 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | arrigo de benedetti | -- | 1825 | 2023-09-08 11:01:26 | | | |
| 2 | Lindsay Dong | + 2 word(s) | 1827 | 2023-09-08 11:31:05 | | | | |
| 3 | Lindsay Dong | Meta information modification | 1827 | 2023-09-11 10:40:29 | | |
Video Upload Options
DNA damage repair lies at the core of all cells’ survival strategy, including the survival strategy of cancerous cells. Therefore, targeting such repair mechanisms forms the major goal of cancer therapeutics. The mechanism of DNA repair has been tousled with the discovery of multiple kinases. Studies on tousled-like kinases have brought significant clarity on the effectors of these kinases which stand to regulate double-strand break (DSB) repair.
1. Introduction
2. Role of TLK1 in DSB Repair
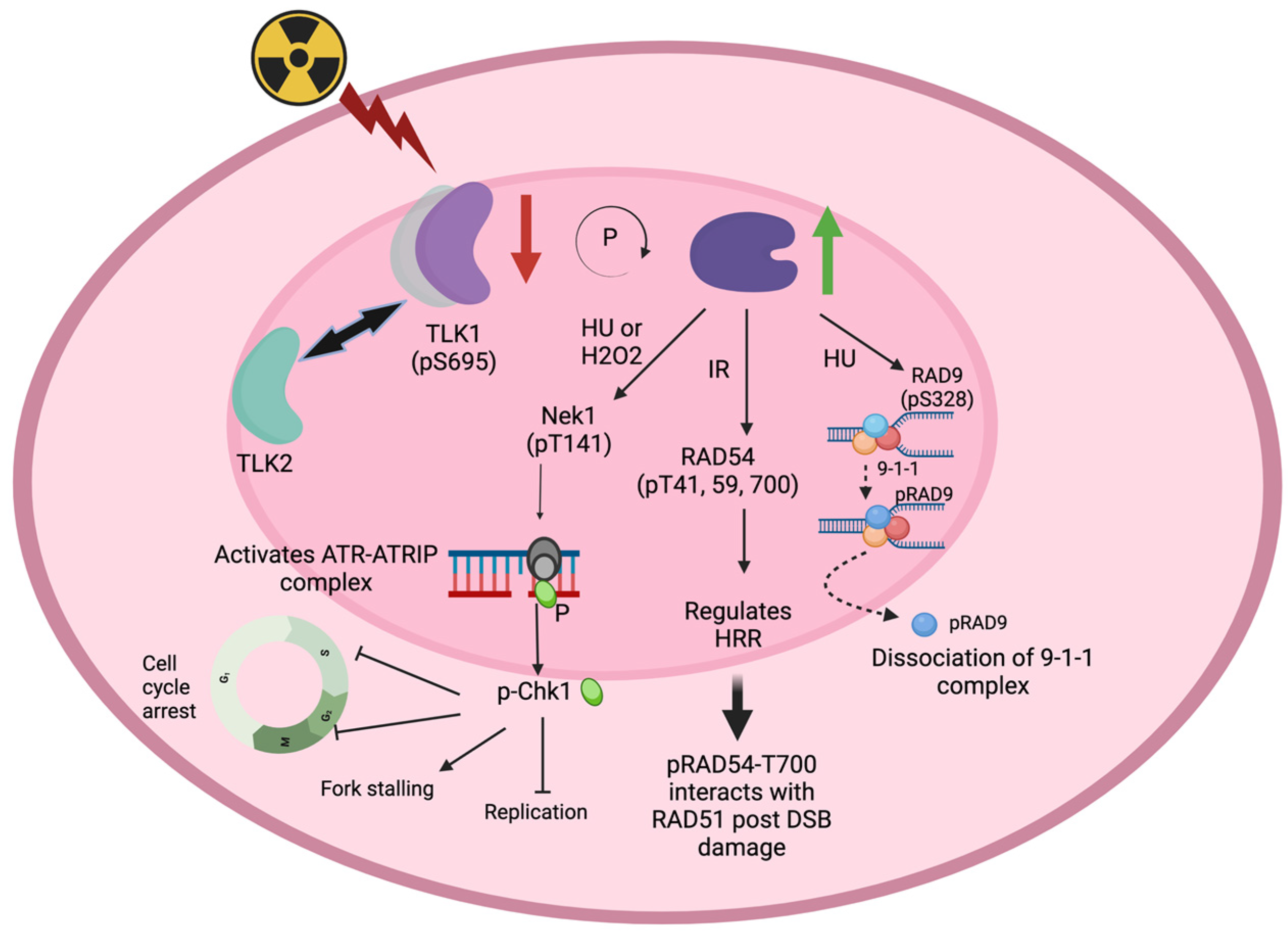
3. Role of TLK1 in Eukaryotic Recombination Repair
4. TLKs and DNA Damage and Checkpoint Functions
5. Role of TLK1 in Cancer
6. Targeting TLK1 for Cancer Treatment
After the seminal discovery that TLK1 has an important modulatory role in DDR and DNA repair, the quest turned rapidly into trying to identify specific TLK inhibitors to enhance the effectiveness of XRT or radiomimetic drugs. This led to the initial identification of certain phenothiazine (PTH) antipsychotics as surprisingly specific TLK inhibitors that, in fact, enhanced the killing of cancer cells, in vitro and in xenografts, when combined with IR or doxorubicin [37]. A newer PTH scaffold, called J54, that substantially lacks any anti-dopaminergic undesirable effect, showed very promising results in the regression of androgen-sensitive prostate cancer cells largely by passing the DDR and thereby enforcing entry into catastrophic mitotic progression, even resulting in substantial tumor regression in SCID xenografts [36]. Other studies on the structure/function of TLKs have revealed additional potential inhibitors [38]. Considering the availability of such drugs, of even greater importance now is the rebound effort to target the activity of TLK1 in RAD9 and RAD54, with the obvious suggestion that translesion (TSL) repair, single-strand gaps, and HRR can be simultaneously targeted. Various chemotherapeutic agents, therefore, fall under the umbrella of TLK inhibitors for their therapeutic potentiation. These include, for example: topoisomerase poisons, bleomycin, MMC, PARPis, and cisplatin, all of which ultimately lead to the formation of SSBs and DSBs. For some of these (e.g., doxorubicin and cisplatin), direct evidence of synthetic lethality in combination with TLK inhibitors has already been verified [37][39].
References
- Silljé, H.H.W.; Takahashi, K.; Tanaka, K.; Van Houwe, G.; Nigg, E.A. Mammalian homologues of the plant Tousled gene code for cell-cycle-regulated kinases with maximal activities linked to ongoing DNA replication. EMBO J. 1999, 18, 5691–5702.
- Li, Y.; DeFatta, R.; Anthony, C.; Sunavala, G.; De Benedetti, A. A translationally regulated Tousled kinase phosphorylates histone H3 and confers radioresistance when overexpressed. Oncogene 2001, 20, 726–738.
- Groth, A.; Lukas, J.; Nigg, E.A.; Silljé, H.H.W.; Wernstedt, C.; Bartek, J.; Hansen, K. Human Tousled like kinases are targeted by an ATM- and Chk1-dependent DNA damage checkpoint. EMBO J. 2003, 22, 1676–1687.
- Ghosh, I.; Kwon, Y.; Shabestari, A.B.; Chikhale, R.; Chen, J.; Wiese, C.; Sung, P.; De Benedetti, A. TLK1-Mediated RAD54 Phosphorylation Spatio-Temporally Regulates Homologous Recombination Repair. Nucleic Acids Res. 2023. ahead of print.
- Segura-Bayona, S.; Stracker, T.H. The Tousled-like kinases regulate genome and epigenome stability: Implications in development and disease. Cell. Mol. Life Sci. 2019, 76, 3827–3841.
- Li, Z.; Gourguechon, S.; Wang, C.C. Tousled-like kinase in a microbial eukaryote regulates spindle assembly and S-phase progression by interacting with Aurora kinase and chromatin assembly factors. J. Cell Sci. 2007, 120, 3883–3894.
- Awate, S.; De Benedetti, A. TLK1B mediated phosphorylation of Rad9 regulates its nuclear/cytoplasmic localization and cell cycle checkpoint. BMC Mol. Biol. 2016, 17, 3.
- Sunavala-Dossabhoy, G.; De Benedetti, A. Tousled homolog, TLK1, binds and phosphorylates Rad9; TLK1 acts as a molecular chaperone in DNA repair. DNA Repair 2009, 8, 87–102.
- Klimovskaia, I.M.; Young, C.; Strømme, C.B.; Menard, P.; Jasencakova, Z.; Mejlvang, J.; Ask, K.; Ploug, M.; Nielsen, M.L.; Jensen, O.N.; et al. Tousled-like kinases phosphorylate Asf1 to promote histone supply during DNA replication. Nat. Commun. 2014, 5, 3394.
- Sen, S.P.; De Benedetti, A. TLK1B promotes repair of UV-damaged DNA through chromatin remodeling by Asf1. BMC Mol. Biol. 2006, 7, 37.
- Singh, V.; Connelly, Z.M.; Shen, X.; De Benedetti, A. Identification of the proteome complement of humanTLK1 reveals it binds and phosphorylates NEK1 regulating its activity. Cell Cycle 2017, 16, 915–926.
- Mengwasser, K.E.; Adeyemi, R.O.; Leng, Y.; Choi, M.Y.; Clairmont, C.; D’Andrea, A.D.; Elledge, S.J. Genetic Screens Reveal FEN1 and APEX2 as BRCA2 Synthetic Lethal Targets. Mol. Cell 2019, 73, 885–899.e886.
- Kiianitsa, K.; Solinger Jachen, A.; Heyer, W.-D. Terminal association of Rad54 protein with the Rad51–dsDNA filament. Proc. Natl. Acad. Sci. USA 2006, 103, 9767–9772.
- Timiri Shanmugam, P.S.; Nair, R.P.; De Benedetti, A.; Caldito, G.; Abreo, F.; Sunavala-Dossabhoy, G. Tousled kinase activator, gallic acid, promotes homologous recombinational repair and suppresses radiation cytotoxicity in salivary gland cells. Free. Radic. Biol. Med. 2016, 93, 217–226.
- Sunavala-Dossabhoy, G.; Li, Y.; Williams, B.; De Benedetti, A. A dominant negative mutant of TLK1 causes chromosome missegregation and aneuploidy in normal breast epithelial cells. BMC Cell Biol. 2003, 4, 16.
- Yang, S.; Liu, L.; Cao, C.; Song, N.; Wang, Y.; Ma, S.; Zhang, Q.; Yu, N.; Ding, X.; Yang, F.; et al. USP52 acts as a deubiquitinase and promotes histone chaperone ASF1A stabilization. Nat. Commun. 2018, 9, 1285.
- Sukackaite, R.; Cornacchia, D.; Jensen Malene, R.; Mas, P.J.; Blackledge, M.; Enervald, E.; Duan, G.; Auchynnikava, T.; Köhn, M.; Hart, D.J.; et al. Mouse Rif1 is a regulatory subunit of protein phosphatase 1 (PP1). Sci. Rep. 2017, 7, 2119.
- Escribano-Díaz, C.; Orthwein, A.; Fradet-Turcotte, A.; Xing, M.; Young Jordan, T.F.; Tkáč, J.; Cook Michael, A.; Rosebrock Adam, P.; Munro, M.; Canny Marella, D.; et al. A Cell Cycle-Dependent Regulatory Circuit Composed of 53BP1-RIF1 and BRCA1-CtIP Controls DNA Repair Pathway Choice. Mol. Cell 2013, 49, 872–883.
- Batenburg, N.L.; Walker, J.R.; Noordermeer, S.M.; Moatti, N.; Durocher, D.; Zhu, X.-D. ATM and CDK2 control chromatin remodeler CSB to inhibit RIF1 in DSB repair pathway choice. Nat. Commun. 2017, 8, 1921.
- Tang, M.; Chen, Z.; Wang, C.; Feng, X.; Lee, N.; Huang, M.; Zhang, H.; Li, S.; Xiong, Y.; Chen, J. Histone chaperone ASF1 acts with RIF1 to promote DNA end-joining in BRCA1-deficient cells. J. Biol. Chem. 2022, 298, 101979.
- Xu, L.; Blackburn, E.H. Human Rif1 protein binds aberrant telomeres and aligns along anaphase midzone microtubules. J. Cell Biol. 2004, 167, 819–830.
- Segura-Bayona, S.; Villamor-Payà, M.; Attolini, C.S.-O.; Koenig, L.M.; Sanchiz-Calvo, M.; Boulton, S.J.; Stracker, T.H. Tousled-Like Kinases Suppress Innate Immune Signaling Triggered by Alternative Lengthening of Telomeres. Cell Rep. 2020, 32, 107983.
- Lisby, M.; Barlow, J.H.; Burgess, R.C.; Rothstein, R. Choreography of the DNA Damage Response: Spatiotemporal Relationships among Checkpoint and Repair Proteins. Cell 2004, 118, 699–713.
- Carrera, P.; Moshkin, Y.M.; Grönke, S.; Silljé, H.H.W.; Nigg, E.A.; Jäckle, H.; Karch, F. Tousled-like kinase functions with the chromatin assembly pathway regulating nuclear divisions. Genes Dev. 2003, 17, 2578–2590.
- Han, Z.; Riefler, G.M.; Saam, J.R.; Mango, S.E.; Schumacher, J.M. The C. elegans Tousled-like Kinase Contributes to Chromosome Segregation as a Substrate and Regulator of the Aurora B Kinase. Curr. Biol. 2005, 15, 894–904.
- Takayama, Y.; Kokuryo, T.; Yokoyama, Y.; Ito, S.; Nagino, M.; Hamaguchi, M.; Senga, T. Silencing of Tousled-like kinase 1 sensitizes cholangiocarcinoma cells to cisplatin-induced apoptosis. Cancer Lett. 2010, 296, 27–34.
- Singh, V.; Jaiswal, P.K.; Ghosh, I.; Koul, H.K.; Yu, X.; De Benedetti, A. The TLK1-Nek1 axis promotes prostate cancer progression. Cancer Lett. 2019, 453, 131–141.
- Singh, V.; Jaiswal, P.K.; Ghosh, I.; Koul, H.K.; Yu, X.; De Benedetti, A. Targeting the TLK1/NEK1 DDR axis with Thioridazine suppresses outgrowth of androgen independent prostate tumors. Int. J. Cancer 2019, 145, 1055–1067.
- Ibrahim, K.; Abdul Murad, N.A.; Harun, R.; Jamal, R.; Ibrahim, K.; Abdul Murad, N.A.; Harun, R.; Jamal, R.; Ibrahim, K.; Abdul Murad, N.A.; et al. Knockdown of Tousled-like kinase 1 inhibits survival of glioblastoma multiforme cells. Int. J. Mol. Med. 2020, 46, 685–699.
- Lee, S.-B.; Segura-Bayona, S.; Villamor-Payà, M.; Saredi, G.; Todd, M.A.M.; Attolini, C.S.-O.; Chang, T.-Y.; Stracker, T.H.; Groth, A. Tousled-like kinases stabilize replication forks and show synthetic lethality with checkpoint and PARP inhibitors. Sci. Adv. 2018, 4, eaat4985.
- Jiang, J.; Jia, P.; Zhao, Z.; Shen, B. Key regulators in prostate cancer identified by co-expression module analysis. BMC Genom. 2014, 15, 1015.
- Kim, J.-A.; Anurag, M.; Veeraraghavan, J.; Schiff, R.; Li, K.; Wang, X.-S. Amplification of TLK2 Induces Genomic Instability via Impairing the G2–M Checkpoint. Mol. Cancer Res. 2016, 14, 920–927.
- Lin, M.; Yao, Z.; Zhao, N.; Zhang, C. TLK2 enhances aggressive phenotypes of glioblastoma cells through the activation of SRC signaling pathway. Cancer Biol. Ther. 2019, 20, 101–108.
- Van Roy, N.; Vandesompele, J.; Berx, G.; Staes, K.; Van Gele, M.; De Smet, E.; De Paepe, A.; Laureys, G.; van der Drift, P.; Versteeg, R.; et al. Localization of the 17q breakpoint of a constitutional 1;17 translocation in a patient with neuroblastoma within a 25-kb segment located between the ACCN1 and TLK2 genes and near the distal breakpoints of two microdeletions in neurofibromatosis type 1 patients. Genes Chromosom. Cancer 2002, 35, 113–120.
- Ghosh, I.; Khalil, M.I.; Mirza, R.; King, J.; Olatunde, D.; De Benedetti, A. NEK1-Mediated Phosphorylation of YAP1 Is Key to Prostate Cancer Progression. Biomedicines 2023, 11, 734.
- Singh, V.; Bhoir, S.; Chikhale, R.V.; Hussain, J.; Dwyer, D.; Bryce, R.A.; Kirubakaran, S.; De Benedetti, A. Generation of Phenothiazine with Potent Anti-TLK1 Activity for Prostate Cancer Therapy. iScience 2020, 23, 101474.
- Ronald, S.; Awate, S.; Rath, A.; Carroll, J.; Galiano, F.; Dwyer, D.; Kleiner-Hancock, H.; Mathis, J.M.; Vigod, S.; De Benedetti, A. Phenothiazine Inhibitors of TLKs Affect Double-Strand Break Repair and DNA Damage Response Recovery and Potentiate Tumor Killing with Radiomimetic Therapy. Genes Cancer 2013, 4, 39–53.
- Mortuza, G.B.; Hermida, D.; Pedersen, A.-K.; Segura-Bayona, S.; López-Méndez, B.; Redondo, P.; Rüther, P.; Pozdnyakova, I.; Garrote, A.M.; Muñoz, I.G.; et al. Molecular basis of Tousled-Like Kinase 2 activation. Nat. Commun. 2018, 9, 2535.
- Bhoir, S.; De Benedetti, A. Targeting Prostate Cancer, the ‘Tousled Way’. Int. J. Mol. Sci. 2023, 24, 11100.

