25O12/601 |
Alterporriol P |
|
(5′R,6′R,7′R)-1′,2,5′,6′,7′,8-hexahydroxy-3′,6-dimethoxy-3,6′-dimethyl-5′,6′,7′,8′,8′a,10′a-hexahydro-[1,2′-bianthracene]-9,9′,10,10′-tetraone |
Al. sp. ZJ-2008003 |
[48] |
|
C32H26O12/602 |
Alterporriol W |
|
(1′R,6R,7R,8R)-2′,4,6,7,8,8′-hexahydroxy-2,6′-dimethoxy-3′,7-dimethyl-5,6,7,8-tetrahydro-[1,1′-bianthracene]-9,9′,10,10′-tetraone |
Stemphylium sp. 33231 |
[93] |
|
C32H30O12/606 |
Alterporriol U |
|
(6R,6′S,7S,7′R)-1′,4,6,6′,7,7′-hexahydroxy-2,3′-di-methoxy-6′,7-dimethyl-5,5′,6,6′,7,7′,8,8′-octahy-dro-[1,2′-bianthracene]-9,9′,10,10′-tetraone |
Stemphylium sp. 33231 |
[93] |
|
C31H32O13/612 |
Alterporriol S |
|
(2′S,3′R,4′S,6R,7S,9′R)-1,2′,3′,4,5′,6,7,8′-octahy-droxy-7′-methoxy-2′,4′,7-trimethyl-2′,3′,4′,4′a,5,6,7,8,9′,9′a-decahydro-[2,9′-bianthracene]-9,10,10′(1′H)-trione |
Al. sp. (SK11) |
[47] |
|
C32H25O13/617 |
(+)-aS-alterporriol C |
|
(1S,5′S,6′R,7′S,8′R)-1′,2,5′,6′,7′,8,8′-heptahydroxy-3′,6-dimethoxy-3,6′-dime-thyl-5′,6′,7′,8′,8′a,10′a-hexahydro-[1,2′-bianthracene]-9,9′,10,10′-tetraone |
Al. sp. (SK11) |
[47] |
|
C32H25O13/617 |
Alterporriol C |
|
(1S,5′R,6′S,7′R,8′S)-1′,2,5′,6′,7′,8,8′-heptahydroxy-3′,6-dimethoxy-3,6′-dime-thyl-5′,6′,7′,8′,8′a,10′a-hexahydro-[1,2′-bianthracene]-9,9′,10,10′-tetraone |
Al. sp. (SK11) Al. sp. ZJ-2008003 |
|
|
C32H29O14/637 |
Alterporriol N |
|
(6R,6′R,7R,7′R,8R,8′R)-4,4′,6,6′,7,7′,8,8′-octahy-droxy-2,2′-dimethoxy-7,7′-dimethyl-5,5′,6,6′,7,7′,8,8′-octahydro-[1,1′-bianthracene]-9,9′,10,10′-tetraone |
Al. sp. ZJ-2008003 |
[48] |
|
C32H29O14/637 |
Alterporriol O |
|
(2R,2′R,3S,3′S)-2,2′,3,3′,8,8′-hexahydroxy-6,6′-dimethoxy-3,3′-dimethyl-1,1′,2,2′,3,3′,4,4′-octahy-dro-[1,1′-bianthracene]-9,9′,10,10′-tetraone |
Al. sp. ZJ-2008003 |
[48] |
|
C34H33O17/713 |
Acetylalterporriol D |
|
(1′S,5S,5′S,6R,6′R,7S,7′S,8R,8′R)-4,4′,5,5′,6′,7,7′,8,8′-nonahydroxy-2,2′-dimethoxy-7,7′-dimethyl-9,9′,10,10′-tetraoxo-5,5′,6,6′,7,7′,8,8′,9,9′,10,10′-dodecahydro-[1,1′-bianthracen]-6-yl acetate |
Stemphylium globuliferum |
[94] |
|
C34H33O17/713 |
Acetylalterporriol E |
|
(1′R,5S,5′S,6R,6′R,7S,7′S,8R,8′R)-4,4′,5,5′,6′,7,7′,8,8′-nonahydroxy-2,2′-dimethoxy-7,7′-dimethyl-9,9′,10,10′-tetraoxo-5,5′,6,6′,7,7′,8,8′,9,9′,10,10′-dodecahydro-[1,1′-bianthracen]-6-yl acetate |
Stemphylium globuliferum |
[94] |
|
C34H33O17/713 |
Alterporriol D |
|
(1S,5S,5′S,6R,6′R,7S,7′S,8R,8′R)-4,4′,5,5′,6,6′,7,7′,8,8′-decahydroxy-2,2′-dimethoxy-7,7′-dimethyl-5,5′,6,6′,7,7′,8,8′-octahy-dro-[1,1′-bianthracene]-9,9′,10,10′-tetraone |
Stemphylium globuliferum |
[94] |
|
C34H33O17/713 |
Alterporriol E |
|
(1R,5S,5′S,6R,6′R,7S,7′S,8R,8′R)-4,4′,5,5′,6,6′,7,7′,8,8′-decahydroxy-2,2′-dimethoxy-7,7′-dimethyl-5,5′,6,6′,7,7′,8,8′-octahy-dro-[1,1′-bianthracene]-9,9′,10,10′-tetraone |
Stemphylium globuliferum |
[94] |
|
C48H40O21/952 |
Stemphylanthranol A |
|
(5S,5′′S,6R,6′′R,7S,7′′S,8R,8′′R)-2′,4,4′′,5,5′′,6,6′′,7,7′′,8,8′,8′′-dodecahydroxy-2,2′′,6′-trimethoxy-3′,7,7′′-trimethyl-5,5′′,6,6′′,7,7′′,8,8′′-octahydro-[1,1′:5′,1′′-teranthracene]-9,9′,9′′,10,10′,10′′-hexaone |
Stemphylium globuliferum |
[94] |
|
C48H40O21/952 |
Stemphylanthranol B |
|
(5S,5′′R,6R,6′′R,7S,7′′R,8R,8′′R)-2′,4,4′′,5,5′′,6,7,7′′,8,8′,8′′-undecahydroxy-2,2′′,6′-trimethoxy-3′,6′′,7,7′′-tetramethyl-5,5′′,6,6′′,7,7′′,8,8′′-octahydro-[1,1′:7′,1′′-tetranthracene]-9,9′,9′′,10,10′,10′′-hexaone |
Stemphylium globuliferum |
[94] |
|
---- |
7-Chloroemodic acid |
---- |
---- |
Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) |
|
|
---- |
7-Chloroemodinal |
---- |
---- |
Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.)L. hafellneri L. leprolytoides |
|
|
---- |
7-Chloro-1,6,8-trihydroxy-3-methyl-10-anthrone |
---- |
---- |
Caloplaca spp. (e.g., C. ehrenbergii, C. schaereri, C. spitsbergensis, etc.) |


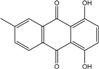
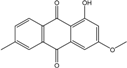
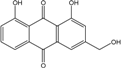
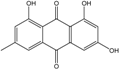

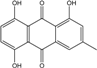
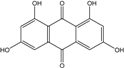
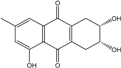
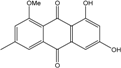

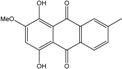
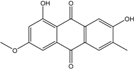
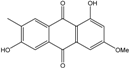
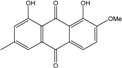
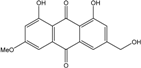
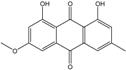
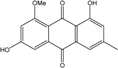
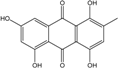

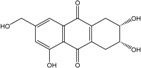
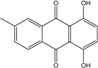
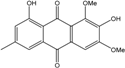

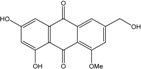












































































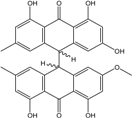






















Abbreviations: A.: Aspergillus; Al.: Alternaria; P.: Penicillium. Orange brown:  ; Orange:
; Orange:  ; Yellow:
; Yellow:  ; Red:
; Red:  ; Bronze:
; Bronze:  .
.
Ubiquitous Fungi
Endophytes and/or Pathogens
Lichens
3. Biosynthesis and Known Roles for Anthraquinones in Fungi
3.1. Biosynthetic Route and Genes Involved
3.2. Roles in the Biology of Fungi
This entry is adapted from the peer-reviewed paper 10.3390/md14040064